Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018)

 , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy and Selection Criteria
2.2. Study Selection
2.3. Data Categorization
2.4. Data Analysis
3. Results
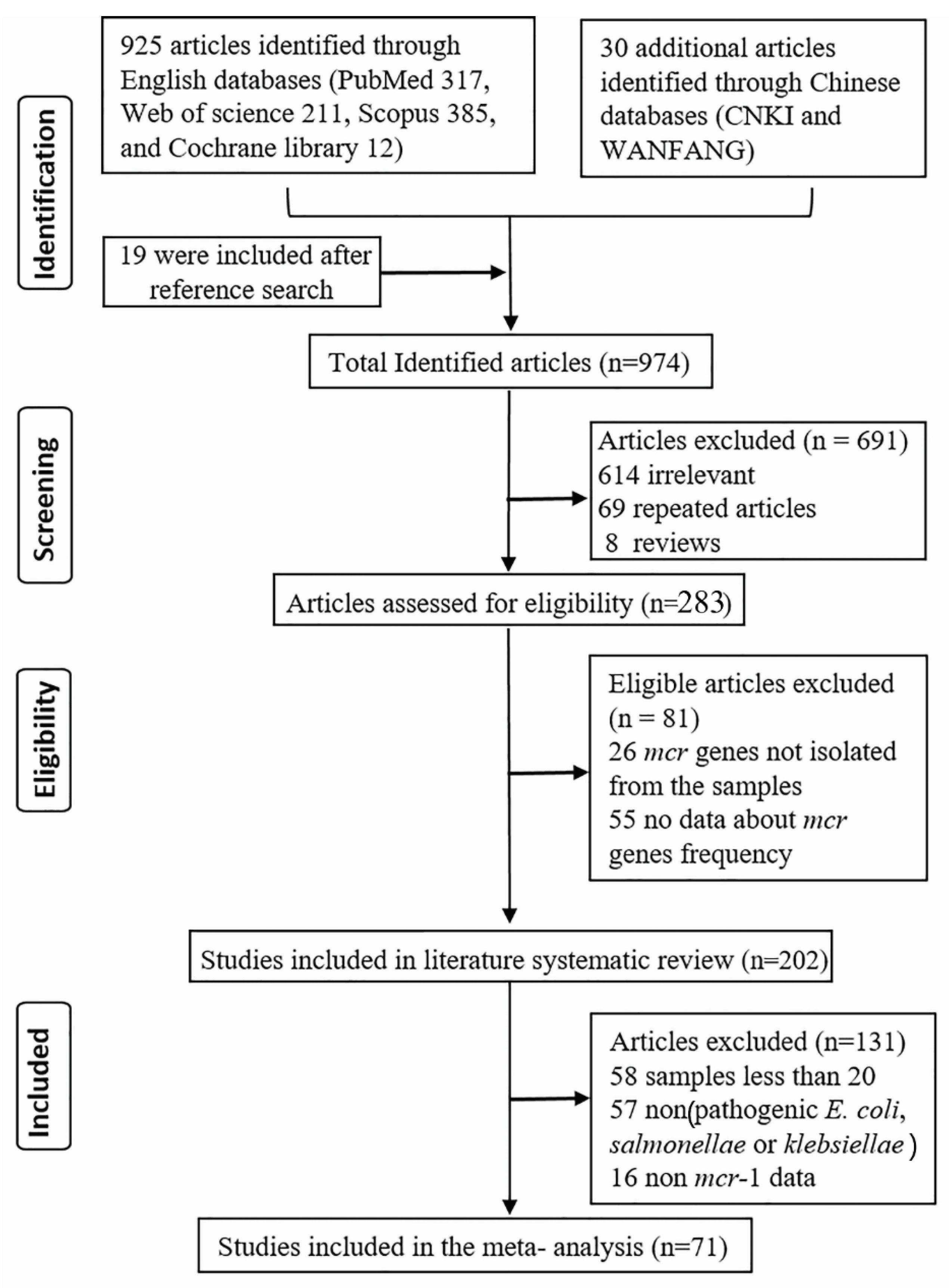
3.1. Data Selection
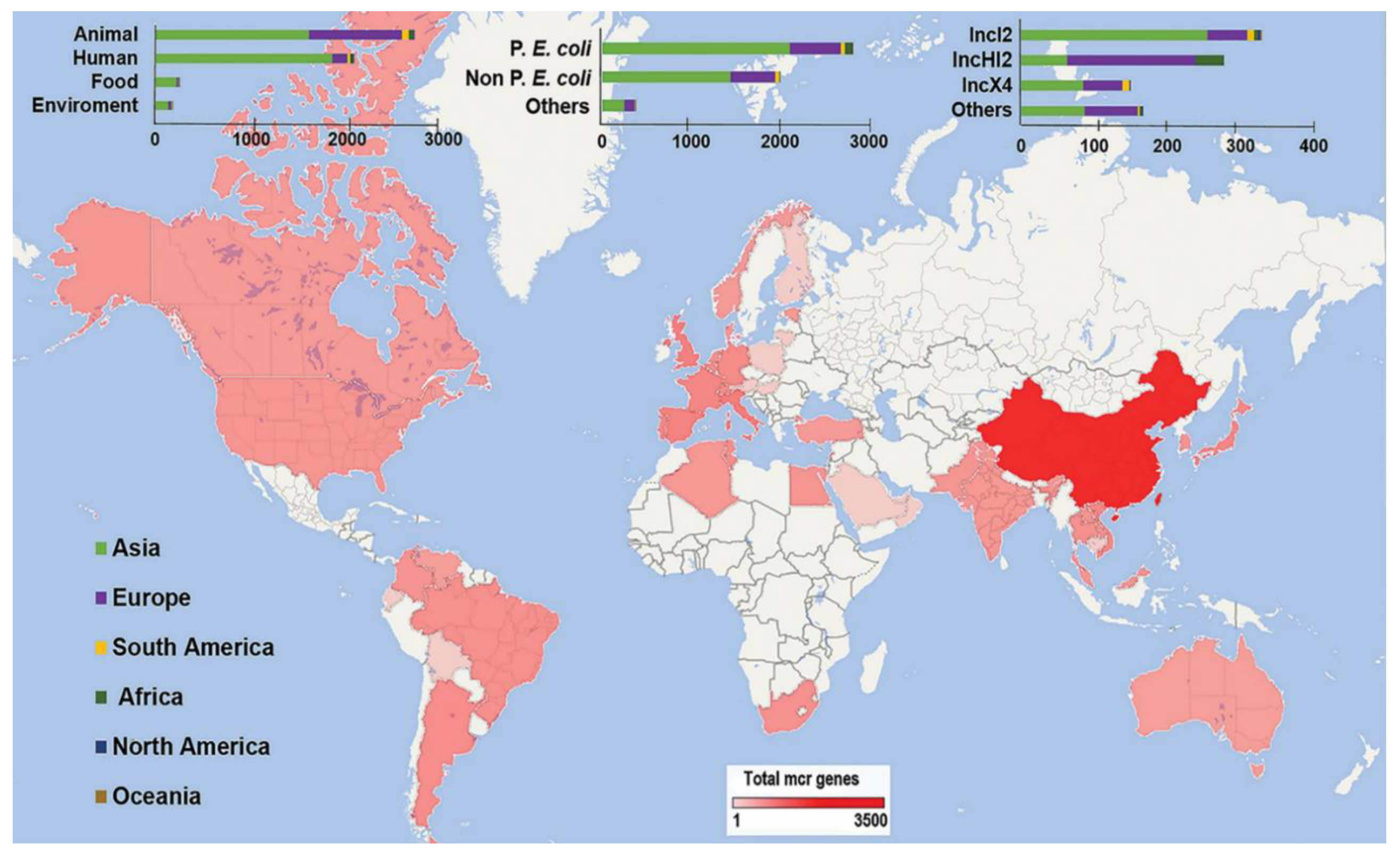
3.2. Global Dissemination of mcr Genes
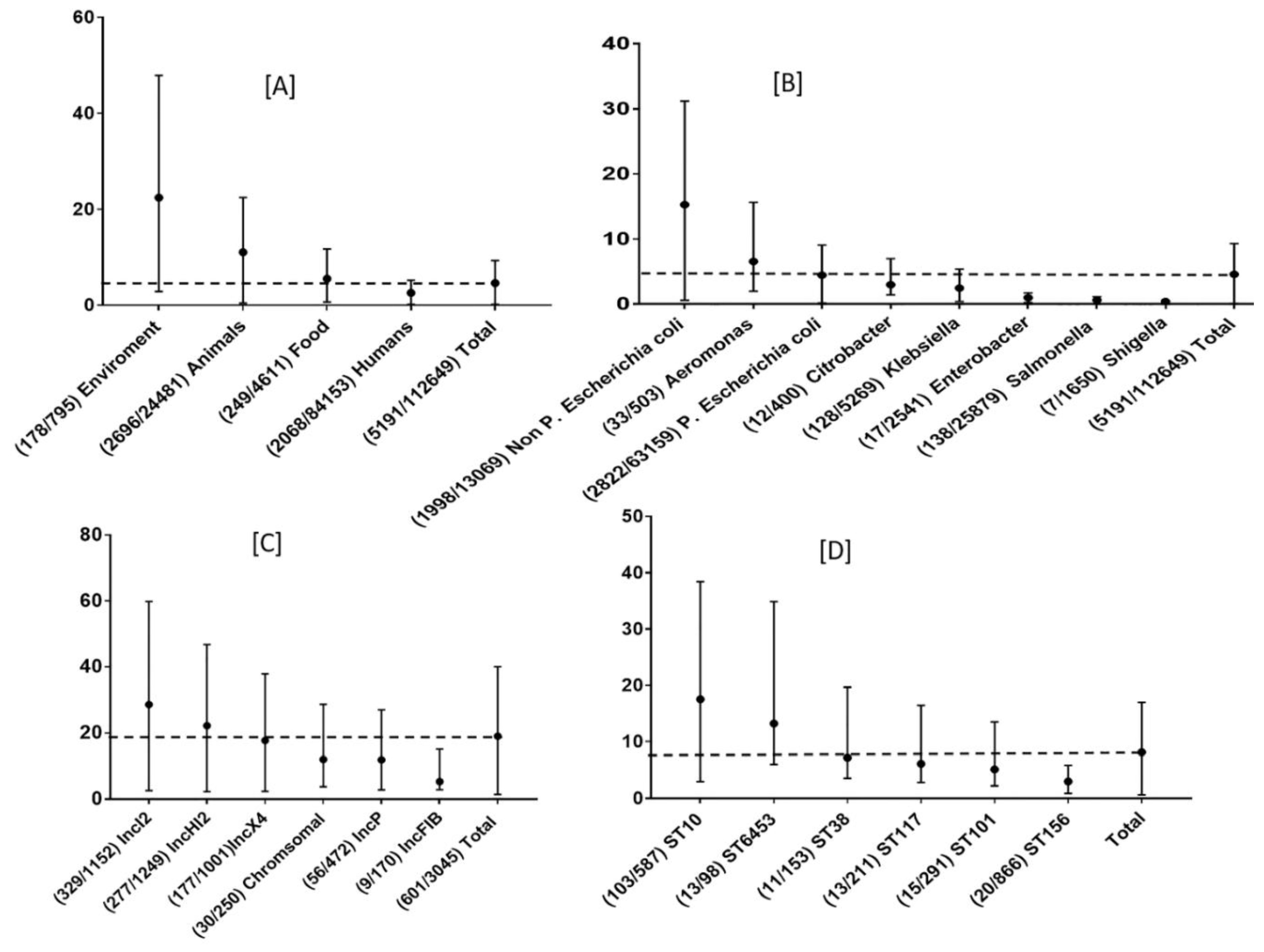
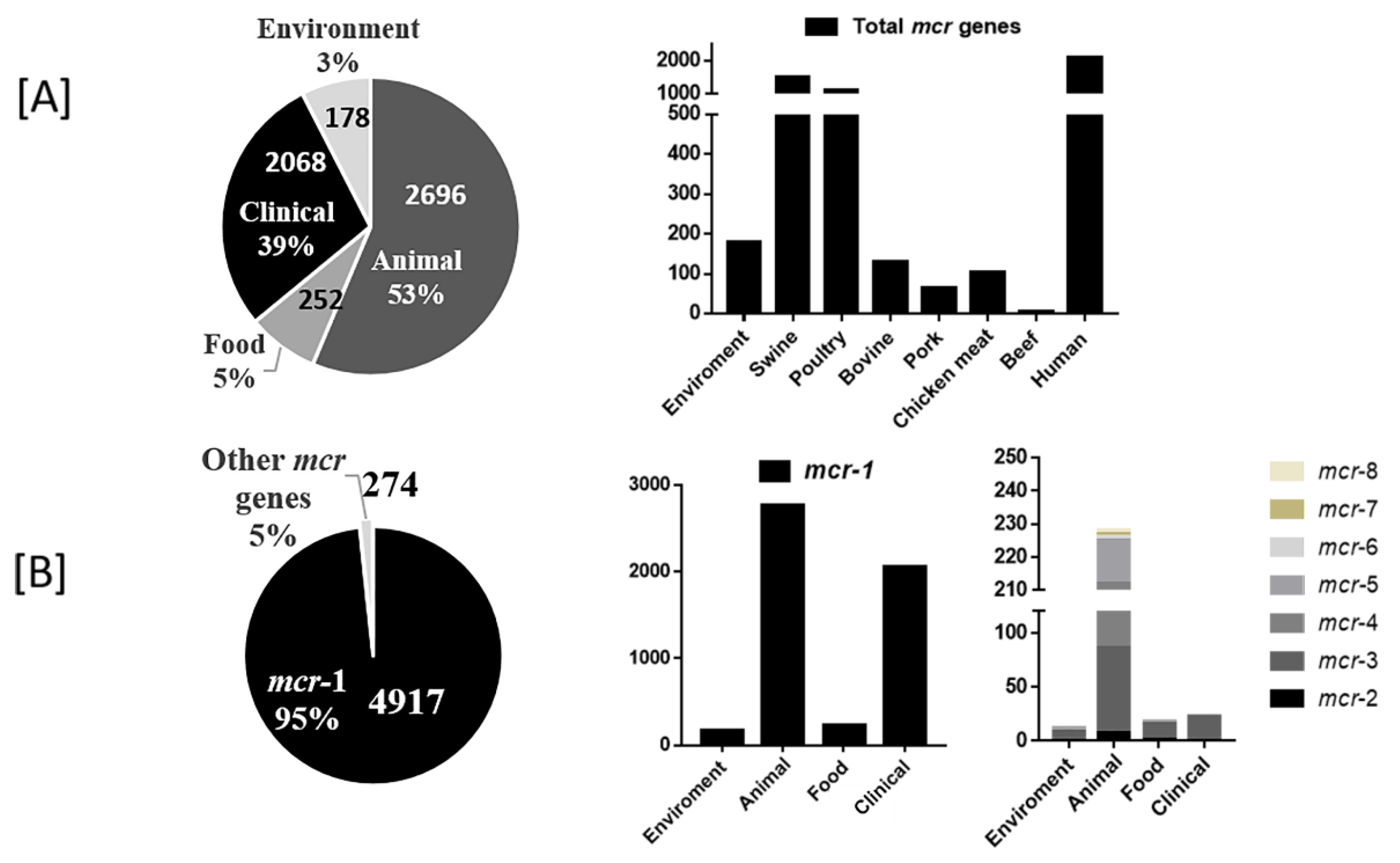
3.3. Global Prevalence of mcr Genes by Sampling Source as a Reservoir
3.4. Global Prevalence of mcr Genes by Bacterial Genera and Species
3.5. Types of mcr Gene and Their Associated Reservoir
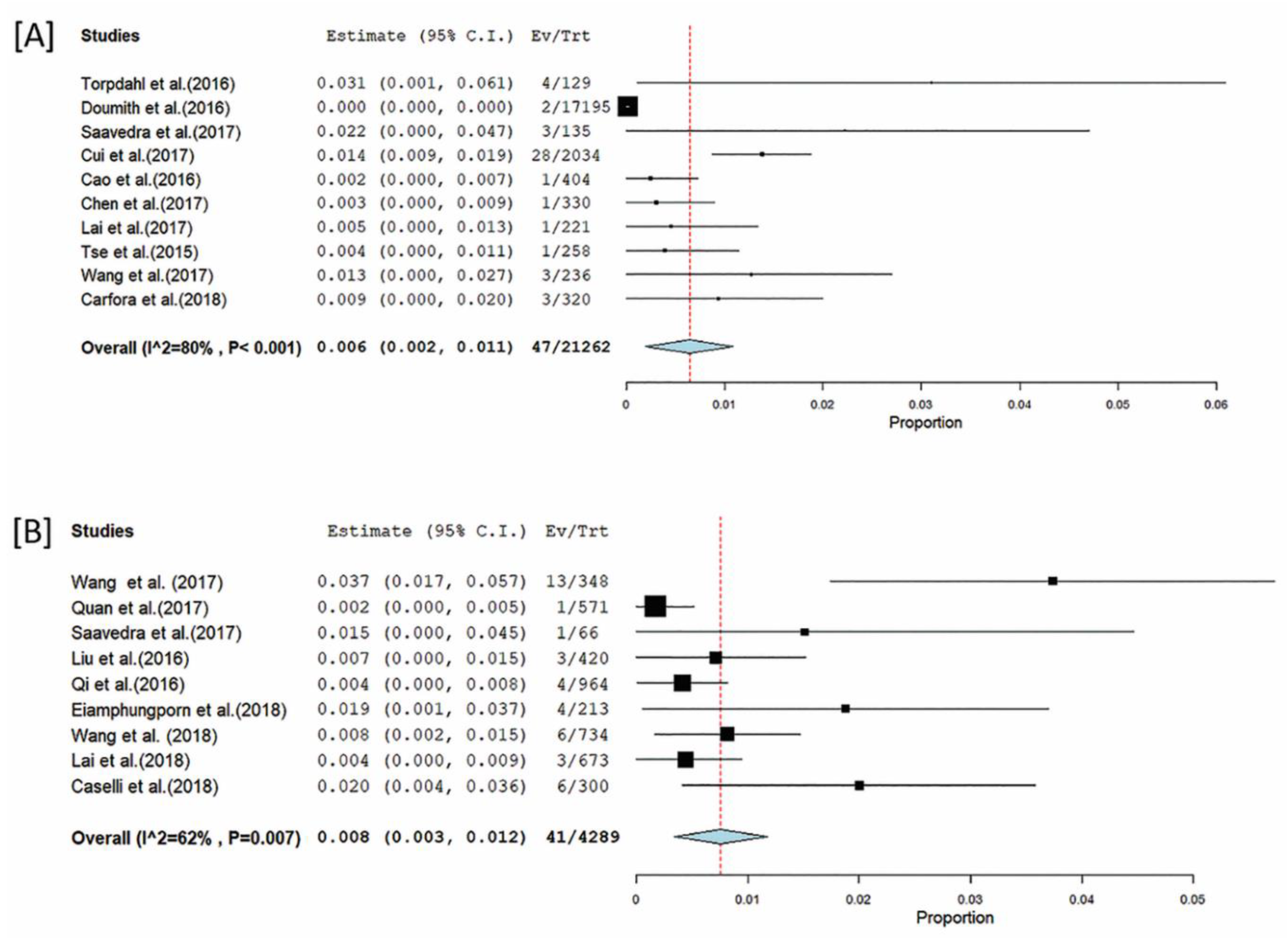
3.6. Antimicrobial Resistance in mcr-Positive Bacteria
3.7. Role of mcr-1 Pathogenic E. coli, Salmonella, and Klebsiella in mcr Gene Dynamics
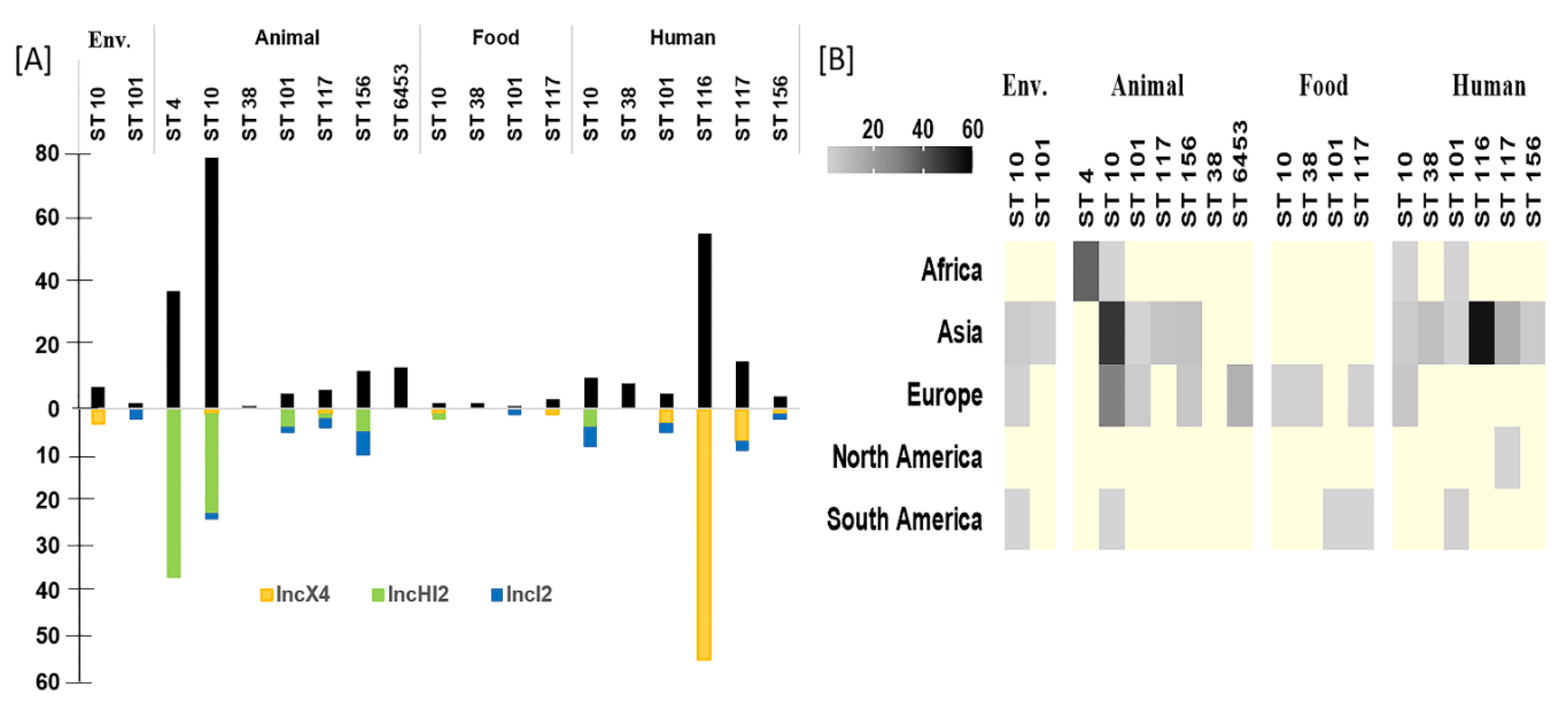
3.8. Common ST of P. E coli with mcr-1
4. Discussion
4.1. Role of mcr-Positive Bacteria via Food-Chain Transmission
4.2. Role of Plasmids in mcr Dissemination
4.3. Role of P. E. coli Genotype for mcr Dissemination
4.4. Rational Antibiotic Choice for mcr-Carrying Bacterial Infections
5. Summary
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Sharing
References
- Nolte, O. Antimicrobial Resistance in the 21st Century: A Multifaceted Challenge. Protein Pept. Lett. 2014, 21, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U. Global antimicrobial resistance in Gram-negative pathogens and clinical need. Curr. Opin. Microbiol. 2017, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Caniaux, I.; van Belkum, A.; Zambardi, G.; Poirel, L.; Gros, M.F. MCR: Modern colistin resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Nation, R.L.; Garonzik, S.M.; Thamlikitkul, V.; Giamarellos-Bourboulis, E.J.; Forrest, A.; Paterson, D.L.; Li, J.; Silveira, F.P. Dosing guidance for intravenous colistin in critically-ill patients. Clin. Infect. Dis. 2017, 64, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to polymyxins in Gram-negative organisms. Int. J. Antimicrob. Agents 2017, 49, 526–535. [Google Scholar] [CrossRef]
- Juhasz, E.; Ivan, M.; Pinter, E.; Pongracz, J.; Kristof, K. Colistin resistance among blood culture isolates at a tertiary care centre in Hungary. J. Glob. Antimicrob. Resist. 2017, 11, 167–170. [Google Scholar] [CrossRef]
- Stoesser, N.; Mathers, A.J.; Moore, C.E.; Day, N.P.; Crook, D.W. Colistin resistance gene mcr-1 and pHNSHP45 plasmid in human isolates of Escherichia coli and Klebsiella pneumoniae. Lancet Infect. Dis. 2016, 16, 285–286. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Shen, Y.; Shen, J.; Wu, C. Early emergence of mcr-1 in Escherichia coli from food-producing animals. Lancet Infect. Dis. 2016, 16, 293. [Google Scholar] [CrossRef]
- Georgios, M.; Lemonia, S. Polymyxin Resistance Mechanisms: From Intrinsic Resistance to Mcr Genes. Recent Pat. Anti Infect. Drug Discov. 2018, 13, 1–9. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.H. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef]
- MacNair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming mcr-1 mediated colistin resistance with colistin in combination with other antibiotics. Nat. Commun. 2018, 9, 458. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Ann. Intern. Med. 2009, 151, 264–269. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC); Division of Bacterial and Mycotic Diseases (DBMD). “Escherichia coli”. Available online: www.cdc.gov/ecoli/ (accessed on 15 March 2019).
- Ryan, K.J.; Ray, C.G.; Ahmad, N.; Drew, W.L.; Lagunoff, M.; Pottinger, P.; Reller, L.B.; Sterling, C.R. “Pathogenesis of Bacterial Infections”. Sherris Medical Microbiology, 6th ed.; McGraw Hill: New York, NY, USA, 2014; pp. 391–406. [Google Scholar]
- Higgins, J.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions Version 5.1.0 [Updated March 2011]; The Cochrane Collaboration: London, UK, 2011; Available online: www.handbook.cochrane.org (accessed on 22 July 2018).
- Payne, M.; Croxen, M.A.; Lee, T.D.; Mayson, B.; Champagne, S.; Leung, V.; Bariso, S.; Hoang, L.; Lowe, C. mcr-1-Positive Colistin-Resistant Escherichia coli in Traveler Returning to Canada from China. Emerg. Infect. Dis. 2016, 22, 1673–1675. [Google Scholar] [CrossRef]
- Macesic, N.; Green, D.; Wang, Z.; Sullivan, S.B.; Shim, K.; Park, S.; Whittier, S.; Furuya, E.Y.; Gomez-Simmonds, A.; Uhlemann, A.C. Detection of mcr-1-Carrying Escherichia coli Causing Bloodstream Infection in a New York City Hospital: Avian Origins, Human Concerns? Open Forum Infect. Dis. 2017, 4, ofx115. [Google Scholar] [CrossRef]
- Tada, T.; Uechi, K.; Nakasone, I.; Shimada, K.; Nakamatsu, M.; Kirikae, T.; Fujita, J. Emergence of a colistin-resistant Escherichia coli clinical isolate harboring mcr-1 in Japan. Int. J. Infect. Dis. 2017, 63, 21–22. [Google Scholar] [CrossRef]
- Nishino, Y.; Shimojima, Y.; Suzuki, Y.; Ida, M.; Fukui, R.; Kuroda, S.; Hirai, A.; Sadamasu, K. Detection of the mcr-1 gene in colistin resistant Escherichia coli from retail meat in Japan. Microbiol. Immunol. 2017, 61, 554–557. [Google Scholar] [CrossRef]
- Grami, R.; Mansour, W.; Mehri, W.; Bouallegue, O.; Boujaafar, N.; Madec, J.Y.; Haenni, M. Impact of food animal trade on the spread of mcr-1-mediated colistin resistance, Tunisia, July 2015. Eurosurveillance 2016, 21, 30144. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, M.; Faccone, D.; Pasteran, F.; Ceriana, P.; Albornoz, E.; Petroni, A.; Corso, A. First Description of mcr-1-Mediated Colistin Resistance in Human Infections Caused by Escherichia coli in Latin America. Antimicrob. Agents Chemother. 2016, 60, 4412–4413. [Google Scholar] [CrossRef] [PubMed]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, I.; Feudi, C.; Wang, Y.; Kaspar, H.; Feßler, A.T.; Lübke-Becker, A.; Michael, G.B.; Shen, J.; Schwarz, S. Identification of novel variants of the colistin resistance gene mcr-3 in Aeromonas spp. from the national resistance monitoring programme GERM-Vet and from diagnostic submissions. J. Antimicrob. Chemother. 2018, 73, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-Q.; Li, Y.-X.; Lei, C.-W.; Zhang, A.-Y.; Wang, H.-N. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef] [PubMed]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Eur. Commun. Dis. Bull. 2016, 21. [Google Scholar] [CrossRef]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi, B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef]
- Carattoli, A.; Carretto, E.; Brovarone, F.; Sarti, M.; Villa, L. Comparative analysis of an mcr-4 Salmonella enterica subsp. enterica monophasic variant of human and animal origin. J. Antimicrob. Chemother. 2018, 73, 3332–3335. [Google Scholar] [CrossRef]
- Hernandez, M.; Iglesias, M.R.; Rodriguez-Lazaro, D.; Gallardo, A.; Quijada, N.; Miguela-Villoldo, P.; Campos, M.J.; Piriz, S.; Lopez-Orozco, G.; de Frutos, C.; et al. Co-occurrence of colistin-resistance genes mcr-1 and mcr-3 among multidrug-resistant Escherichia coli isolated from cattle, Spain, September 2015. Eur. Commun. Dis. Bull. 2017, 22. [Google Scholar] [CrossRef]
- Litrup, E.; Kiil, K.; Hammerum, A.M.; Roer, L.; Nielsen, E.M.; Torpdahl, M. Plasmid-borne colistin resistance gene mcr-3 in Salmonella isolates from human infections, Denmark, 2009–17. Eurosurveillance 2017, 22, 30587. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Graells, C.; De Keersmaecker, S.C.J.; Vanneste, K.; Pochet, B.; Vermeersch, K.; Roosens, N.; Dierick, K.; Botteldoorn, N. Detection of Plasmid-Mediated Colistin Resistance, mcr-1 and mcr-2 Genes, in Salmonella spp. Isolated from Food at Retail in Belgium from 2012 to 2015. Foodborne Pathog. Dis. 2017, 15, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Nordmann, P.; Poirel, L. Moraxella Species as Potential Sources of MCR-Like Polymyxin Resistance Determinants. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Li, Y.; Elbediwi, M.; Yue, M. Emergence and Dissemination of mcr-Carrying Clinically Relevant Salmonella Typhimurium Monophasic Clone ST34. Microorganisms 2019, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Kieffer, N.; Fernandez-Garayzabal, J.F.; Vela, A.I.; Larpin, Y.; Nordmann, P. MCR-2-mediated plasmid-borne polymyxin resistance most likely originates from Moraxella pluranimalium. J. Antimicrob. Chemother. 2017, 72, 2947–2949. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.T.; Song, F.J.; Zou, M.; Zhang, Q.D.; Shan, H. High Incidence of Escherichia coli Strains Coharboring mcr-1 and blaNDM from Chickens. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Kong, L.H.; Lei, C.W.; Ma, S.Z.; Jiang, W.; Liu, B.H.; Wang, Y.X.; Guan, R.; Men, S.; Yuan, Q.W.; Cheng, G.Y.; et al. Various Sequence Types of Escherichia coli Isolates Coharboring blaNDM-5 and mcr-1 Genes from a Commercial Swine Farm in China. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Jiang, Z.; Paudyal, N.; Xu, Y.; Deng, T.; Li, F.; Pan, H.; Peng, X.; He, Q.; Yue, M. Antibiotic Resistance Profiles of Salmonella Recovered From Finishing Pigs and Slaughter Facilities in Henan, China. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic Resistance in Salmonella Typhimurium Isolates Recovered From the Food Chain Through National Antimicrobial Resistance Monitoring System Between 1996 and 2016. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Pan, H.; Paudyal, N.; Li, X.; Fang, W.; Yue, M. Multiple Food-Animal-Borne Route in Transmission of Antibiotic-Resistant Salmonella Newport to Humans. Front. Microbiol. 2018, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caltagirone, M.; Nucleo, E.; Spalla, M.; Zara, F.; Novazzi, F.; Marchetti, V.M.; Piazza, A.; Bitar, I.; De Cicco, M.; Paolucci, S.; et al. Occurrence of Extended Spectrum β-Lactamases, KPC-Type, and MCR-1.2-Producing Enterobacteriaceae from Wells, River Water, and Wastewater Treatment Plants in Oltrepò Pavese Area, Northern Italy. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Sellera, F.P.; Esposito, F.; Sabino, C.P.; Cerdeira, L.; Lincopan, N. Colistin-Resistant mcr-1-Positive Escherichia coli on Public Beaches, an Infectious Threat Emerging in Recreational Waters. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Hartl, R.; Kerschner, H.; Lepuschitz, S.; Ruppitsch, W.; Allerberger, F.; Apfalter, P. Detection of the mcr-1 Gene in a Multidrug-Resistant Escherichia coli Isolate from an Austrian Patient. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Shen, Y.; Zhou, H.; Xu, J.; Wang, Y.; Zhang, Q.; Walsh, T.R.; Shao, B.; Wu, C.; Hu, Y.; Yang, L.; et al. Anthropogenic and environmental factors associated with high incidence of mcr-1 carriage in humans across China. Nat. Microbiol. 2018, 3, 1054–1062. [Google Scholar] [CrossRef]
- Paudyal, N.; Yue, M. Antimicrobial Resistance in the “Dark Matter”. Clin. Infect. Dis. 2019, 69, 379–380. [Google Scholar] [CrossRef]
- Pan, H.; Zhou, X.; Chai, W.; Paudyal, N.; Li, S.; Zhou, X.; Zhou, K.; Wu, Q.; Wu, B.; Li, G.; et al. Diversified sources for human infections by Salmonella enterica serovar newport. Transbound. Emerg. Dis. 2019, 66, 1044–1048. [Google Scholar] [CrossRef]
- Pan, H.; Zhou, X.; Chai, W.; Paudyal, N.; Li, S.; Zhou, X.; Zhou, K.; Wu, Q.; Wu, B.; Li, G.; et al. Cover Image. Transbound. Emerg. Dis. 2019, 66. [Google Scholar] [CrossRef]
- Ellem, J.A.; Ginn, A.N.; Chen, S.C.; Ferguson, J.; Partridge, S.R.; Iredell, J.R. Locally Acquired mcr-1 in Escherichia coli, Australia, 2011 and 2013. Emerg. Infect. Dis. 2017, 23, 1160–1163. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, G.B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.Y.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef]
- Tijet, N.; Faccone, D.; Rapoport, M.; Seah, C.; Pasteran, F.; Ceriana, P.; Albornoz, E.; Corso, A.; Petroni, A.; Melano, R.G. Molecular characteristics of mcr-1-carrying plasmids and new mcr-1 variant recovered from polyclonal clinical Escherichia coli from Argentina and Canada. PLoS ONE 2017, 12, e0180347. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Partridge, S.R.; He, L.; Zeng, Z.; He, D.; Ye, J.; Liu, J.-H. Genetic Characterization of IncI2 Plasmids Carrying blaCTX-M-55 Spreading in both Pets and Food Animals in China. Antimicrob. Agents Chemother. 2013, 57, 2824–2827. [Google Scholar] [CrossRef] [PubMed]
- Tato, M.; Coque, T.M.; Baquero, F.; Canton, R. Dispersal of carbapenemase blaVIM-1 gene associated with different Tn402 variants, mercury transposons, and conjugative plasmids in Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2010, 54, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Wannemeuhler, Y.M.; Scaccianoce, J.A.; Johnson, S.J.; Nolan, L.K. Complete DNA sequence, comparative genomics, and prevalence of an IncHI2 plasmid occurring among extraintestinal pathogenic Escherichia coli isolates. Antimicrob. Agents Chemother. 2006, 50, 3929–3933. [Google Scholar] [CrossRef]
- Poirel, L.; Kieffer, N.; Nordmann, P. In Vitro Study of ISApl1-Mediated Mobilization of the Colistin Resistance Gene mcr-1. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Snesrud, E.; He, S.; Chandler, M.; Dekker, J.P.; Hickman, A.B.; McGann, P.; Dyda, F. A Model for Transposition of the Colistin Resistance Gene mcr-1 by ISApl1. Antimicrob. Agents Chemother. 2016, 60, 6973–6976. [Google Scholar] [CrossRef]
- Snesrud, E.; McGann, P.; Chandler, M. The Birth and Demise of the ISApl1-mcr-1-ISApl1 Composite Transposon: The Vehicle for Transferable Colistin Resistance. mBio 2018, 9. [Google Scholar] [CrossRef]
- Zurfluh, K.; Nüesch-Inderbinen, M.; Klumpp, J.; Poirel, L.; Nordmann, P.; Stephan, R. Key features of mcr-1-bearing plasmids from Escherichia coli isolated from humans and food. Antimicrob. Resist. Infect. Control 2017, 6, 91. [Google Scholar] [CrossRef]
- Aibinu, I.; Odugbemi, T.; Koenig, W.; Ghebremedhin, B. Sequence Type ST131 and ST10 Complex (ST617) predominant among CTX-M-15-producing Escherichia coli isolates from Nigeria. Clin. Microbiol. Infect. 2012, 18, E49–E51. [Google Scholar] [CrossRef]
- Oteo, J.; Diestra, K.; Juan, C.; Bautista, V.; Novais, Â.; Pérez-Vázquez, M.; Moyá, B.; Miró, E.; Coque, T.M.; Oliver, A.; et al. Extended-spectrum β-lactamase-producing Escherichia coli in Spain belong to a large variety of multilocus sequence typing types, including ST10 complex/A, ST23 complex/A and ST131/B2. Int. J. Antimicrob. Agents 2009, 34, 173–176. [Google Scholar] [CrossRef]
- Guillouzouic, A.; Caroff, N.; Dauvergne, S.; Lepelletier, D.; Perrin Guyomard, A.; Kempf, I.; Reynaud, A.; Corvec, S. MLST typing of Escherichia coli isolates overproducing AmpC beta-lactamase. J. Antimicrob. Chemother. 2009, 63, 1290–1292. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Saralaya, V.; Adhikari, P.; Shenoy, S.; Baliga, S.; Hegde, A. Characterization of Escherichia coli Phylogenetic Groups Associated with Extraintestinal Infections in South Indian Population. Ann. Med Health Sci. Res. 2015, 5, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Nunez, B.; Avila, P.; de la Cruz, F. Genes involved in conjugative DNA processing of plasmid R6K. Mol. Microbiol. 1997, 24, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.K.M.; De Masi, L.; Yue, M.; Galván, E.M.; Chen, H.; Wang, F.; Schifferli, D.M. Adhesive Properties of YapV and Paralogous Autotransporter Proteins of Yersinia pestis. Infect. Immun. 2015, 83, 1809. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.S.; Matamouros, S.; Hager, K.R.; Brittnacher, M.J.; Rohmer, L.; Radey, M.C.; Weiss, E.J.; Kim, K.B.; Jacobs, M.A.; Sims-Day, E.H.; et al. Genomic Analysis of Salmonella enterica Serovar Typhimurium Characterizes Strain Diversity for Recent, U.S. Salmonellosis Cases and Identifies Mutations Linked to Loss of Fitness under Nitrosative and Oxidative Stress. mBio 2016, 7, e00154-16. [Google Scholar] [CrossRef]
- Yue, M.; Han, X.; Masi, L.D.; Zhu, C.; Ma, X.; Zhang, J.; Wu, R.; Schmieder, R.; Kaushik, R.S.; Fraser, G.P.; et al. Allelic variation contributes to bacterial host specificity. Nat. Commun. 2015, 6, 8754. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Moura, Q.; Sartori, L.; Silva, K.C.; Cunha, M.P.; Esposito, F.; Lopes, R.; Otutumi, L.K.; Goncalves, D.D.; Dropa, M.; et al. Silent dissemination of colistin-resistant Escherichia coli in South America could contribute to the global spread of the mcr-1 gene. Eur. Commun. Dis. Bull. 2016, 21. [Google Scholar] [CrossRef]
- Pham Thanh, D.; Thanh Tuyen, H.; Nguyen Thi Nguyen, T.; Chung The, H.; Wick, R.R.; Thwaites, G.E.; Baker, S.; Holt, K.E. Inducible colistin resistance via a disrupted plasmid-borne mcr-1 gene in a 2008 Vietnamese Shigella sonnei isolate. J. Antimicrob. Chemother. 2016, 71, 2314–2317. [Google Scholar] [CrossRef]
- Liassine, N.; Assouvie, L.; Descombes, M.-C.; Tendon, V.D.; Kieffer, N.; Poirel, L.; Nordmann, P. Very low prevalence of MCR-1/MCR-2 plasmid-mediated colistin resistance in urinary tract Enterobacteriaceae in Switzerland. Int. J. Infect. Dis. 2016, 51, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.; Li, X.; Chen, Y.; Jiang, Y.; Zhou, Z.; Zhang, H.; Sun, L.; Ruan, Z.; Feng, Y.; Akova, M.; et al. Prevalence of mcr-1 in Escherichia coli and Klebsiella pneumoniae recovered from bloodstream infections in China: A multicentre longitudinal study. Lancet Infect. Dis. 2017, 17, 400–410. [Google Scholar] [CrossRef]
- Di Pilato, V.; Arena, F.; Tascini, C.; Cannatelli, A.; Henrici De Angelis, L.; Fortunato, S.; Giani, T.; Menichetti, F.; Rossolini, G.M. mcr-1.2, a New mcr Variant Carried on a Transferable Plasmid from a Colistin-Resistant KPC Carbapenemase-Producing Klebsiella pneumoniae Strain of Sequence Type 512. Antimicrob. Agents Chemother. 2016, 60, 5612–5615. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.-T.; Yang, R.-S.; Liao, X.-P.; Liu, Y.-H.; Zhou, Y.-F.; Sun, J.; Feng, Y. Increased activity of colistin in combination with amikacin against Escherichia coli co-producing NDM-5 and MCR-1. J. Antimicrob. Chemother. 2017, 72, 1723–1730. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Includes | Sub-Includes |
|---|---|---|
| Environmental | Sewage, river, and sea water samples | |
| Animal | Pig, poultry, cattle, and other animal samples | |
| Poultry | Chicken, turkey, and muscovy duck | |
| Other animals | Migratory birds, penguins, reptiles, kelp gulls and dogs | |
| Food | Pork, chicken meat, beef, and other foods samples | |
| Other foods | Vegetables and foods from unknown sources | |
| Human | Fecal and other samples | |
| Other samples | Blood, urine, pus, sperm, abdominal and peritoneal fluids, sputum, and ascites | |
| Harboring bacteria | Pathogenic E. coli (P. E. coli), Salmonella spp., Klebsiella pneumoniae, other bacteria | |
| Salmonella spp. | S. Typhimurium, S. Paratyphi B, S. Virchow, S. Rissen, S. Indiana and S. Derby, S. London | |
| Other bacteria | Citrobacter freundii, non-pathogenic. E. coli, Citrobacter braakii, Enterobacter cloacae, Shigella sonnei, Aeromonas veronii, Enterobacter aerogenes, Cronobacter sakazakii, Kluyvera ascorbata, Acinetobacter, Klebsiella variicola, Moraxella pluranimalium, Klebsiella oxytoca, Providencia alcalifaciens, and Raoultella ornithinolytica | |
| Plasmid types | IncI2, IncHI2, IncX4 | |
| Other plasmids | IncF, IncFIB, Inck, Incx, IncP, IncFII, and ColE |
| Sources | CST | TGC | GEN | CIP | AMK | CTX | CFX | TET | TMP-SMX | FOS | MEM | IMP | AMP | KAN | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | P. E. coli | 98.69% (993) | 3.39% (59) | 59.55% (178) | 44.09% (973) | 22.60% (146) | 79.61% (152) | 83.72% (43) | 96.43% (28) | 83.08% (65) | 48.62% (109) | 2.76% (979) | 2.76% (979) | 93.33% (150) | 95.00% (20) |
| Swine | 99.37% (158) | 100.00% (3) | 86.67% (15) | 90.91% (11) | 6.06% (33) | 47.06% (34) | 100.00% (3) | 100.00% 98) | 98.98% (98) | 100.00% (27) | 43.64% (55) | 40.38% (52) | 43.75% (48) | 80.65% (33) | |
| Poultry | 100.00% (125) | 0.00% (38) | 61.90% (21) | 97.58% (124) | 16.67% (18) | 100.00% (55) | 100.00% (3) | 96.43% (56) | 98.04% (51) | 100.00% (54) | 0.00% (3) | 0.00% (3) | 94.59% (74) | 26.32% (19) | |
| Bovine | 100.00% (17) | 0% (5) | 80.00% (5) | 100.00% (6) | NA | 83.33% (6) | 83.33% (6) | 100.00% (6) | 100.00% (5) | 0.00% (5) | 0.00% (1) | 0.00% (1) | 100.00% (6) | 100.00% (1) | |
| Food | 97.83% (45) | 100.00% (1) | 33.33% (18) | 69.57% (7) | 100.00% (1) | 81.25% (16) | 100.00% (5) | 84.21% (19) | 87.50% (24) | 100.00% (3) | 0.00% (7) | 0.00% (7) | 81.25% (16) | 40.00% (5) | |
| Environmental | 87.10% (31) | 33.33% (24) | 60.00% (5) | 36.00% (25) | 72.73% (11) | 100.00% (1) | 100.00% (2) | 100.00% (3) | 100.00% (1) | 100.00% (1) | 9.09% (30) | 9.09% (33) | 100.00% (3) | NA | |
| Human | K. pneumoniae | 100.00% (36) | 21.40% (7) | 91.67% (12) | 63.64% (11) | 33.33% (6) | 91.67% (12) | 100.00% (6) | 100.00% (4) | 60.00% (5) | 66.67% (3) | 53.85% (13) | 53.85% (13) | 90.00% (10) | 33.3% (6) |
| Animals | 100.00% (13) | NA | 100.00% (4) | 100.00% (4) | 100.00% (1) | 100.00% (4) | 100.00% (4) | 91.67% (11) | 100.00% (9) | NA | 0.00% (5) | 0.00% (5) | 100.00% (4) | 0.00% (1) | |
| Human | Salmonella | 100.00% (32) | NA | 43.59% (39) | 24.14% (29) | NA | 65.52% (29) | 100.00% (1) | 80.65% (31) | 63.16% (19) | NA | 0.00% (2) | 0.00% (2) | 90.00% (30) | 90.00% (30) |
| Animals | 100.00% (36) | NA | 96.55% (29) | 97.30% (36) | 96.4% (26) | 87.50% (8) | 0.00% (36) | 100.00% (10) | 100.00% (34) | 96.15% (26) | 0.00% (2) | 0.00% (2) | 100.00% (54) | 100.00% (3) | |
| Human | Other isolates | 60.00% (5) | 100.00% (1) | 100.00% (3) | 75.00% (4) | 66.67% (3) | 100.00% (1) | 0.00% (2) | 100.00% (2) | NA | NA | 14.2% (6) | 75.00% (5) | 100.00% (2) | NA |
| Animals | 96.34% (273) | 0.00% (152) | 31.84% (223) | 76.50% (234) | 1.94% (155) | 93.33% (165) | 99.34% (152) | 96.41% (232) | NA | 100.00% (3) | 0.86% (233) | 48.07% (231) | 100.00% (71) | 100.00% (3) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbediwi, M.; Li, Y.; Paudyal, N.; Pan, H.; Li, X.; Xie, S.; Rajkovic, A.; Feng, Y.; Fang, W.; C. Rankin, S.; et al. Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018). Microorganisms 2019, 7, 461. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100461
Elbediwi M, Li Y, Paudyal N, Pan H, Li X, Xie S, Rajkovic A, Feng Y, Fang W, C. Rankin S, et al. Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018). Microorganisms. 2019; 7(10):461. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100461
Chicago/Turabian StyleElbediwi, Mohammed, Yan Li, Narayan Paudyal, Hang Pan, Xiaoliang Li, Shaohua Xie, Andreja Rajkovic, Youjun Feng, Weihuan Fang, Shelley C. Rankin, and et al. 2019. "Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018)" Microorganisms 7, no. 10: 461. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7100461