Biodiversity among Brettanomyces bruxellensis Strains Isolated from Different Wine Regions of Chile: Key Factors Revealed about Its Tolerance to Sulphite

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Selection and Cell Maintenance
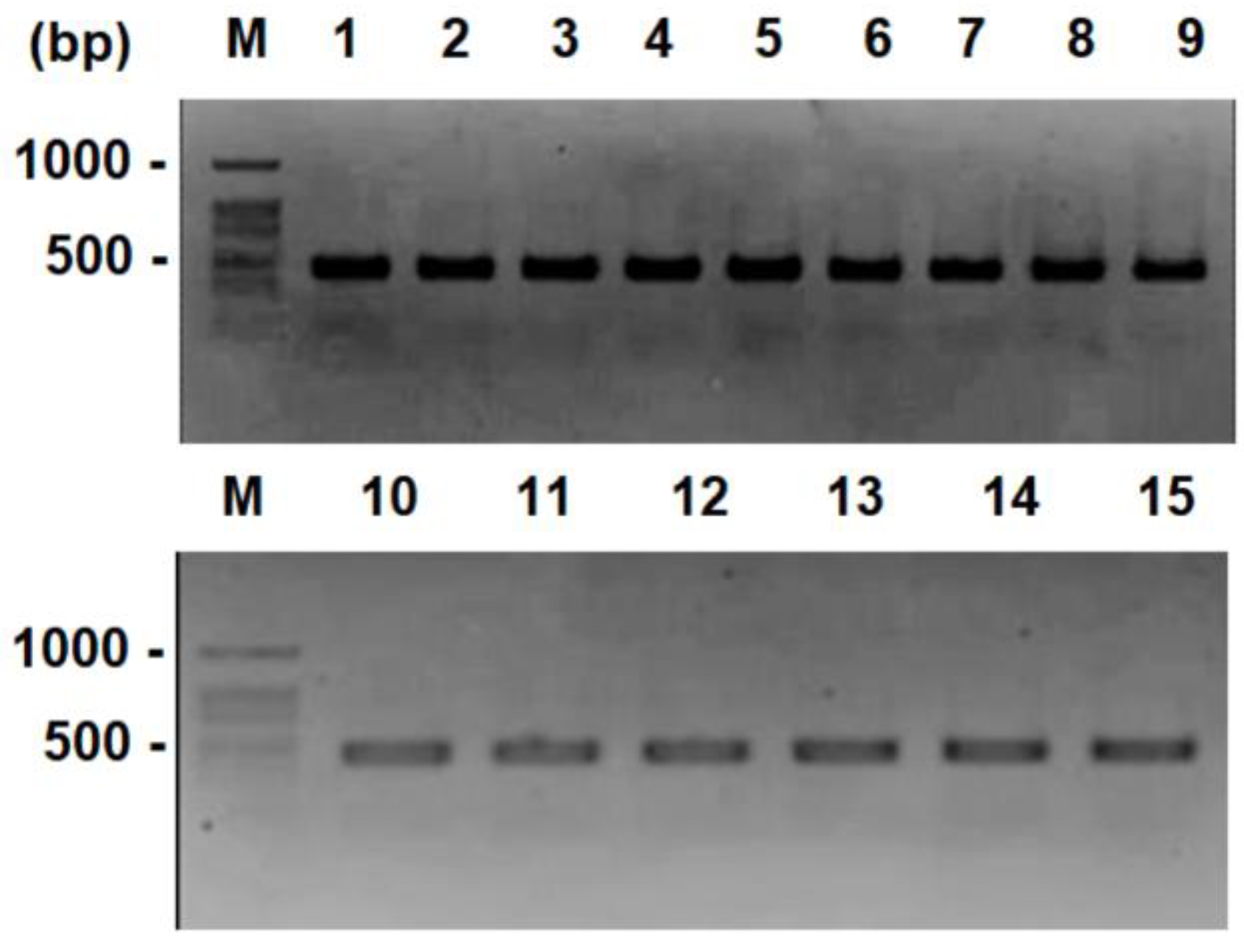
2.2. Molecular Identification
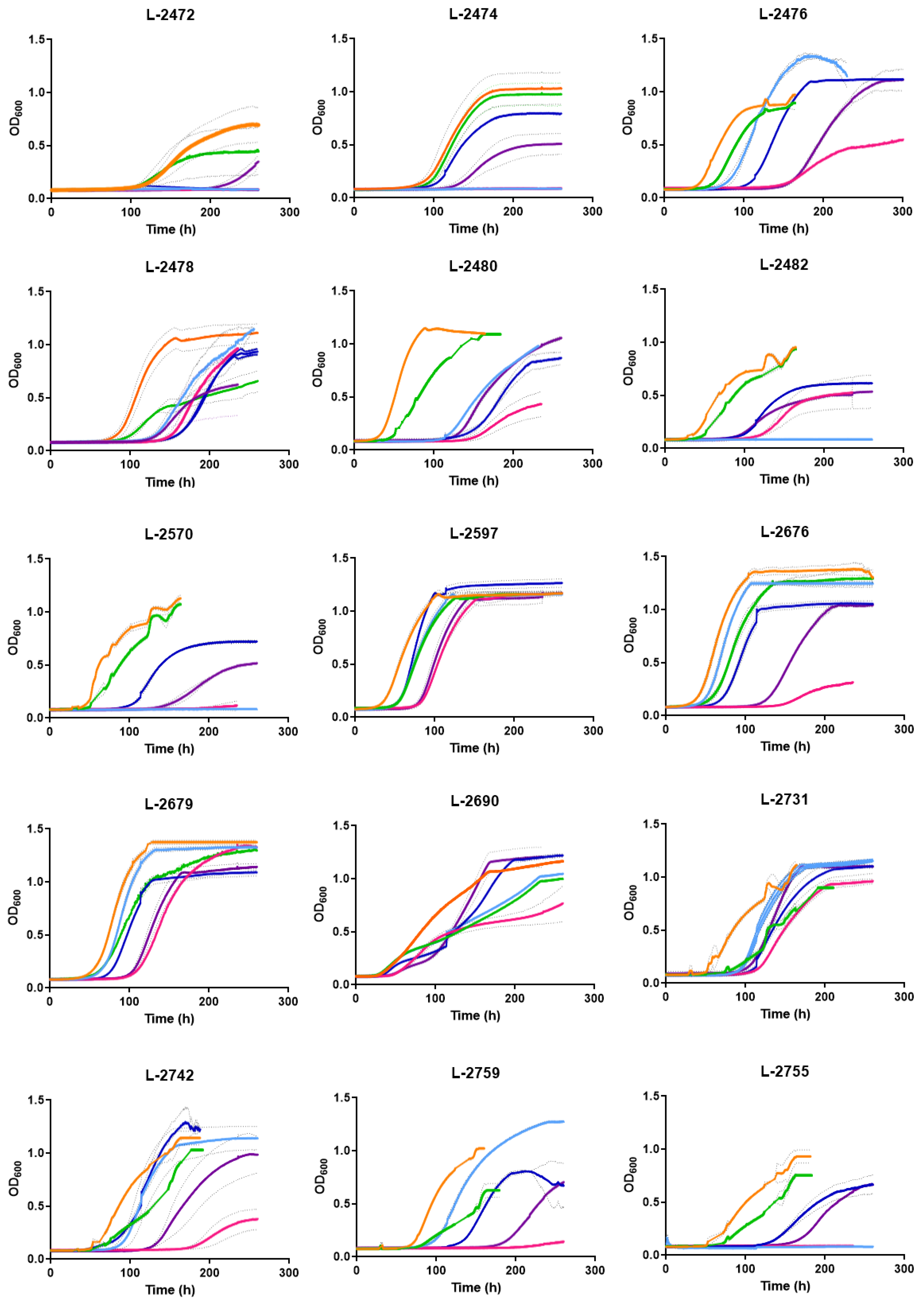
2.3. Growth Kinetics Using Micro-Cultures in the Presence of p-Coumaric Acid (APC) and Sulphur Dioxide (SO2)
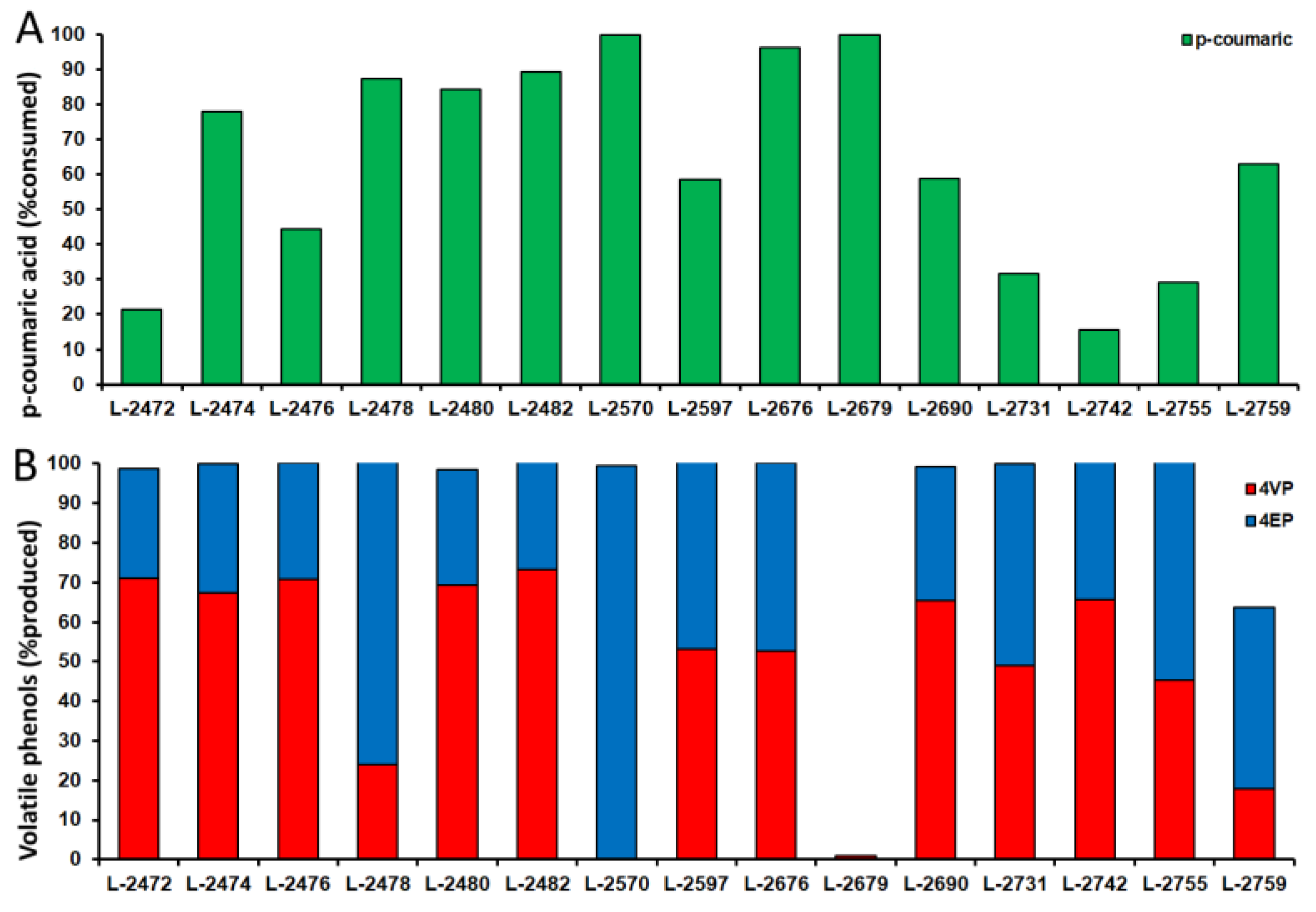
2.4. Quantification of Volatile Phenols
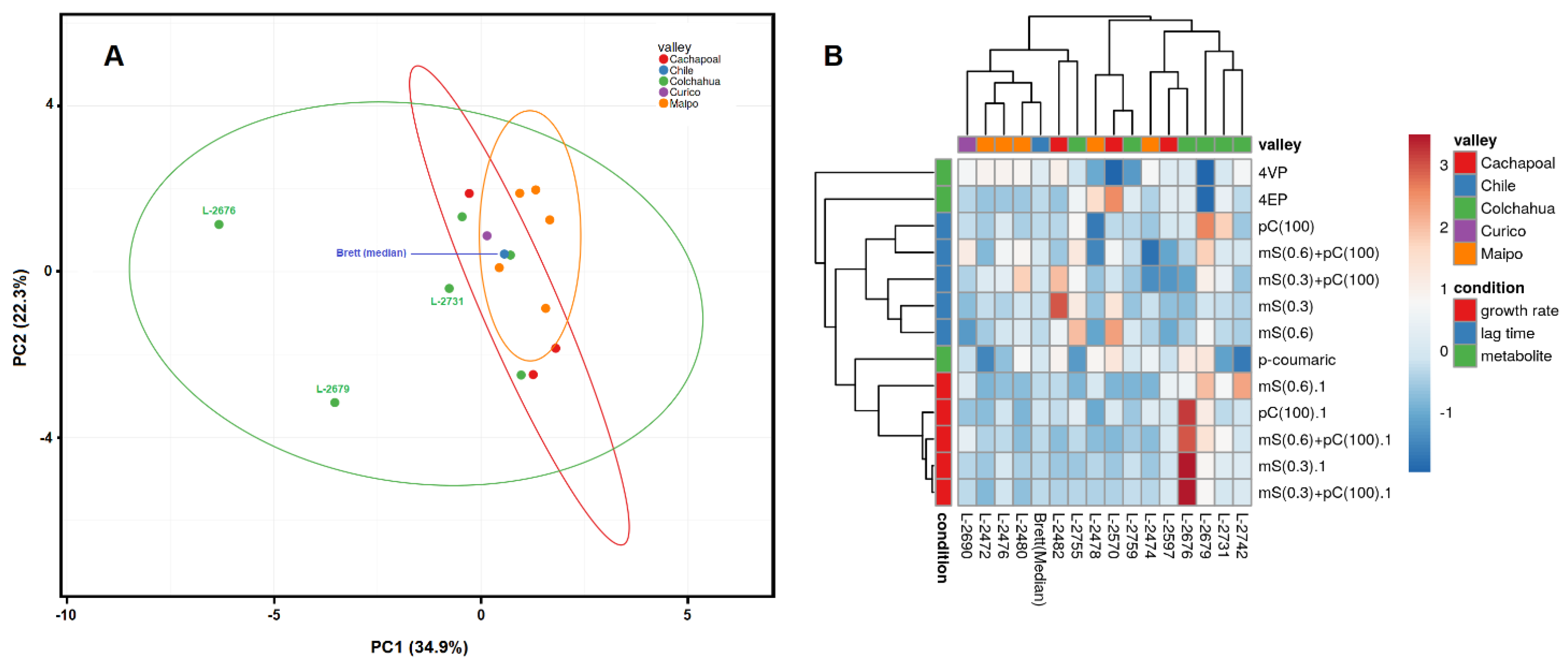
2.5. Principal Component Analysis and Clustering
3. Results
3.1. Molecular Characterization of Yeast Strains
3.2. Yeast Response to Inhibitors
3.3. Response of Yeast Strains to p-Coumaric Inhibitors Acting Simultaneously with mSO2
3.4. Metabolic Products in the Yeast Cultures
3.5. Cluster Analysis of the Brett Wine Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wedral, D.; Shewfelt, R.; Frank, J. The challenge of Brettanomyces in wine. LWT Food Sci. Technol. 2010, 43, 1474–1479. [Google Scholar] [CrossRef]
- Woolfit, M.; Rozpędowska, E.; Piškur, J.; Wolfe, K.H. Genome survey sequencing of the wine spoilage yeast Dekkera (Brettanomyces) bruxellensis. Eukaryot. Cell 2007, 6, 721–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramovič, H. Antioxidant Properties of Hydroxycinnamic Acid Derivatives: A Focus on Biochemistry, Physicochemical Parameters, Reactive Species, and Biomolecular Interactions. In Coffee in Health and Disease Prevention, 1st ed.; Preedy, V., Ed.; Academic Press: London, UK, 2015; pp. 843–852. [Google Scholar] [CrossRef]
- Campolongo, S.; Siegumfeldt, H.; Aabo, T.; Cocolin, L.; Arneborg, N. The effects of extracellular pH and hydroxycinnamic acids influence the intracellular pH of Brettanomyces bruxellensis DSM 7001. LWT Food Sci. Technol. 2014, 59, 1088–1092. [Google Scholar] [CrossRef]
- Vaquero, M.R.; Alberto, M.R.; de Nadra, M.M. Antibacterial effect of phenolic compounds from different wines. Food Control 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Laforgue, R.; Lonvaud-Funel, A. Hydroxycinnamic acid decarboxylase activity of Brettanomyces bruxellensis involved in volatile phenol production: Relationship with cell viability. Food Microbiol. 2012, 32, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; García, V.; Peña, R.; Martínez, C.; Ganga, M.A. Identification of the Dekkera bruxellensis phenolic acid decarboxylase (PAD) gene responsible for wine spoilage. Food Control. 2014, 45, 81–86. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-vinyl and 4-ethyl derivatives from hydroxycinnamic acids: Occurrence of volatile phenolic flavour compounds in beer and distribution of Pad1-activity among brewing yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Edlin, D.A.; Narbad, A.; Dickinson, J.R.; Lloyd, D. The biotransformation of simple phenolic compounds by Brettanomyces anomalus. FEMS Microbiol. Lett. 1995, 125, 311–315. [Google Scholar] [CrossRef]
- Agnolucci, M.; Rea, F.; Sbrana, C.; Cristani, C.; Fracassetti, D.; Tirelli, A.; Nuti, M. Sulphur dioxide affects culturability and volatile phenol production by Brettanomyces/Dekkera bruxellensis. Int. J. Food Microbiol. 2010, 143, 76–80. [Google Scholar] [CrossRef]
- Coulon, J.; Perello, M.C.; Lonvaud-Funel, A.; De Revel, G.; Renouf, V. Brettanomyces bruxellensis evolution and volatile phenols production in red wines during storage in bottles. J. Appl. Microbiol. 2010, 108, 1450–1458. [Google Scholar] [CrossRef]
- Serpaggi, V.; Remize, F.; Recorbet, G.; Gaudot-Dumas, E.; Sequeira-Le Grand, A.; Alexandre, H. Characterization of the “viable but nonculturable” (VBNC) state in the wine spoilage yeast Brettanomyces. Food Microbiol. 2012, 30, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. The Chemistry of Wine-Stabilization and Treatments. In Handbook of Enology, 2nd ed.; Wiley: Hoboken, NJ, USA, 2006; Volume 2, ISBN 0471973637. [Google Scholar]
- Morata, A. Red Wine Technology, 1st ed.; Academic Press: London, UK, 2018; pp. 219–235. ISBN 9780128143995. [Google Scholar]
- Pagano, D.A.; Zeiger, E.; Stark, A.A. Autoxidation and mutagenicity of sodium bisulfite. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1990, 228, 89–96. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8 S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. E Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona, L.; Varela, J.; Godoy, L.; Ganga, M.A. Comparative proteome analysis of Brettanomyces bruxellensis under hydroxycinnamic acid growth. Electron. J. Biotechnol. 2016, 23, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Avramova, M.; Vallet-Courbin, A.; Maupeu, J.; Masneuf-Pomarède, I.; Albertin, W. Molecular diagnosis of Brettanomyces bruxellensis sulfur dioxide sensitivity through genotype specific method. Front. Microbiol. 2018, 9, 1260. [Google Scholar] [CrossRef]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A comparative study on the phenolic acids identified and quantified in dry beans using HPLC as affected by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Zambrano, A.Y.; Demey, J.R.; Fuenmayor, F.; Segovia, V.; Gutiérrez, Z. Diversidad genética de una colección de yuca a través de marcadores moleculares RAPDs. Agron. Trop. 2003, 53, 155–174. [Google Scholar]
- Martorell, P.; Barata, A.; Malfeito-Ferreira, M.; Fernández-Espinar, M.T.; Loureiro, V.; Querol, A. Molecular typing of the yeast species Dekkera bruxellensis and Pichia guilliermondii recovered from wine related sources. Int. J. Food Microbiol. 2006, 106, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Agnolucci, M.; Vigentini, I.; Capurso, G.; Merico, A.; Tirelli, A.; Compagno, C.; Nuti, M. Genetic diversity and physiological traits of Brettanomyces bruxellensis strains isolated from Tuscan Sangiovese wines. Int. J. Food Microbiol. 2009, 130, 238–244. [Google Scholar] [CrossRef]
- Miot-Sertier, C.; Lonvaud-Funel, A. Development of a molecular method for the typing of Brettanomyces bruxellensis (Dekkera bruxellensis) at the strain level. J. Appl. Microbiol. 2007, 102, 555–562. [Google Scholar] [CrossRef]
- Mitrakul, C.M.; Henick-Kling, T.; Egli, C.M. Discrimination of Brettanomyces/Dekkera yeast isolates from wine by using various DNA finger-printing methods. Food Microbiol. 1999, 16, 3–14. [Google Scholar] [CrossRef]
- Martínez, C.; Cosgaya, P.; Vásquez, C.; Gac, S.; Ganga, A. High degree of correlation between molecular polymorphism and geographic origin of wine yeast strains. J. Appl. Microbiol. 2007, 103, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Garrido, D.; Martínez, C.; Saavedra, J.; Combina, M.; Ganga, M.A. Study of the coumarate decarboxylase and vinylphenol reductase activities of Dekkera bruxellensis (anamorph Brettanomyces bruxellensis) isolates. Lett. Appl. Microbiol. 2009, 48, 452–457. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Godoy, L.; Ganga, M.A. Identification of a second PAD1 in Brettanomyces bruxellensis LAMAP2480. Antonie Van Leeuwenhoek 2017, 110, 291–296. [Google Scholar] [CrossRef]
- Oelofse, A.; Lonvaud-Funel, A.; Du Toit, M. Molecular identification of Brettanomyces bruxellensis strains isolated from red wines and volatile phenol production. Food Microbiol. 2009, 26, 377–385. [Google Scholar] [CrossRef]
- Lisanti, M.T.; Blaiotta, G.; Nioi, C.; Moio, L. Alternative methods to SO2 for microbiological stabilization of wine. Compr. Rev. Food Sci. Food Saf. 2019, 18, 455–479. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.; Caldeira, J.; Botelheiro, R.; Pagliara, D.; Malfeito-Ferreira, M.; Loureiro, V. Survival patterns of Dekkera bruxellensis in wines and inhibitory effect of sulphur dioxide. Int. J. Food Microbiol. 2008, 121, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Longin, C.; Degueurce, C.; Julliat, F.; Guilloux-Benatier, M.; Rousseaux, S.; Alexandre, H. Efficiency of population-dependent sulfite against Brettanomyces bruxellensis in red wine. Food Res. Int. 2016, 89, 620–630. [Google Scholar] [CrossRef]
- Morgan, S.C.; Haggerty, J.J.; Johnston, B.; Jiranek, V.; Durall, D.M. Response to Sulfur Dioxide Addition by Two Commercial Saccharomyces cerevisiae Strains. Fermentation 2019, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Bartel, C.; Roach, M.; Borneman, A.; Curtin, C. Brettanomyces bruxellensis SSU1 haplotypes confer different levels of sulfite tolerance when expressed in a Saccharomyces cerevisiae SSU1 null mutant. Appl. Environ. Microbiol. 2019, 85, e02429-18. [Google Scholar] [CrossRef] [Green Version]
- Louw, M.; Du Toit, M.; Alexandre, H.; Divol, B. Comparative morphological characteristics of three Brettanomyces bruxellensis wine strains in the presence/absence of sulfur dioxide. Int. J. Food Microbiol. 2016, 238, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Conterno, L.; Joseph, C.L.; Arvik, T.J.; Henick-Kling, T.; Bisson, L.F. Genetic and physiological characterization of Brettanomyces bruxellensis strains isolated from wines. Am. J. Enol. Viticult. 2006, 57, 139–147. [Google Scholar]
- Curtin, C.D.; Borneman, A.R.; Chambers, P.J.; Pretorius, I.S. De-novo assembly and analysis of the heterozygous triploid genome of the wine spoilage yeast Dekkera bruxellensis AWRI1499. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Avramova, M.; Grbin, P.; Borneman, A.; Albertin, W.; Masneuf-Pomarède, I.; Varela, C. Competition experiments between Brettanomyces bruxellensis strains reveal specific adaptation to sulfur dioxide and complex interactions at intraspecies level. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, A.; Durand, C.; Loira, N.; Durrens, P.; Sherman, D.J.; Marullo, P. QTL dissection of lag phase in wine fermentation reveals a new translocation responsible for Saccharomyces cerevisiae adaptation to sulfite. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentz, M.; Harris, C. Analysis of growth inhibition and metabolism of hydroxycinnamic acids by brewing and spoilage strains of Brettanomyces yeast. Foods 2015, 4, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces yeasts—From spoilage organisms to valuable contributors to industrial fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef] [Green Version]
- Cokol, M.; Weinstein, Z.B.; Yilancioglu, K.; Tasan, M.; Doak, A.; Cansever, D.; Guvenek, A. Large-scale identification and analysis of suppressive drug interactions. Chem. Biol. 2014, 21, 541–551. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Total Number of Bands | Number of Polymorphic Bands | Polymorphism (%) | Cophenetic Correlation Coefficient |
|---|---|---|---|---|
| OPAD08 | 23 | 20 | 86.9 | 0.897 |
| OPAE09 | 21 | 17 | 80.8 | 0.945 |
| OPAE12 | 17 | 11 | 64.7 | 0.927 |
| Total bands | 61 | 48 | 77.5 |
| No Inhibitor | p-Coumaric Acid (100 mg L−1) | SO2 (0.3 mg L−1) | SO2 (0.3 mg L−1) + p-Coumaric Acid (100 mg L−1) | SO2 (0.6 mg L−1) | SO2 (0.6 mg L−1) + p-Coumaric Acid (100 mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Valley | Lag (h) | µ (h−1) | Lag (h) | µ (h−1) | Lag (h) | µ (h−1) | Lag (h) | µ (h−1) | Lag (h) | µ (h−1) | Lag (h) | µ (h−1) |
| 2472 | Maipo | 92.7 ± 2.5 | 0.005 ± 0.001 | 101.6 ± 4.6 | 0.002 ± 0.001 | 0 | 0 | 0 | 0 | 0 | 0 | 191.3 ± 2.499 | 0.003 ± 0.004 |
| 2474 | Maipo | 86.3 ± 1.6 | 0.013 ± 0.001 | 93.1 ± 3.5 | 0.014 ± 0.004 | 0 | 0 | 95.8± 5.648 | 0.010 ± 0.005 | 0 | 0 | 86.3 ± 3.22 | 0.003 ± 0.001 |
| 2476 | Maipo | 39.0 ± 1.3 | 0.016 ± 0.002 | 54.8 ± 2.6 | 0.013 ± 0.003 | 87.2 ± 3.0 | 0.022 ± 0.004 | 111.4 ± 8.562 | 0.017 ± 0.004 | 135.6 ± 4.9 | 0.002 ± 0.001 | 139.6 ± 4.441 | 0.011 ± 0.003 |
| 2478 | Maipo | 84.2 ± 0.7 | 0.018 ± 0.001 | 44.0 ± 1.7 | 0.003 ± 0.001 | 125.4 ± 5.6 | 0.009 ± 0.002 | 159.5 ± 3.885 | 0.011 ± 0.002 | 148.2 ± 3.7 | 0.011 ± 0.003 | 108.4 ± 2.554 | 0.005 ± 0.001 |
| 2680 | Maipo | 31.6 ± 2.2 | 0.035 ± 0.002 | 35.3 ± 2.4 | 0.010 ± 0.002 | 96.4 ± 4.9 | 0.010 ± 0.003 | 138.0 ± 2.445 | 0.008 ± 0.003 | 123.9 ± 4.2 | 0.003 ± 0.001 | 121.8 ± 2.645 | 0.009 ± 0.002 |
| 2482 | Cachapoal | 18.5 ± 1.7 | 0.011 ± 0.004 | 22.0 ± 3.6 | 0.008 ± 0.002 | 0 | 0 | 85.4 ± 4.533 | 0.007 ± 0.003 | 86.3 ± 2.5 | 0.004 ± 0.001 | 42.3 ± 3.441 | 0.003 ± 0.004 |
| 2570 | Cachapoal | 31.5 ± 1.4 | 0.014 ± 0.006 | 38.4 ± 3.7 | 0.014 ± 0.004 | 0 | 0 | 75.6 ± 2.114 | 0.009 ± 0.001 | 0 | 0 | 114.0 ± 2.566 | 0.003 ± 0.006 |
| 2597 | Cachapoal | 46.1 ± 2.4 | 0.021 ± 0.002 | 62.1 ± 3.1 | 0.019 ± 0.005 | 54.9 ± 4.3 | 0.022 ± 0.003 | 57.0 ± 2.159 | 0.029 ± 0.002 | 82.1 ± 4.3 | 0.019 ± 0.003 | 81.9 ± 1.823 | 0.022 ± 0.002 |
| 2676 | Colchahua | 49.7 ± 5.0 | 0.003 ± 0.002 | 67.9 ± 4.2 | 0.01 ± 0.002 | 51.8 ± 4.5 | 0.027 ± 0.004 | 71.8 ± 3.299 | 0.021 ± 0.002 | 139.4 ± 4.8 | 0.003 ± 0.002 | 125.4 ± 4.411 | 0.012 ± 0.003 |
| 2679 | Colchahua | 21.7 ± 3.2 | 0.008 ± 0.001 | 57.6 ± 2.9 | 0.014 ± 0.003 | 68.5 ± 2.4 | 0.026 ± 0.002 | 77.3 ± 4.558 | 0.021 ± 0.002 | 109.6 ± 2.6 | 0.016 ± 0.003 | 104.9 ± 2.956 | 0.019 ± 0.002 |
| 2731 | Colchahua | 35.3 ± 1.5 | 0.010 ± 0.003 | 79.0 ± 4.2 | 0.007 ± 0.001 | 94.5 ± 4.3 | 0.017 ± 0.002 | 105.9 ± 6.05 | 0.013 ± 0.002 | 112.0 ± 5.3 | 0.011 ± 0.003 | 107.3 ± 2.461 | 0.018 ± 0.004 |
| 2742 | Colchahua | 43.8 ± 2.4 | 0.011 ± 0.004 | 46.7 ± 4.2 | 0.009 ± 0.002 | 95.5 ± 2.3 | 0.018 ± 0.003 | 84.3 ± 3.24 | 0.016 ± 0.004 | 124.6 ± 4.6 | 0.024 ± 0.002 | 127.6 ± 3.224 | 0.010 ± 0.001 |
| 2755 | Colchahua | 34.0 ± 2.7 | 0.007 ± 0.001 | 59.3 ± 3.3 | 0.008 ± 0.001 | 0 | 0 | 111.9 ± 4.66 | 0.005 ± 0.002 | 0 | 0 | 145.8 ± 2.163 | 0.005 ± 0.001 |
| 2759 | Colchahua | 66.3 ± 3.6 | 0.014 ± 0.004 | 78.3 ± 2.6 | 0.007 ± 0.002 | 93.2 ± 4.3 | 0.013 ± 0.001 | 131.2 ± 3.685 | 0.013 ± 0.002 | 0 | 0 | 176.9 ± 4.998 | 0.007 ± 0.003 |
| 2690 | Curicó | 19.5 ± 2.8 | 0.009 ± 0.002 | 24.111 ± 2.4 | 0.004 ± 0.001 | 23.0 ± 2.5 | 0.005 ± 0.001 | 41.9 ± 4.627 | 0.007 ± 0.001 | 28.5 ± 1.4 | 0.006 ± 0.002 | 80.7 ± 1.437 | 0.012 ± 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
G-Poblete, C.; Peña-Moreno, I.C.; de Morais, M.A., Jr.; Moreira, S.; Ganga, M.A. Biodiversity among Brettanomyces bruxellensis Strains Isolated from Different Wine Regions of Chile: Key Factors Revealed about Its Tolerance to Sulphite. Microorganisms 2020, 8, 557. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040557
G-Poblete C, Peña-Moreno IC, de Morais MA Jr., Moreira S, Ganga MA. Biodiversity among Brettanomyces bruxellensis Strains Isolated from Different Wine Regions of Chile: Key Factors Revealed about Its Tolerance to Sulphite. Microorganisms. 2020; 8(4):557. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040557
Chicago/Turabian StyleG-Poblete, Camila, Irina Charlot Peña-Moreno, Marcos Antonio de Morais, Jr., Sandra Moreira, and María Angélica Ganga. 2020. "Biodiversity among Brettanomyces bruxellensis Strains Isolated from Different Wine Regions of Chile: Key Factors Revealed about Its Tolerance to Sulphite" Microorganisms 8, no. 4: 557. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040557