Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
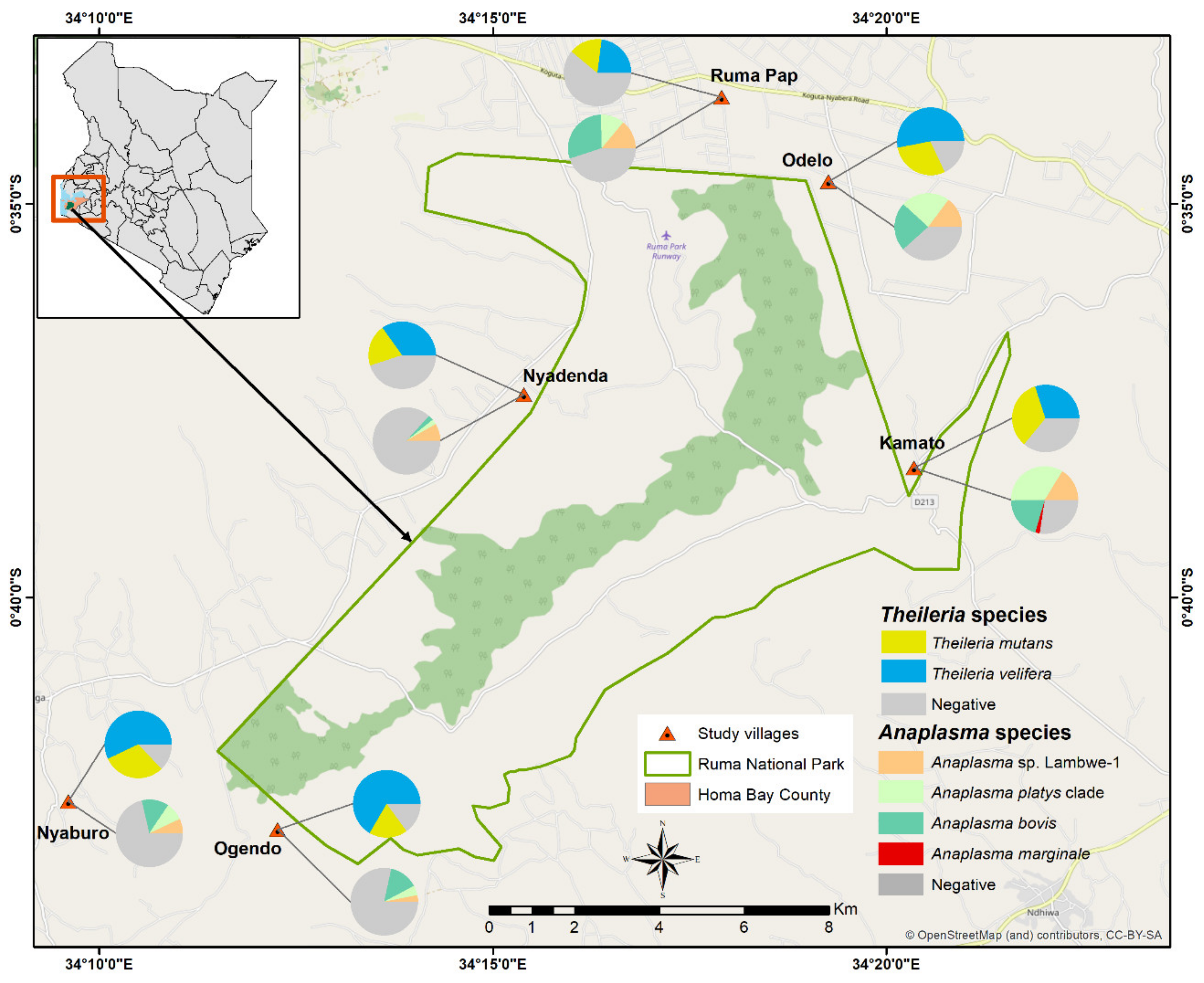
2.1. Study Area
2.2. Study Design and Sample Size Determination
2.3. Ethical Approval
2.4. Blood Sample Collection and Processing
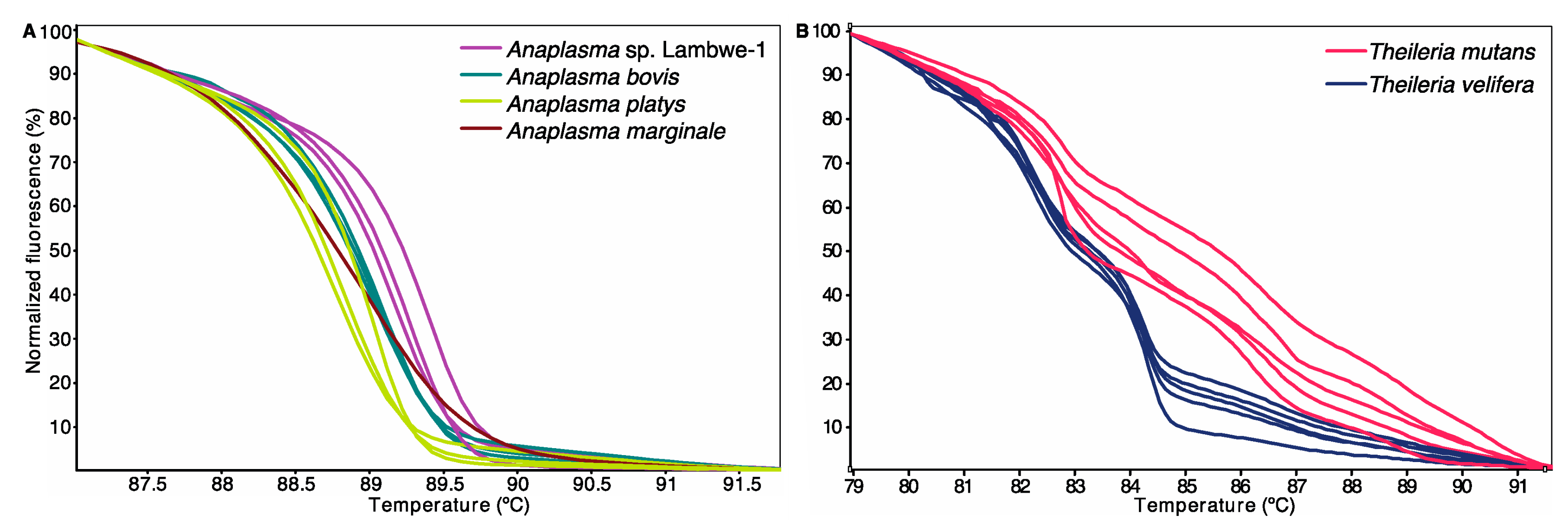
2.5. PCR and High-Resolution Melting (HRM) Analysis
2.6. Nested PCR for Anaplasma-Positive Samples
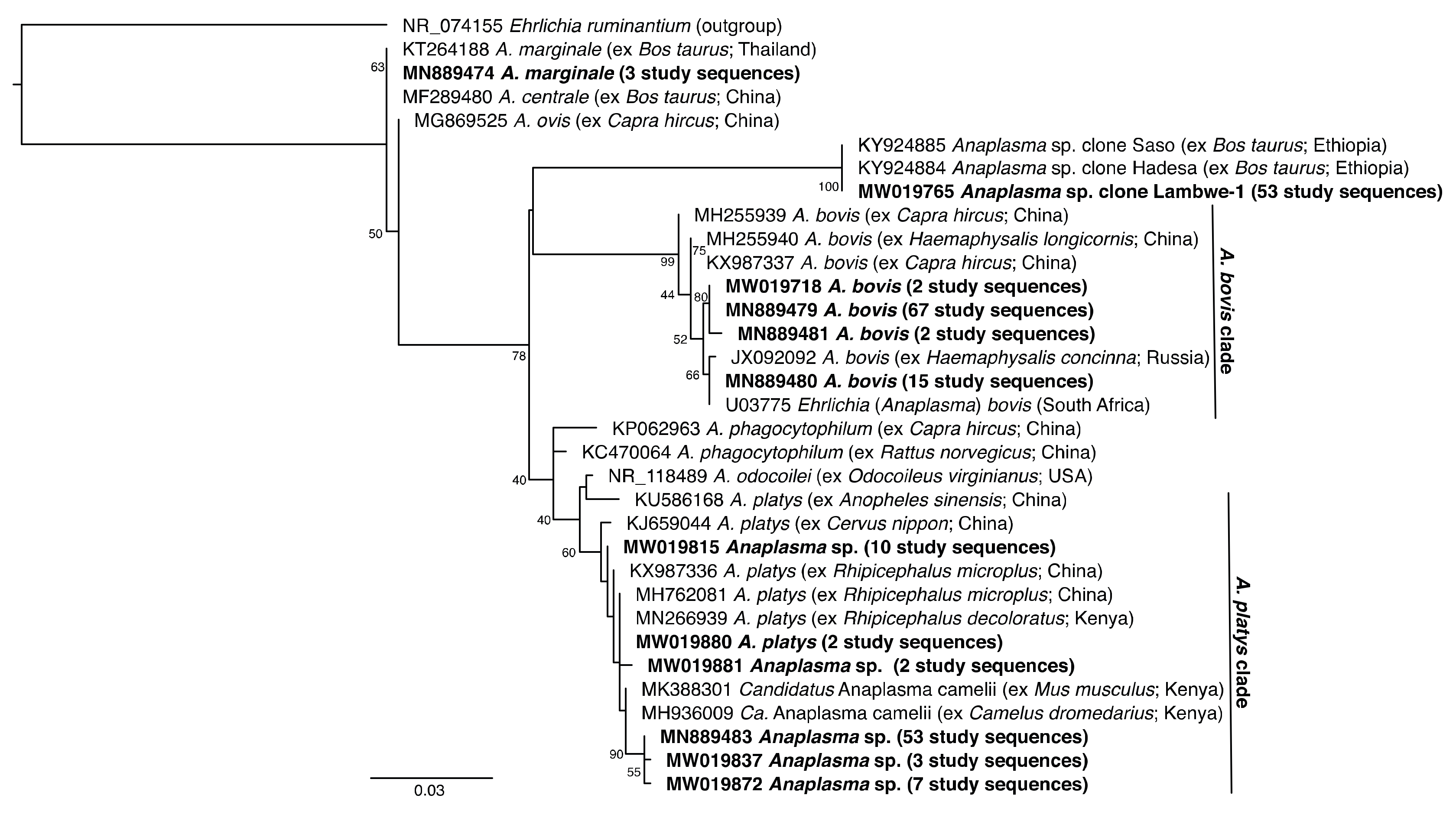
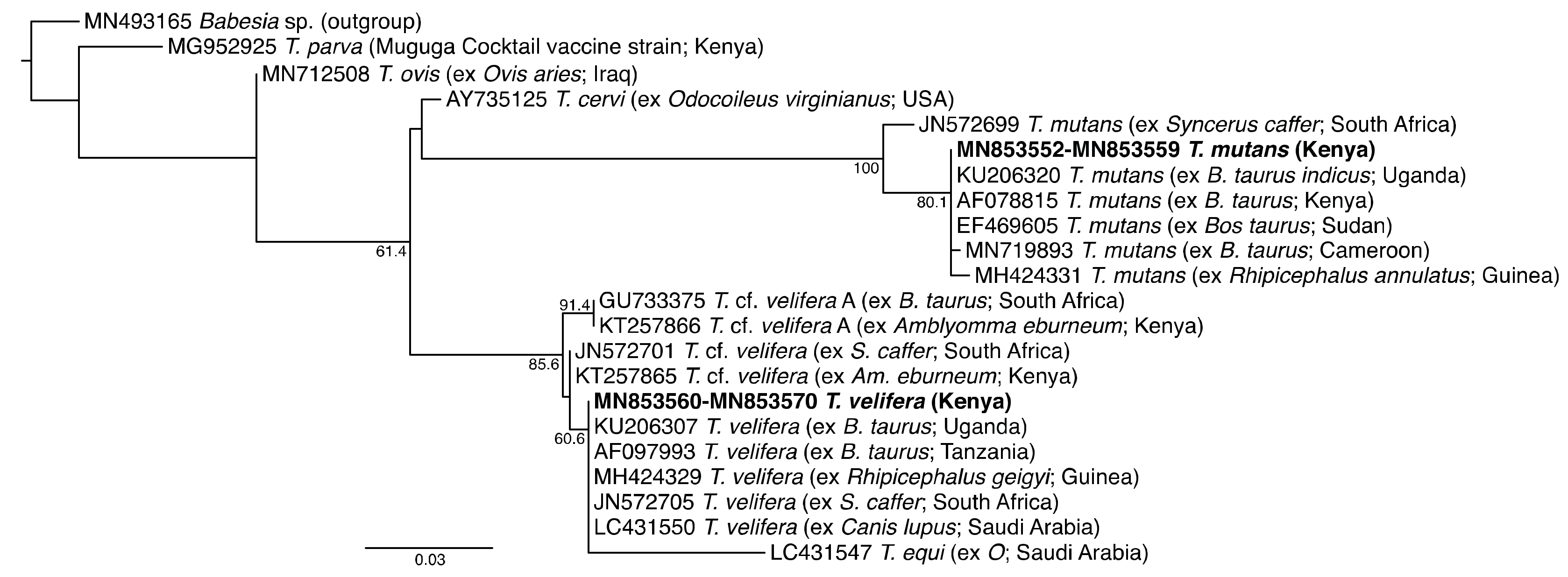
2.7. Phylogenetic Analysis
2.8. Data Management and Analysis
3. Results
3.1. Pathogen Diversity and Prevalence
3.2. Risk Factors Associated with Anaplasma and Theileria Infections
3.3. Prevalence of Co-Infections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grootenhuis, J.G.; Olubayo, R.O. Disease research in the wildlife-livestock interface in Kenya. Vet. Q. 1993, 15, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Miguel, E.; Gomo, C.; Makaya, P.; Pfukenyi, D.M.; Foggin, C.; Hove, T.; De Garine-Wichatitsky, M. Relationship between burden of infection in ungulate populations and wildlife/livestock interfaces. Epidemiol. Infect. 2013, 141, 1522–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.S.; Farnsworth, M.L.; Malmberg, J.L. Diseases at the livestock–wildlife interface: Status, challenges, and opportunities in the United States. Prev. Vet. Med. 2013, 110, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwamuye, M.M.; Kariuki, E.; Omondi, D.; Kabii, J.; Odongo, D.; Masiga, D.; Villinger, J. Novel Rickettsia and emergent tick-borne pathogens: A molecular survey of ticks and tick-borne pathogens in Shimba Hills National Reserve, Kenya. Ticks Tick Borne Dis. 2017, 8, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife- threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Minjauw, B.; McLeod, A. Tick-Borne Diseases and Poverty. The Impact of Ticks and Tick-Borne Diseases on the Livelihood of Small-Scale and Marginal Livestock Owners in India and Eastern and Southern Africa; Research Report, DFID Animal Health Programme; University of Edinburgh: Edinburgh, UK, 2003. [Google Scholar]
- Ngeranwa, J.J.; Shompole, S.P.; Venter, E.H.; Wambugu, A.; Crafford, J.E.; Penzhorn, B.L. Detection of Anaplasma antibodies in wildlife and domestic species in wildlife-livestock interface areas of Kenya by major surface protein 5 competitive inhibition enzyme-linked immunosorbent assay. Onderstepoort J. Vet. Res. 2008, 75, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Ndeereh, D.; Muchemi, G.; Thaiyah, A.; Otiende, M.; Angelone-Alasaad, S.; Jowers, M.J. Molecular survey of Coxiella burnetii in wildlife and ticks at wildlife-livestock interfaces in Kenya. Exp. Appl. Acarol. 2017, 72, 277–289. [Google Scholar] [CrossRef]
- Omondi, D.; Masiga, D.K.; Ajamma, Y.U.; Fielding, B.C.; Njoroge, L.; Villinger, J. Unraveling host-vector-arbovirus interactions by two-gene high resolution melting mosquito bloodmeal analysis in a Kenyan wildlife-livestock interface. PLoS ONE 2015, 10, e0134375. [Google Scholar] [CrossRef]
- Oundo, J.W.; Villinger, J.; Jeneby, M.; Ong’amo, G.; Otiende, M.Y.; Makhulu, E.E.; Musa, A.A.; Ouso, D.O.; Wambua, L. Pathogens, endosymbionts, and blood-meal sources of host-seeking ticks in the fast-changing Maasai Mara wildlife ecosystem. PLoS ONE 2020, 15, e0228366. [Google Scholar] [CrossRef]
- Njiiri, N.E.; Bronsvoort, B.M.d.; Collins, N.E.; Steyn, H.C.; Troskie, M.; Vorster, I.; Thumbi, S.M.; Sibeko, K.P.; Jennings, A.; van Wyk, I.C.; et al. The epidemiology of tick-borne haemoparasites as determined by the reverse line blot hybridization assay in an intensively studied cohort of calves in western Kenya. Vet. Parasitol. 2015, 210, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Kock, R.A. What is this infamous “wildlife/livestock disease interface?” A review of current knowledge for the African continent. In Conservation and Development Interventions at the Wildlife/Livestock Interface: Implications for Wildlife, Livestock and Human Health; Osofsky, S.A., Cleaveland, S., Karesh, W.B., Kock, M.D., Nyhus, P.J., Star, L., Yang, A., Eds.; International Union for Conservation of Nature: Cambridge, UK, 2005; pp. 1–13. [Google Scholar]
- Morrison, W.I.; Hemmink, J.D.; Toye, P.G. Theileria parva: A parasite of African buffalo, which has adapted to infect and undergo transmission in cattle. Int. J. Parasitol. 2020, 50, 403–412. [Google Scholar] [CrossRef]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector-Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Kenya Wildlife Conservancies Association. State of Wildlife Conservancies in Kenya Report. 2016. Available online: https://kwcakenya.com/download/state-of-wildlife-conservancies-in-kenya-report/ (accessed on 28 October 2020).
- Muriuki, G.W.; Njoka, T.J.; Reid, R.S.; Nyariki, D.M. Tsetse control and land-use change in Lambwe Valley, south-western Kenya. Agric. Ecosyst. Environ. 2005, 106, 99–107. [Google Scholar] [CrossRef]
- Wellde, B.T.; Chumo, D.A.; Reardon, M.J.; Waema, D.; Smith, D.H.; Gibson, W.C.; Wanyama, L.; Siongok, T.A. Epidemiology of Rhodesian sleeping sickness in the Lambwe Valley, Kenya. Ann. Trop. Med. Parasitol. 1989, 83, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Ogutu, J.O.; Piepho, H.P.; Said, M.Y.; Ojwang, G.O.; Njino, L.W.; Kifugo, S.C.; Wargute, P.W. Extreme wildlife declines and concurrent increase in livestock numbers in Kenya: What are the causes? PLoS ONE 2016, 11, e0163249. [Google Scholar] [CrossRef]
- Morse, S.S. Factors in the Emergence of Infectious Diseases; Price-Smith, A.T., Ed.; Palgrave Macmillan UK: London, UK, 2001; pp. 8–26. [Google Scholar] [CrossRef] [Green Version]
- Otieno, D.O.; K’Otuto, G.O.; Jákli, B.; Schröttle, P.; Maina, J.N.; Jung, E.; Onyango, J.C. Spatial heterogeneity in ecosystem structure and productivity in a moist Kenyan savanna. Plant. Ecol. 2011, 212, 769–783. [Google Scholar] [CrossRef]
- Bennett, S.; Woods, T.; Liyanage, W.M.; Smith, D.L. A simplified general method for cluster-sample surveys of health in developing countries. World Health Stat. Q. 1991, 44, 98–106. [Google Scholar]
- Dohoo, I.R.; Martin, S.W.; Stryhn, H. Veterinary Epidemiologic Research; VER, Inc.: Charlottetown, PE, Canada, 2009; pp. 33–55. [Google Scholar]
- Thrusfield, M.V.; Christley, R. Veterinary Epidemiology, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2018; p. 864. [Google Scholar]
- Otte, M.J.; Gumm, I.D. Intra-cluster correlation coefficients of 20 infections calculated from the results of cluster-sample surveys. Prev. Vet. Med. 1997, 31, 147–150. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Bodaan, C.; Postigo, M.; Nieuwenhuijs, H.; Opsteegh, M.; Franssen, L.; Jebbink, F.; Jongejan, F. Ticks and associated pathogens collected from domestic animals in the Netherlands. Vector Borne Zoonotic Dis. 2007, 7, 585–595. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Mohammed, O.B.; Bennett, N.C.; Petevinos, C.; Alagaili, A.N. Molecular detection of novel Anaplasmataceae closely related to Anaplasma platys and Ehrlichia canis in the dromedary camel (Camelus dromedarius). Vet. Microbiol. 2015, 179, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Georges, K.; Loria, G.R.; Riili, S.; Greco, A.; Caracappa, S.; Jongejan, F.; Sparagano, O. Detection of haemoparasites in cattle by reverse line blot hybridisation with a note on the distribution of ticks in Sicily. Vet. Parasitol. 2001, 99, 273–286. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree; Version 1.4. 2; University of Edinburgh: Edinburgh, UK, 2014. [Google Scholar]
- Ringo, A.E.; Rizk, M.A.; Moumouni, P.F.A.; Liu, M.; Galon, E.M.; Li, Y.; Ji, S.; Tumwebaze, M.; Byamukama, B.; Thekisoe, O. Molecular detection and characterization of tick-borne haemoparasites among cattle on Zanzibar Island, Tanzania. Acta Trop. 2020, 211, 105598. [Google Scholar] [CrossRef]
- Tayebwa, D.S.; Vudriko, P.; Tuvshintulga, B.; Guswanto, A.; Nugraha, A.B.; Gantuya, S.; Batiha, G.E.-S.; Musinguzi, S.P.; Komugisha, M.; Bbira, J.S. Molecular epidemiology of Babesia species, Theileria parva, and Anaplasma marginale infecting cattle and the tick control malpractices in Central and Eastern Uganda. Ticks Tick Borne Dis. 2018, 9, 1475–1483. [Google Scholar] [CrossRef]
- Hailemariam, Z.; Krucken, J.; Baumann, M.; Ahmed, J.S.; Clausen, P.H.; Nijhof, A.M. Molecular detection of tick-borne pathogens in cattle from Southwestern Ethiopia. PLoS ONE 2017, 12, e0188248. [Google Scholar] [CrossRef] [Green Version]
- Battilani, M.; De Arcangeli, S.; Balboni, A.; Dondi, F. Genetic diversity and molecular epidemiology of Anaplasma. Infect. Genet. Evol. 2017, 49, 195–211. [Google Scholar] [CrossRef]
- Omondi, D.; Masiga, D.K.; Fielding, B.C.; Kariuki, E.; Ajamma, Y.U.; Mwamuye, M.M.; Ouso, D.O.; Villinger, J. Molecular detection of tick-borne pathogen diversities in ticks from livestock and reptiles along the shores and adjacent islands of Lake Victoria and Lake Baringo, Kenya. Front. Vet. Sci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Arraga-Alvarado, C.M.; Qurollo, B.A.; Parra, O.C.; Berrueta, M.A.; Hegarty, B.C.; Breitschwerdt, E.B. Case report: Molecular evidence of Anaplasma platys infection in two women from venezuela. Am. J. Trop. Med. Hyg. 2014, 91, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Matei, I.A.; D’Amico, G.; Yao, P.K.; Ionică, A.M.; Kanyari, P.W.N.; Daskalaki, A.A.; Dumitrache, M.O.; Sándor, A.D.; Gherman, C.M.; Qablan, M. Molecular detection of Anaplasma platys infection in free-roaming dogs and ticks from Kenya and Ivory Coast. Parasit Vectors. 2016, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Li, K.; Sun, Y.; Shi, J.; Li, H.; Chen, Y.; Yang, H.; Li, X.; Wu, B.; Li, X. Molecular epidemiology and risk factors of Anaplasma spp., Babesia spp. and Theileria spp. infection in cattle in Chongqing, China. PLoS ONE 2019, 14, e0215585. [Google Scholar] [CrossRef] [Green Version]
- Said, M.B.; Belkahia, H.; El Mabrouk, N.; Saidani, M.; Alberti, A.; Zobba, R.; Cherif, A.; Mahjoub, T.; Bouattour, A.; Messadi, L. Anaplasma platys-like strains in ruminants from Tunisia. Infect. Genet. Evol. 2017, 49, 226–233. [Google Scholar] [CrossRef]
- André, M.R.; Calchi, A.C.; Herrera, H.M.; de Souza Zanatto, D.C.; Horta, B.d.C.L.S.; Tasso, J.B.; de Souza Ramos, I.A.; de Mello, V.V.C.; Machado, R.Z. The co-infection with Ehrlichia minasensis, Anaplasma marginale and Anaplasma platys is not associated with anemia in beef cattle in the Brazilian Pantanal. Vet. Parasitol. Reg. Stud. Reports 2020, 21, 100437. [Google Scholar] [CrossRef]
- Peter, S.G.; Aboge, G.O.; Kariuki, H.W.; Kanduma, E.G.; Gakuya, D.W.; Maingi, N.; Mulei, C.M.; Mainga, A.O. Molecular prevalence of emerging Anaplasma and Ehrlichia pathogens in apparently healthy dairy cattle in peri-urban Nairobi, Kenya. BMC Vet. Res. 2020, 16, 364. [Google Scholar] [CrossRef]
- Adjou Moumouni, P.F.; Aboge, G.O.; Terkawi, M.A.; Masatani, T.; Cao, S.; Kamyingkird, K.; Jirapattharasate, C.; Zhou, M.; Wang, G.; Liu, M.; et al. Molecular detection and characterization of Babesia bovis, Babesia bigemina, Theileria species and Anaplasma marginale isolated from cattle in Kenya. Parasit. Vectors 2015, 8, 496. [Google Scholar] [CrossRef] [Green Version]
- Byaruhanga, C.; Collins, N.E.; Knobel, D.; Chaisi, M.E.; Vorster, I.; Steyn, H.C.; Oosthuizen, M.C. Molecular investigation of tick-borne haemoparasite infections among transhumant zebu cattle in Karamoja Region, Uganda. Vet. Parasitol. Reg. Stud. Reports 2016, 3-4, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Chiuya, T.; Masiga, D.; Falzon, L.; Bastos, A.; Fevre, E.; Villinger, J. Tick-borne pathogens, including Crimean-Congo haemorrhagic fever virus, at livestock markets and slaughterhouses in western Kenya. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Simuunza, M.; Weir, W.; Courcier, E.; Tait, A.; Shiels, B. Epidemiological analysis of tick-borne diseases in Zambia. Vet. Parasitol. 2011, 175, 331–342. [Google Scholar] [CrossRef]
- Moll, G.; Lohding, A.; Young, A.S.; Leitch, B.L. Epidemiology of theileriosis in calves in an endemic area of Kenya. Vet. Parasitol. 1986, 19, 255–273. [Google Scholar] [CrossRef]
- Ringo, A.E.; Aboge, G.O.; Adjou Moumouni, P.F.; Lee, S.H.; Jirapattharasate, C.; Liu, M.; Gao, Y.; Guo, H.; Zheng, W.; Efstratiou, A.; et al. Molecular detection and genetic characterisation of pathogenic Theileria, Anaplasma and Ehrlichia species among apparently healthy sheep in central and western Kenya. Onderstepoort J. Vet. Res. 2019, 86, 1630. [Google Scholar] [CrossRef] [PubMed]
- Telfer, S.; Lambin, X.; Birtles, R.; Beldomenico, P.; Burthe, S.; Paterson, S.; Begon, M. Species interactions in a parasite community drive infection risk in a wildlife population. Science 2010, 330, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.E.J.; Thumbi, S.M.; Jennings, A.; Chase-Topping, M.; Callaby, R.; Kiara, H.; Oosthuizen, M.C.; Mbole-Kariuki, M.N.; Conradie, I.; Handel, I.G.; et al. Co-infections determine patterns of mortality in a population exposed to parasite infection. Sci. Adv. 2015, 1, e1400026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, A.C.; Armstrong, S.D.; Bah, G.S.; Kaur, G.; Hughes, M.A.; Kay, S.M.; Koldkjær, P.; Rainbow, L.; Radford, A.D.; Blaxter, M.L. Analysis of gene expression from the Wolbachia genome of a filarial nematode supports both metabolic and defensive roles within the symbiosis. Genome Res. 2012, 22, 2467–2477. [Google Scholar] [CrossRef] [Green Version]
- Ola-Fadunsin, S.D.; Gimba, F.I.; Abdullah, D.A.; Sharma, R.S.K.; Abdullah, F.J.F.; Sani, R.A. Epidemiology and risk factors associated with Anaplasma marginale infection of cattle in Peninsular Malaysia. Parasitol. Int. 2018, 67, 659–665. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Pathogens | Target Gene | Primer Name | Sequence (5′ to 3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| Rickettsia spp. | 16S rRNA | Rick-F | GAACGCTATCGGTATGCTTAACACA | 364 | [26] |
| Rick-R | CATCACTCACTCGGTATTGCTGGA | ||||
| Ehrlichia spp. | 16S rRNA | EhrlichiaJV F | GCAACCCTCATCCTTAGTTACCA | 300 | [5] |
| EhrlichiaJV R | TGTTACGACTTCACCCTAGTCAC | ||||
| Anaplasma spp. | 16S rRNA | AnaplasmaJV F | CGGTGGAGCATGTGGTTTAATTC | 300 | [5] |
| AnaplasmaJV R | CGRCGTTGCAACCTATTGTAGTC | ||||
| Anaplasmataceae | 16S rRNA | EHR16SD | GGTACCYACAGAAGAAGTCC | [27] | |
| pH1522 | AAGGAGGTGATCCAGCCGCA | 1060 | |||
| pH1492 | GGCTACCTTGTTACGACTT | 1030 | |||
| Theileria and Babesia spp. | 18S rRNA | RLB F | GAGGTAGTGACAAGAAATAACAATA | 450 | [28] |
| RLB R | TCTTCGATCCCCTAACTTTC |
| Pathogen | Individual Prevalence | Herd Prevalence | ||||
|---|---|---|---|---|---|---|
| na | Prevalence (%) | 95% CI | nb | Prevalence (%) | 95% CI | |
| Anaplasma spp. | 311 | 45.7 | 42.0, 49.5 | 75 | 78.9 | 70.8, 87.2 |
| A. bovis | 118 | 17.4 | 14.5, 20.2 | 55 | 57.9 | 48.0, 67.8 |
| A. platys clade | 115 | 16.9 | 14.1, 19.7 | 49 | 51.6 | 41.5, 61.6 |
| A. marginale | 4 | 0.6 | 0.0, 1.2 | 4 | 4.2 | 0.2, 8.3 |
| Anaplasma sp. Lambwe-1 | 79 | 11.6 | 9.2, 14.0 | 39 | 41.1 | 31.2, 50.9 |
| Theileria spp. | 432 | 63.5 | 59.9, 67.2 | 84 | 88.4 | 82.0, 94.9 |
| T. velifera | 272 | 40.0 | 36.3, 43.7 | 72 | 75.8 | 67.2, 84.4 |
| T. mutans | 175 | 25.7 | 22.5, 29.0 | 62 | 65.3 | 55.7, 74.8 |
| Overall TBPs | 680 | 78.5 | 75.3, 81.5 | 91 | 95.8 | 91.8, 99.8 |
| Pathogen Detected | Number Positive (% Prevalence) | 95% CI |
|---|---|---|
| Single infections | 319 (46.9) | 43.7, 50.7 |
| A. marginale | 2 (0.3) | 0.0, 0.7 |
| A. bovis | 36 (5.3) | 3.6, 6.9 |
| A. platys clade | 35 (5.2) | 3.5, 6.8 |
| Anaplasma sp. Lambwe-1 | 29 (4.3) | 2.8, 5.8 |
| T. velifera | 145 (21.0) | 18.2, 24.4 |
| T. mutans | 72 (11.0) | 8.3, 12.9 |
| Double infections | 200 (29.4) | 26.0, 32.8 |
| A. bovis + T. mutans | 31 (4.6) | 3.0, 6.1 |
| A. bovis + T. velifera | 45 (6.6) | 4.8, 8.5 |
| A. marginale + T. velifera | 2 (0.3) | 0.0, 0.7 |
| A. platys clade + T. mutans | 36 (5.3) | 3.5, 6.8 |
| A. platys clade + T. velifera | 35 (5.2) | 3.5, 6.8 |
| Anaplasma sp. Lambwe-1 + T. velifera | 29 (4.3) | 2.8, 5.8 |
| Anaplasma sp. Lambwe-1 + T. mutans | 17 (2.5) | 1.3, 3.7 |
| T. velifera + T. mutans | 6 (1.0) | 0.2, 1.6 |
| Triple infections | 15 (2.2) | 1.1, 3.3 |
| A. bovis + A. platys clade + T. mutans | 1 (0.2) | 0.0, 0.4 |
| Anaplasma sp. Lambwe-1 + A. platys clade + T. mutans | 3 (0.4) | 0.0, 0.9 |
| Anaplasma sp. Lambwe-1 + A. platys clade + T. velifera | 1 (0.2) | 0.0, 0.4 |
| A. bovis + T. velifera + T. mutans | 5(0.7) | 0.0, 1.4 |
| A. platys clade + T. mutans + T. velifera | 4 (0.6) | 0.0, 1.2 |
| Total | 534 (78.5) | 75.4, 81.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okal, M.N.; Odhiambo, B.K.; Otieno, P.; Bargul, J.L.; Masiga, D.; Villinger, J.; Kalayou, S. Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface. Microorganisms 2020, 8, 1830. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111830
Okal MN, Odhiambo BK, Otieno P, Bargul JL, Masiga D, Villinger J, Kalayou S. Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface. Microorganisms. 2020; 8(11):1830. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111830
Chicago/Turabian StyleOkal, Michael N., Brenda Kisia Odhiambo, Peter Otieno, Joel L. Bargul, Daniel Masiga, Jandouwe Villinger, and Shewit Kalayou. 2020. "Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface" Microorganisms 8, no. 11: 1830. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111830