Dynamic Changes in the Bacterial Community During the Fermentation of Traditional Chinese Fish Sauce (TCFS) and Their Correlation with TCFS Quality

Abstract
:1. Introduction
2. Materials and Methods
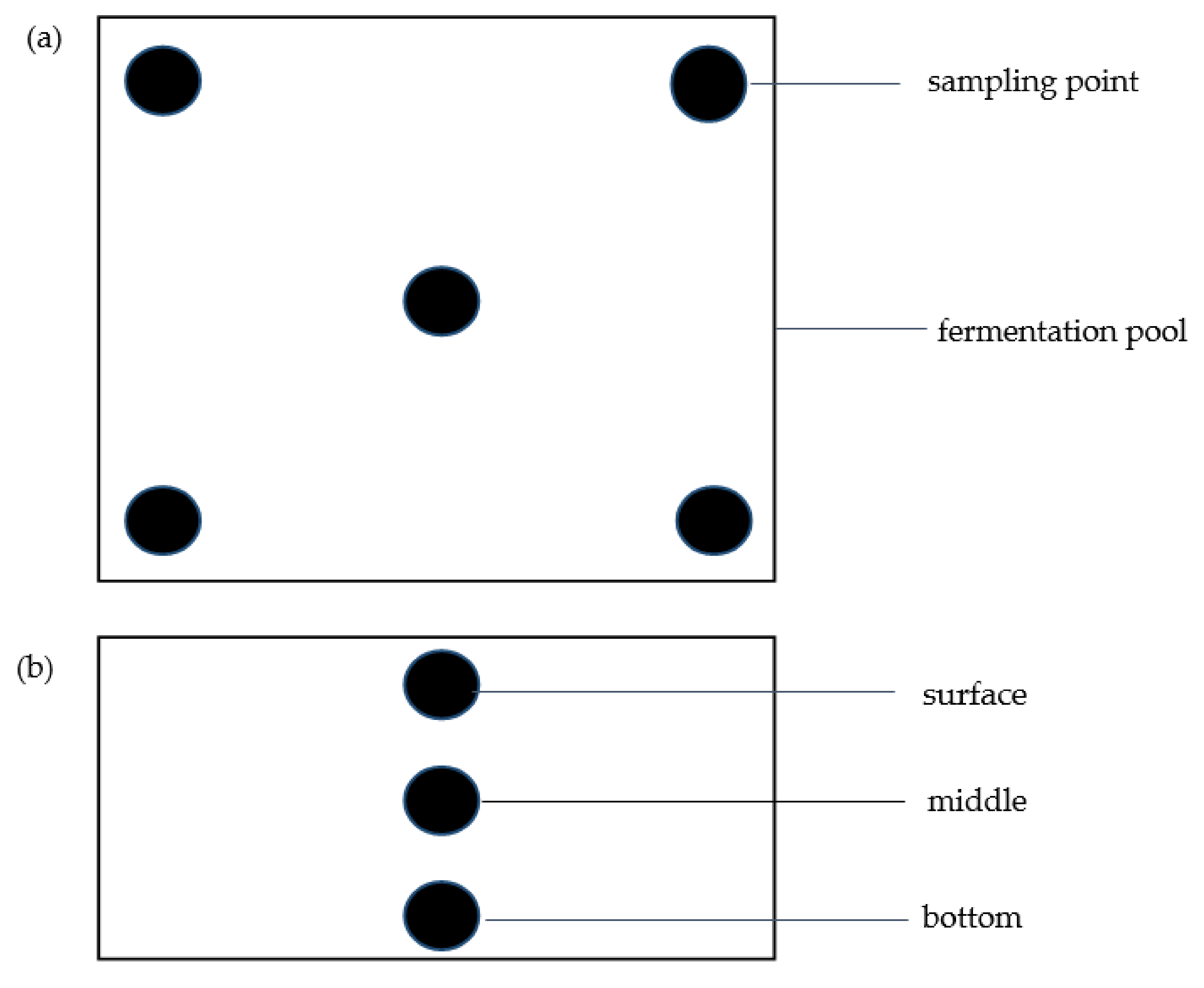
2.1. Sample Collection and Analysis
2.2. DNA Extraction and Illumina MiSeq Sequencing
2.3. Sequencing Processing and Data Analysis
2.4. Statistical Analysis
2.5. Nucleotide Sequence Accession Numbers
3. Results
3.1. Physical and Chemical Indicator Changes in Fish Sauce Samples During Fermentation
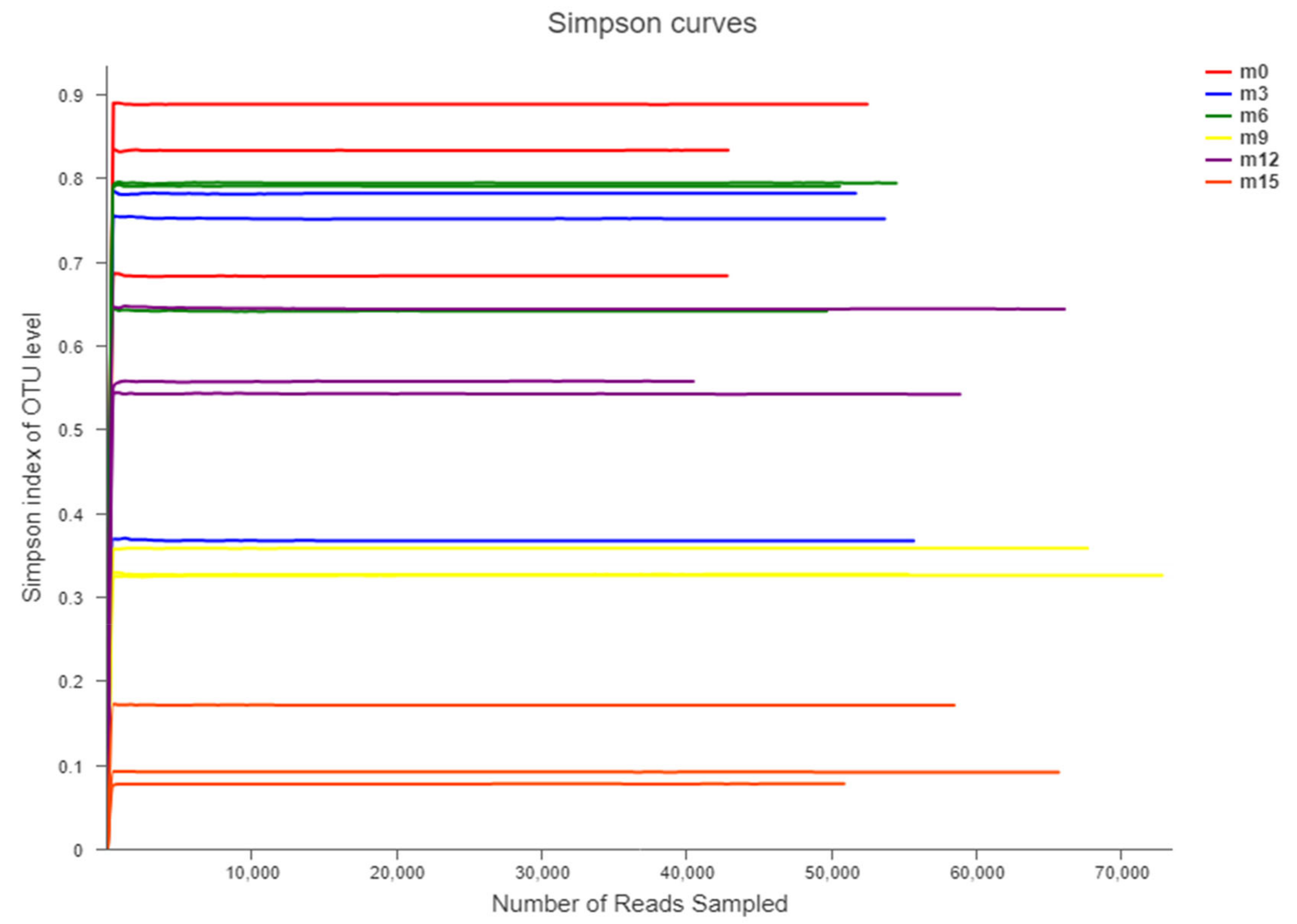
3.2. Illumina MiSeq Sequencing Data Statistics
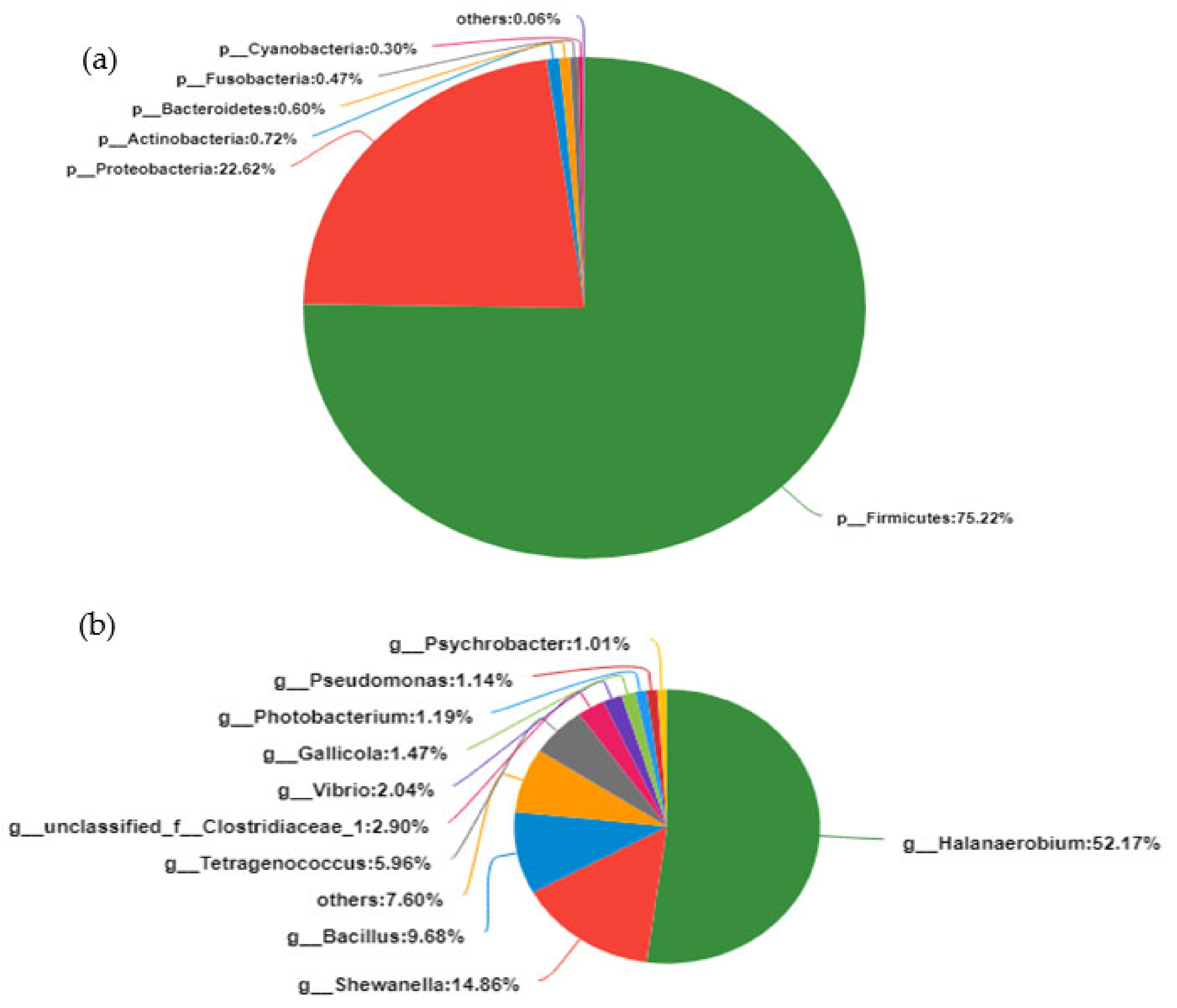
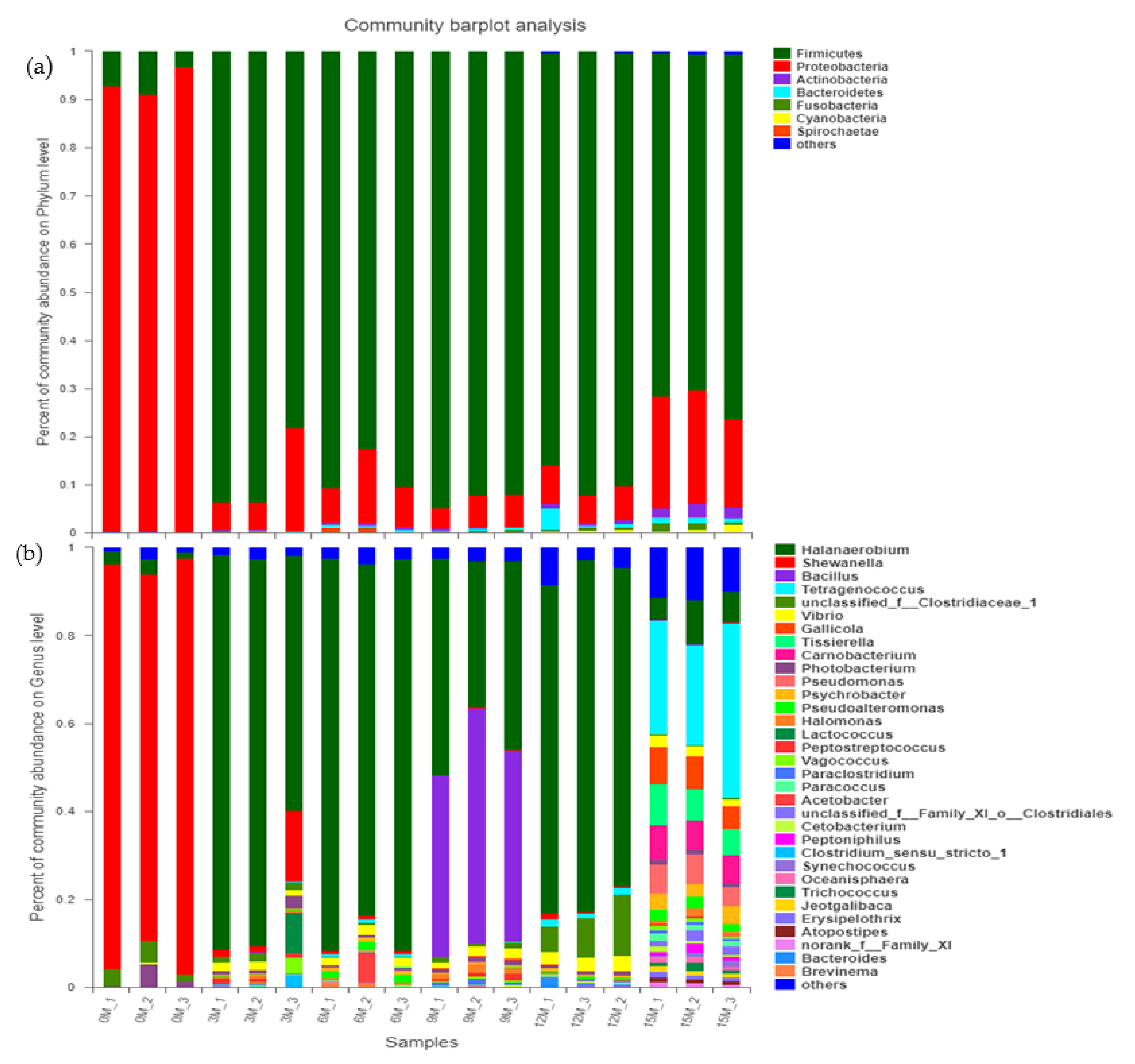
3.3. Analysis of Bacterial Community Composition in the Fish Sauce Fermentation Process
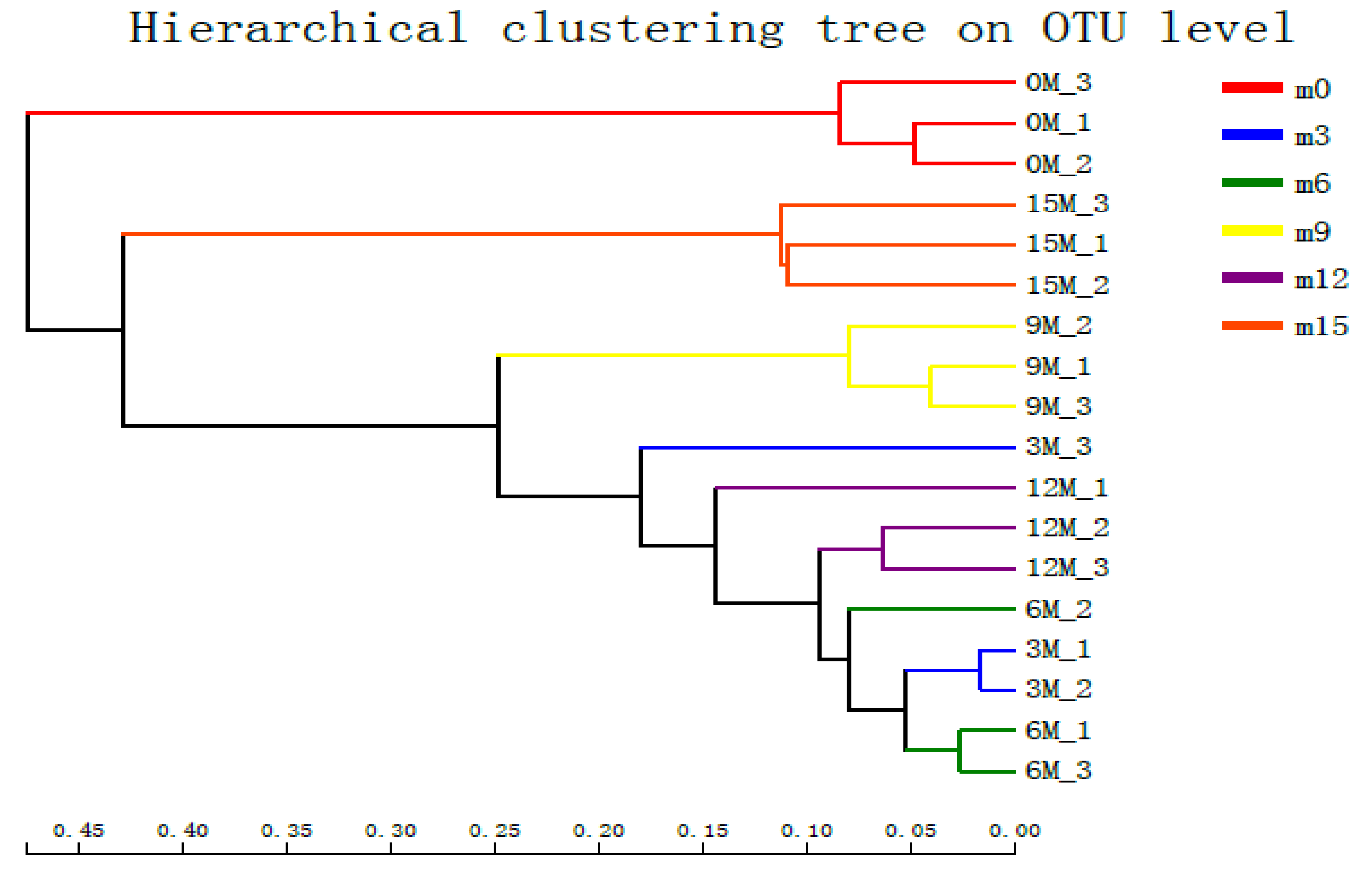
3.4. Beta Diversity Analysis
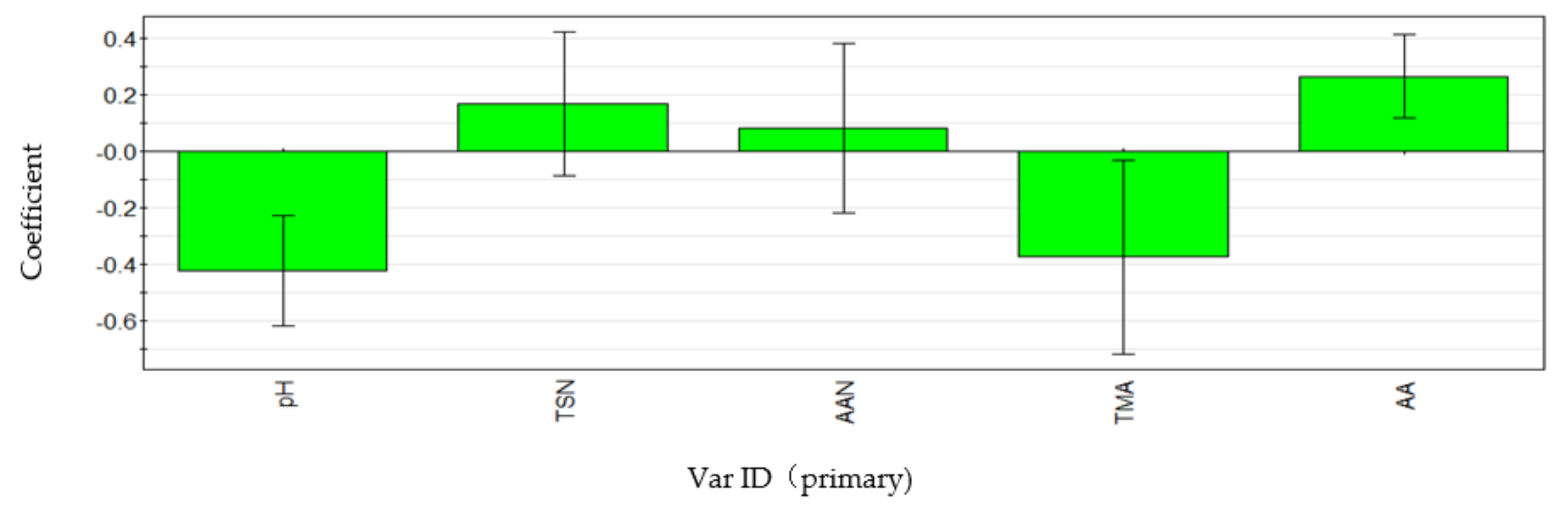
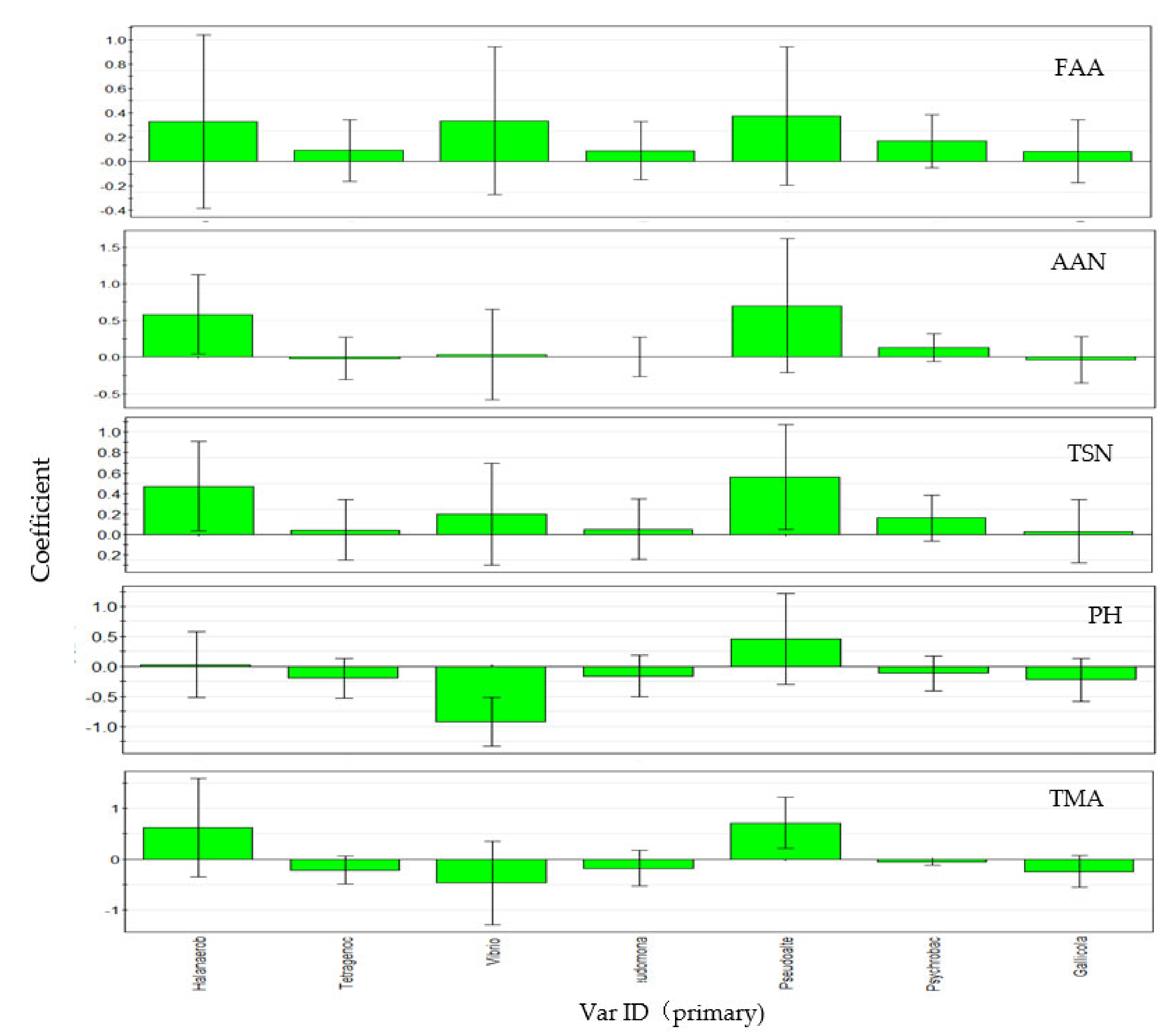
3.5. Correlation Analysis of Environmental Factors
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Carvalho, N.M.D.; Costa, E.M.; Silva, S.; Pimentel, L.; Fernandes, T.H.; Pintado, M.E. Fermented foods and beverages in human diet and their influence on gut microbiota and health. Fermentation 2018, 4, 90. [Google Scholar] [CrossRef]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Choi, Y.; Hwang, I.; Hong, S.; Lee, M. Relationship between chemical characteristics and bacterial community of a Korean salted-fermented anchovy sauce, Myeolchi-Aekjeot. LWT 2016, 73, 251–258. [Google Scholar] [CrossRef]
- Lopecharat, K.P.J. Characteristics of fish sauce made from pacific whiting and surimi by-products during fermentation stage. Food Chem. Toxicol. 2002, 67, 511–516. [Google Scholar] [CrossRef]
- Jiang, J.; Zeng, Q.; Zhu, Z.; Zhang, L. Chemical and sensory changes associated Yu-lu fermentation process-A traditional Chinese fish sauce. Food Chem. 2007, 104, 1629–1634. [Google Scholar] [CrossRef]
- Udomsil, N.; Chen, S.; Rodtong, S.; Yongsawatdigul, J. Quantification of viable bacterial starter cultures of Virgibacillus sp. and Tetragenococcus halophilus in fish sauce fermentation by real-time quantitative PCR. Food Microbiol. 2016, 57, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K. Improvement of fish-sauce odor by treatment with bacteria isolated from the fish-sauce mush (moromi) made from frigate mackerel. J. Food Sci. 2004, 69, 45–49. [Google Scholar] [CrossRef]
- Akolkar, A.V.; Durai, D.; Desai, A.J. Halobacterium sp. SP1(1) as a starter culture for accelerating fish sauce fermentation. J. Appl. Microbiol. 2010, 109, 44–53. [Google Scholar]
- Lopetcharat, K.; Choi, Y.J.; Park, J.W.; Daeschel, M.A. Fish sauce products and manufacturing: A review. Food Rev. Int. 2001, 17, 65–88. [Google Scholar] [CrossRef]
- Namwong, S.; Hiraga, K.; Takada, K.; Tsunemi, M.; Tanasupawat, S.; Oda, K. A Halophilic Serine Proteinase from Halobacillus sp. SR5-3 Isolated from fish sauce: Purification and characterization. Biosci. Biotechnol. Biochem. 2006, 70, 1395–1401. [Google Scholar] [CrossRef]
- Hiraga, K.; Nishikata, Y.; Namwong, S.; Tanasupawat, S.; Takada, K.; Oda, K. Purification and characterization of serine proteinase from a halophilic bacterium, Filobacillus sp. RF2-5. Biosci. Biotechnol. Biochem. 2005, 69, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sinsuwan, S.; Rodtong, S.; Yongsawatdigul, J. Production and characterization of NaCl-activated proteinases from Virgibacillus sp. SK33 isolated from fish sauce fermentation. Process. Biochem. 2008, 43, 185–192. [Google Scholar] [CrossRef]
- Uchida, H.; Kondo, D.; Yamashita, S.; Tanaka, L.H.; Nagano, H.; Uwajima, T. Purification and properties of a protease produced by Bacillus subtilis CN2 isolated from a Vietnamese fish sauce. World J. Microb. Biotech. 2004, 20, 579–582. [Google Scholar] [CrossRef]
- Montriwong, A.; Rodtong, S.; Yongsawatdigul, J. Detergent-stable salt-activated proteinases from Virgibacillus halodenitrificans SK1-3-7 isolated from fish sauce fermentation. Appl. Biochem. Biotech. 2015, 176, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.S.; Zhang, B.C.; Qi, X.Y.; Sun, Z.H.; He, X.L.; Liu, Y.Z.; Li, J.; Chen, K.K.; Lin, Z.X. Root-Associated Endophytic Bacterial Community Composition of Pennisetum sinese from Four Representative Provinces in China. Microorganisms 2019, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Landsman, A.; St-Pierre, B.; Rosales-Leija, M.; Brown, M.; Gibbons, W. Investigation of the Potential Effects of Host Genetics and Probiotic Treatment on the Gut Bacterial Community Composition of Aquaculture-raised Pacific Whiteleg Shrimp, Litopenaeus vannamei. Microorganisms 2019, 7, 217. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhuang, X.; Yu, Z.; Wang, Z.; Guo, X.; Xiang, W.; Huang, S. Community Structures and Antifungal Activity of Root-Associated Endophytic Actinobacteria of Healthy and Diseased Soybean. Microorganisms 2019, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Danilo, E.; Francesca, D.F.; Antonietta, L.S.; Michele, I. “Remake” by high-throughput sequencing of the microbiota involved in the production of water buffalo mozzarella cheese. Appl. Environ. Microb. 2012, 78, 8142–8145. [Google Scholar]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef]
- Mi-Ja, J.; Young-Do, N.; Seong Woon, R.; Jin-Woo, B. Unexpected convergence of fungal and bacterial communities during fermentation of traditional Korean alcoholic beverages inoculated with various natural starters. Food Microbiol. 2012, 30, 112–123. [Google Scholar]
- Wang, P.; Mao, J.; Meng, X.; Li, X.; Liu, Y.; Feng, H. Changes in flavour characteristics and bacterial diversity during the traditional fermentation of Chinese rice wines from Shaoxing region. Food Control 2014, 44, 58–63. [Google Scholar] [CrossRef]
- Berbegal, C.; Borruso, L.; Fragasso, M.; Tufariello, M.; Russo, P.; Brusetti, L.; Spano, G.; Capozzi, V. A metagenomic-based approach for the characterization of bacterial diversity associated with spontaneous malolactic fermentations in wine. Int. J. Mol. Sci. 2019, 20, 3980. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lu, Z.; Shi, J.; Xu, Z. Exploring flavour-producing core microbiota in multispecies solid-state fermentation of traditional Chinese vinegar. Sci. Rep. UK 2016, 6, 26818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.; Lee, S.; Lim, S. Microbial community analysis of Korean soybean pastes by next-generation sequencing. Int. J. Food Microbiol. 2012, 155, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.; Yi, S.; Lim, S. Bacterial diversity of cheonggukjang, a traditional Korean fermented food, analyzed by barcoded pyrosequencing. Food Control 2012, 28, 135–142. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Microbial community dynamics during fermentation of doenjang-meju, traditional Korean fermented soybean. Int. J. Food Microbiol. 2014, 185, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef]
- Hiroshi, O.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Monitoring of the microbiota profile in nukadoko, a naturally fermented rice bran bed for pickling vegetables. J. Biosci. Bioeng. 2014, 118, 520–525. [Google Scholar]
- Myungjin, K.; Jongsik, C. Bacterial community structure in kimchi, a Korean fermented vegetable food, as revealed by 16S rRNA gene analysis. Int. J. Food Microbiol. 2005, 103, 91–96. [Google Scholar]
- Patricia, E.; Alba, P.A.; Alba, Y.; Cecilia, A.O.; Eugenia, J.; Pier Sandro, C.; Graciela, V.; Rosa, A. Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chicha, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 2015, 198, 9–18. [Google Scholar]
- Bjerke, G.A.; Rudi, K.; Avershina, E.; Moen, B.; Blom, H.; Axelsson, L. Exploring the brine microbiota of a traditional norwegian fermented fish product (rakfisk) from six different producers during two consecutive seasonal productions. Foods 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Beddows, G.G.; Ismall, M.; Steinkraus, K.H. The use of bromelain in the hydrolysis of mackerel and the investigation of fermented fish aroma. Int. J. Food Sci Tech. 1976, 11, 379–388. [Google Scholar] [CrossRef]
- Dissaraphong, S.; Benjakul, S.; Visessanguan, W.; Kishimura, H. The influence of storage conditions of tuna viscera before fermentation on the chemical, physical and microbiological changes in fish sauce during fermentation. Bioresour. Technol. 2006, 97, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xia, W.; Jiang, Q.; Guan, L. Biochemical and sensory characteristics of whole carp inoculated with autochthonous starter cultures. J. Aquat. Food Prod. Technol. 2015, 24, 52–67. [Google Scholar] [CrossRef]
- Anne, C.; John, B. Estimating the number of species in a stochastic abundance model. Biometrics 2002, 58, 531–539. [Google Scholar]
- Nicholas, J.G. Ecology. Biodiversity in the scales. Nature 2002, 419, 575–576. [Google Scholar]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. 1994, 34, 101–118. [Google Scholar]
- Anal, A. Quality ingredients and safety concerns for traditional fermented foods and beverages from Asia: A review. Fermentation 2019, 5, 8. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous Food Fermentations and Potential Risks for Human Health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Jeon, C.O. Microbial succession and metabolite changes during fermentation of saeu-jeot: Traditional Korean salted seafood. Food Microbiol. 2013, 34, 360–368. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, Y.J.; Jeon, C.O. Microbial Successions and Metabolite Changes during fermentation of salted shrimp (saeu-jeot) with different salt concentrations. PLoS ONE 2014, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jung, Y.J.; Jeon, C.O. Effects of temperature on microbial succession and metabolite change during saeu-jeot fermentation. Food Microbiol. 2014, 38, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Hu, X.; Li, M.; Miao, J.; Du, J.; Wu, R. Composition and metabolic activities of the bacterial community in shrimp sauce at the flavor-forming stage of fermentation as revealed by metatranscriptome and 16S rRNA gene sequencings. J. Agric. Food Chem. 2016, 64, 2591–2603. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kwak, H.S.; Jung, H.Y.; Kim, S.S. Microbial communities related to sensory attributes in Korean fermented soy bean paste (Doenjang). Food Res. Int. 2016, 89, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Ansaldi, M.; Théraulaz, L.; Baraquet, C.; Panis, G.; Méjean, V. Aerobic TMAO respiration in Escherichia coli. Mol. Microbiol. 2007, 66, 484–494. [Google Scholar] [CrossRef]
- Dos Santos, J.P.; Couillault, C.; Giordano, G.; Mejean, V.; Iobbi-Nivol, C. Molecular analysis of the trimethylamine N-oxide (TMAO) reductase respiratory system from a Shewanella species. J. Mol. Biol. 1998, 284, 421–433. [Google Scholar] [CrossRef]
- Ansaldi, M.; Bordi, C.; Lepelletier, M.; Méjean, V. TorC apocytochrome negatively autoregulates the trimethylamine N-oxide (TMAO) reductase operon in Escherichia coli. Mol. Microbiol. 1999, 33, 284–295. [Google Scholar] [CrossRef] [Green Version]
- James, P.R.; Hardman, S.M.; Patterson, D.L. Osteomyelitis and possible endocarditis secondary to Lactococcus garvieae: A first case report. Postgrad. Med. J. 2000, 76, 301–303. [Google Scholar] [CrossRef]
- Goyache, J.; Goyache, J.; Vela, A.I.; Gibello, A.; Blanco, M.M.; Briones, V.; González, S.; Téllez, S.; Ballesteros, C.; Domínguez, L.; et al. Lactococcus lactis subsp. lactis infection in waterfowl: First confirmation in animals. Emerg. Infect. Dis. 2001, 7, 884–886. [Google Scholar] [CrossRef]
- Udomsil, N.; Rodtong, S.; Tanasupawat, S.; Yongsawatdigul, J. Proteinase-producing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds. Int. J. Food Microbiol. 2010, 141, 186–194. [Google Scholar] [CrossRef]
- Udomsil, N.; Rodtong, S.; Choi, Y.J.; Hua, Y.; Yongsawatdigul, J. Use of Tetragenococcus halophilus as a starter culture for flavor improvement in fish sauce fermentation. J. Agric. Food Chem. 2011, 59, 8401–8408. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Izawa, Y.; Ishii, S.; Takahashi, H.; Torido, Y.; Kimura, B. Suppressive effect of Tetragenococcus halophilus, isolated from fish-nukazuke, on histamine accumulation in salted and fermented fish. Food Chem. 2012, 130, 569–574. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, X.; Wei, L.; Wu, G.; Kumar, A.; Mao, T.; Liu, Z. A cold-adapted, solvent and salt tolerant esterase from marine bacterium Psychrobacter pacificensis. Int. J. Biol. Macromol. 2015, 81, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Von Neubeck, M.; Baur, C.; Krewinkel, M.; Stoeckel, M.; Kranz, B.; Stressler, T.; Fischer, L.; Hinrichs, J.; Scherer, S.; Wenning, M. Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int. J. Food Microbiol. 2015, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.S.; Oh, J.J.; Han, S.; Park, C.; Lee, C.; Choi, Y.E. Complete genome sequence of Pseudoalteromonas sp. MEBiC 03485, isolated from deep-sea sediment. Mar. Genomics 2019, in press. [Google Scholar] [CrossRef]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Uknalis, J.; Boyd, E.F.; Fay, J.P. Mechanisms for Pseudoalteromonas piscicida-induced killing of Vibrios and Mechanisms for Pseudoalteromonas piscicida-induced killing of Vibrios and Other Bacterial Pathogens. Appl. Environ. Microbiol. 2017, 83, 1–17. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (months) | Average content ± SD | ||||
|---|---|---|---|---|---|
| pH | TSN (mg/mL) | AAN (mg/mL) | TMA (μg/mL) | FAA (mg/mL) | |
| 0 | 6.2 ± 0.3 a | 4.7 ± 0.1 d | 2.0 ± 0.1 e | 105 ± 5.5 a | 4.2 ± 0.1 f |
| 3 | 5.9 ± 0.0 b | 8.8 ± 0.0 c | 4.9 ± 0.2 d | 140 ± 12.4 a | 10.0 ± 0.1 e |
| 6 | 6.0 ± 0.2 ab | 12.1 ± 0.2 a | 10.2 ± 0.2 a | 140 ± 0.2 a | 27.1 ± 0.3 b |
| 9 | 5.8 ± 0.1 bc | 8.8 ± 0.1 c | 6.3 ± 0.0 c | 111 ± 0.0 a | 16.1 ± 0.1 d |
| 12 | 5.5 ± 0.0 d | 9.5 ± 0.2 b | 6.1 ± 0.4 c | 104 ± 11.6 a | 24.2 ± 0.2 c |
| 15 | 5.6 ± 0.1 cd | 12.0 ± 0.1 a | 7.7 ± 0.0 b | 94 ± 5.8 a | 30.6 ± 0.2 a |
| Sample | Seq. Num. | Total OTUs | Chao Index | Shannon Index | Simpson Index |
|---|---|---|---|---|---|
| 0M_1 | 44729 | 133 | 237 | 0.480 | 0.833 |
| 0M_2 | 46051 | 163 | 331 | 0.879 | 0.683 |
| 0M_3 | 54946 | 115 | 194 | 0.362 | 0.888 |
| 3M_1 | 53488 | 217 | 323 | 0.781 | 0.782 |
| 3M_2 | 54578 | 251 | 380 | 0.886 | 0.751 |
| 3M_3 | 61583 | 220 | 415 | 1.712 | 0.367 |
| 6M_1 | 55788 | 350 | 461 | 0.778 | 0.794 |
| 6M_2 | 51342 | 379 | 556 | 1.170 | 0.641 |
| 6M_3 | 51945 | 358 | 513 | 0.797 | 0.790 |
| 9M_1 | 70991 | 346 | 517 | 1.577 | 0.358 |
| 9M_2 | 58086 | 392 | 645 | 1.781 | 0.327 |
| 9M_3 | 77459 | 421 | 692 | 1.770 | 0.325 |
| 12M_1 | 41841 | 458 | 604 | 1.625 | 0.557 |
| 12M_2 | 60149 | 486 | 609 | 1.408 | 0.542 |
| 12M_3 | 67121 | 425 | 559 | 1.145 | 0.643 |
| 15M_1 | 68797 | 708 | 801 | 3.559 | 0.091 |
| 15M_2 | 52977 | 662 | 757 | 3.662 | 0.077 |
| 15M_3 | 60803 | 647 | 730 | 3.136 | 0.171 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, F.; Zhang, X.; Gu, H.; Song, J.; Gao, X. Dynamic Changes in the Bacterial Community During the Fermentation of Traditional Chinese Fish Sauce (TCFS) and Their Correlation with TCFS Quality. Microorganisms 2019, 7, 371. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090371
Du F, Zhang X, Gu H, Song J, Gao X. Dynamic Changes in the Bacterial Community During the Fermentation of Traditional Chinese Fish Sauce (TCFS) and Their Correlation with TCFS Quality. Microorganisms. 2019; 7(9):371. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090371
Chicago/Turabian StyleDu, Fangmin, Xiaoyong Zhang, Huarong Gu, Jiajia Song, and Xiangyang Gao. 2019. "Dynamic Changes in the Bacterial Community During the Fermentation of Traditional Chinese Fish Sauce (TCFS) and Their Correlation with TCFS Quality" Microorganisms 7, no. 9: 371. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090371