Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy

 , ,
, ,  , , , , and
, , , , and 
Abstract
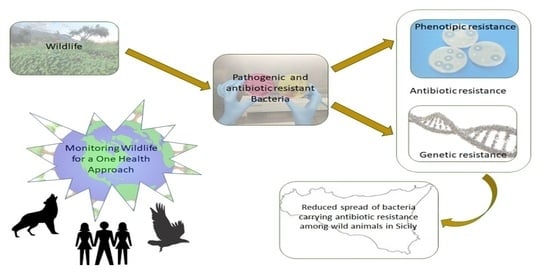
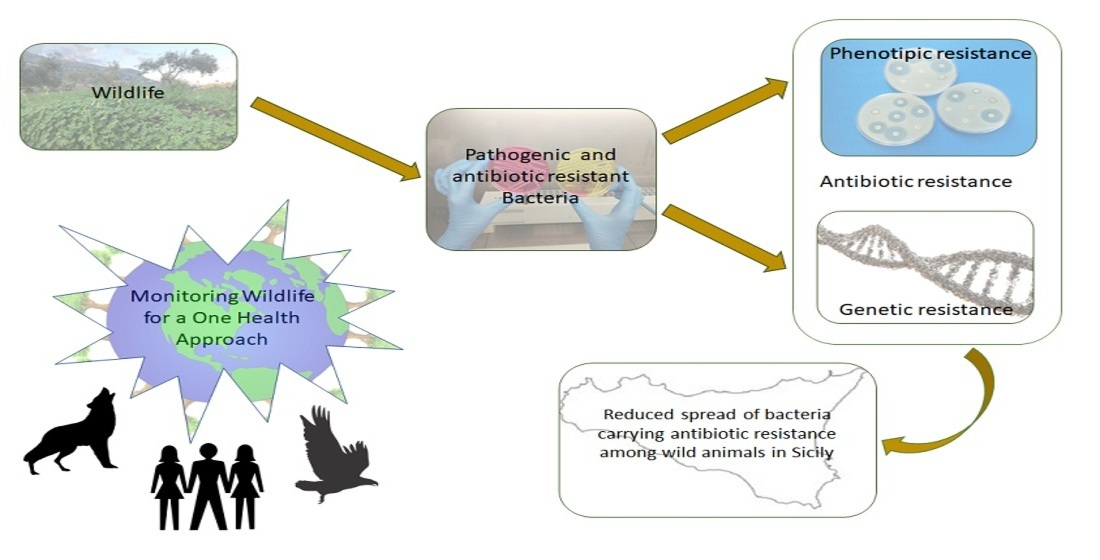
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Bacterial Isolation and Identification
2.3. Antimicrobial Susceptibility Using the Disk Diffusion Method
2.4. Detection of Antibiotic Resistance Genes and Class-1 Integron
3. Results
3.1. Host Animal Species and Samples Collected
3.2. Bacterial Detection
3.3. Antimicrobial Susceptibility Using the Disk Diffusion Method
3.4. Detention of Resistance Genes and Int1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Travis, D.A.; Watson, R.P.; Tauer, A. The Spread of Pathogens through Trade in Wildlife: -EN- -FR- La Dissémination d’agents Pathogènes Lors Des Échanges Internationaux d’animaux Sauvages-ES-Propagación de Agentes Patógenos Por El Comercio de Animales Salvajes. Rev. Sci. Tech. OIE 2011, 30, 219–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfranchi, P.; Ferroglio, E.; Poglayen, G.; Guberti, V. Wildlife Veterinarian, Conservation and Public Health. Vet. Res. Commun. 2003, 27, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Friend, M. Why Bother about Wildlife Disease? Circular 1401; U.S. Geological Survey: Reston, VA, USA, 2014; Volume 1401, p. 90.
- Abrantes, J.; Van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit Haemorrhagic Disease (RHD) and Rabbit Haemorrhagic Disease Virus (RHDV): A Review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE (World Organization for Animal Health). Report of the Meeting of the OIE Working Group on Wildlife Diseases; OIE: Paris, France, 2004. [Google Scholar]
- International Office of Epizootics; Biological Standards Commission. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: (Mammals, Birds and Bees); OIE: Paris, France, 2018; ISBN 978-92-95108-18-9. [Google Scholar]
- Yon, L.; Duff, J.P.; Ågren, E.O.; Erdélyi, K.; Ferroglio, E.; Godfroid, J.; Hars, J.; Hestvik, G.; Horton, D.; Kuiken, T.; et al. Recent Changes in Infectious Diseases in European Wildlife. J. Wildl. Dis. 2019, 55, 3. [Google Scholar] [CrossRef] [PubMed]
- Karesh, W.B.; Cook, R.A.; Bennett, E.L.; Newcomb, J. Wildlife Trade and Global Disease Emergence. Emerg. Infect. Dis. 2005, 11, 1000–1002. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.E.; Williams, N.J.; Bennett, M. ‘Disperse Abroad in the Land’: The Role of Wildlife in the Dissemination of Antimicrobial Resistance. Biol. Lett. 2016, 12, 20160137. [Google Scholar] [CrossRef] [Green Version]
- Gwenzi, W.; Musiyiwa, K.; Mangori, L. Sources, Behaviour and Health Risks of Antimicrobial Resistance Genes in Wastewaters: A Hotspot Reservoir. J. Environ. Chem. Eng. 2020, 8, 102220. [Google Scholar] [CrossRef]
- Peterson, E.; Kaur, P. Antibiotic Resistance Mechanisms in Bacteria: Relationships Between Resistance Determinants of Antibiotic Producers, Environmental Bacteria, and Clinical Pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef]
- Karkman, A.; Pärnänen, K.; Larsson, D.G.J. Fecal Pollution Can Explain Antibiotic Resistance Gene Abundances in Anthropogenically Impacted Environments. Nat. Commun. 2019, 10, 80. [Google Scholar] [CrossRef]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Mazel, D. Integrons: Agents of Bacterial Evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Furness, L.E.; Campbell, A.; Zhang, L.; Gaze, W.H.; McDonald, R.A. Wild Small Mammals as Sentinels for the Environmental Transmission of Antimicrobial Resistance. Environ. Res. 2017, 154, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Huang, Y.; Rao, D.; Zhang, Y.; Yang, K. Evidence for Environmental Dissemination of Antibiotic Resistance Mediated by Wild Birds. Front. Microbiol. 2018, 9, 745. [Google Scholar] [CrossRef] [PubMed]
- Alduina, R.; Gambino, D.; Presentato, A.; Gentile, A.; Sucato, A.; Savoca, D.; Filippello, S.; Visconti, G.; Caracappa, G.; Vicari, D.; et al. Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea? Antibiotics 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, K.H.; Khor, W.C.; Quek, J.Y.; Low, Z.X.; Arivalan, S.; Humaidi, M.; Chua, C.; Seow, K.L.G.; Guo, S.; Tay, M.Y.F.; et al. Occurrence and Antimicrobial Resistance Traits of Escherichia Coli from Wild Birds and Rodents in Singapore. IJERPH 2020, 17, 5606. [Google Scholar] [CrossRef]
- Lupindu, A.M. Isolation and Characterization of Escherichia coli from Animals, Humans, and Environment. In Escherichia coli—Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; Samie, A., Ed.; InTech: West Palm Beach, FL, USA, 2017; ISBN 978-953-51-3329-2. [Google Scholar]
- Boone, D.R.; Castenholz, R.W.; Garrity, G.M. (Eds.) Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2001; ISBN 978-0-387-98771-2. [Google Scholar]
- Rahmani, S.; Forozandeh, M.; Mosavi, M.; Rezaee, A. Detection of Bacteria by Amplifying the 16S RRNA Gene with Universal Primers and RFLP. Med. J. Islam. Rep. Iran 2006, 19, 333–338. [Google Scholar]
- CLSI Performance Standards for Antimicrobial Disk and Dilution Susceptibility. Tests for Bacteria Isolated from Animals CLSI Supplement VET08, 4th ed.; Committee for Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; ISBN 978-1-68440-010-2. [Google Scholar]
- EUCAST The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0 2020. Available online: http://www.eucast.org (accessed on 27 April 2020).
- EUCAST Intrinsic Resistance and Unusual Phenotypes, Version 3.2 2020. February 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Expert_Rules/2020/Intrinsic_Resistance_and_Unusual_Phenotypes_Tables_v3.2_20200225.pdf (accessed on 27 April 2020).
- Giacopello, C.; Foti, M.; Mascetti, A. Antimicrobial Resistance Patterns of Enterobacteriaceae in European Wild Bird Species Admitted in a Wildlife Rescue Centre. Vet. Ital. 2016, 139–144. [Google Scholar] [CrossRef]
- Silva, R.O.S.; Almeida, L.R.; Junior, C.A.O.; Lima, P.C.S.; Soares, D.F.M.; Pereira, P.L.L.; Silva, I.J.; Lobato, F.C.F. Isolation and Genotyping of Clostridium Perfringens from Free-Living South American Coati (Nasua Nasua). J. Zoo Wildl. Med. 2016, 47, 333–336. [Google Scholar] [CrossRef]
- Rahman, M.S.; Sharma, R.; Borah, P.; Chakraborty, A.; Devi, M.R.; Longjam, N. Characterization of Clostridium Perfringens Isolated from Mammals and Birds from Guwahati City, India. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Bertelloni, F.; Cilia, G.; Bogi, S.; Ebani, V.V.; Turini, L.; Nuvoloni, R.; Cerri, D.; Fratini, F.; Turchi, B. Pathotypes and Antimicrobial Susceptibility of Escherichia Coli Isolated from Wild Boar (Sus Scrofa) in Tuscany. Animals 2020, 10, 744. [Google Scholar] [CrossRef]
- Turchi, B.; Dec, M.; Bertelloni, F.; Winiarczyk, S.; Gnat, S.; Bresciani, F.; Viviani, F.; Cerri, D.; Fratini, F. Antibiotic Susceptibility and Virulence Factors in Escherichia Coli from Sympatric Wildlife of the Apuan Alps Regional Park (Tuscany, Italy). Microb. Drug Resist. 2019, 25, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, D.; Poeta, P.; Sáenz, Y.; Vinué, L.; Coelho, A.C.; Matos, M.; Rojo-Bezares, B.; Rodrigues, J.; Torres, C. Mechanisms of Antibiotic Resistance in Escherichia Coli Isolates Recovered from Wild Animals. Microb. Drug Resist. 2008, 14, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial Resistant Bacteria in Wild Mammals and Birds: A Coincidence or Cause for Concern? IRVETJ 2014, 67, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasyl, D.; Zając, M.; Lalak, A.; Skarżyńska, M.; Samcik, I.; Kwit, R.; Jabłoński, A.; Bocian, Ł.; Woźniakowski, G.; Hoszowski, A.; et al. Antimicrobial Resistance in Escherichia Coli Isolated from Wild Animals in Poland. Microb. Drug Resist. 2018, 24, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Beltrán, D.A.; Villar, D.; López-Osorio, S.; Ferguson, D.; Monsalve, L.K.; Chaparro-Gutiérrez, J.J. Prevalence of Antimicrobial Resistance in Bacterial Isolates from Dogs and Cats in a Veterinary Diagnostic Laboratory in Colombia from 2016–2019. Vet. Sci. 2020, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Liu, H.; Zhang, Z.; Pan, Y.; Zhao, Y. Mismatch between Antimicrobial Resistance Phenotype and Genotype of Pathogenic Vibrio Parahaemolyticus Isolated from Seafood. Food Control 2016, 59, 207–211. [Google Scholar] [CrossRef]
- Gow, S.P.; Waldner, C.L.; Harel, J.; Boerlin, P. Associations between Antimicrobial Resistance Genes in Fecal Generic Escherichia Coli Isolates from Cow-Calf Herds in Western Canada. Appl. Environ. Microb. 2008, 74, 3658–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guney, A.K. A Study on Class I Integrons and Antimicrobial Resistance among Clinical Staphylococci Isolates from a Turkish Hospital. Clin. Microbiol. 2014, 3. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.; Maurer, J.J.; Hofacre, C.; Summers, A.O. Gram-Positive Bacteria Are a Major Reservoir of Class 1 Antibiotic Resistance Integrons in Poultry Litter. Prod. Natl. Acad. Sci. USA 2004, 101, 7118–7122. [Google Scholar] [CrossRef] [Green Version]
- Skurnik, D. Effect of Human Vicinity on Antimicrobial Resistance and Integrons in Animal Faecal Escherichia Coli. J. Antimicrob. Chemother. 2006, 57, 1215–1219. [Google Scholar] [CrossRef]
- Di Cesare, A.; Eckert, E.M.; D’Urso, S.; Bertoni, R.; Gillan, D.C.; Wattiez, R.; Corno, G. Co-Occurrence of Integrase 1, Antibiotic and Heavy Metal Resistance Genes in Municipal Wastewater Treatment Plants. Water Res. 2016, 94, 208–214. [Google Scholar] [CrossRef]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y.-G. Using the Class 1 Integron-Integrase Gene as a Proxy for Anthropogenic Pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Nardelli, M.; Scalzo, P.M.; Ramírez, M.S.; Quiroga, M.P.; Cassini, M.H.; Centrón, D. Class 1 Integrons in Environments with Different Degrees of Urbanization. PLoS ONE 2012, 7, e39223. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Hughes, J.M. Critical Importance of a One Health Approach to Antimicrobial Resistance. EcoHealth 2019, 16, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segawa, T.; Takeuchi, N.; Rivera, A.; Yamada, A.; Yoshimura, Y.; Barcaza, G.; Shinbori, K.; Motoyama, H.; Kohshima, S.; Ushida, K. Distribution of Antibiotic Resistance Genes in Glacier Environments: Antibiotic Resistance Genes in Snow and Ice. Environ. Microbiol. Rep. 2013, 5, 127–134. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Target | Primer Sequence (5′–3′) | Annealing Temperature (°C) | Amplicon Size (bp) |
|---|---|---|---|
| tetA | GCTACATCCTGCTTGCCTTC CATAGATCGCCGTGAAGAGG | 58 | 210 |
| qnrS | GACGTGCTAACTTGCGTGAT TGGCATTGTTGGAAACTTG | 60 | 118 |
| sull II | TCCGGTGGAGGCCGGTATCTGG CGGGAATGCCATCTGCCTTGAG | 60 | 191 |
| blaTEM | TTCCTGTTTTTGCTCACCCAG CTCAAGGATCTTACCGCTGTTG | 58 | 112 |
| blaCTXM | CTATGGCACCACCAACGATA ACGGCTTTCTGCCTTAGGTT | 58 | 103 |
| int1 | GGCTTCGTGATGCCTGCTT CATTCCTGGCCGTGGTTCT | 55 | 148 |
| Class | Common Name (Scientific Name) | Number |
|---|---|---|
| Bird | Gray Heron (Ardea cinerea) | 2 |
| Tawny owl (Strix aluco) | 10 | |
| European scops owl (Otus scops) | 1 | |
| Golden eagle (Aquila chrysaetos) | 2 | |
| Barn Owl (Tyto alba) | 10 | |
| Egyptian vulture (Neophron percnopterus) | 1 | |
| White stork (Ciconia ciconia) | 2 | |
| Great cormorant (Phalacrocorax carbo) | 1 | |
| Hooded crow (Corvus cornix) | 1 | |
| Crow (Corvus corax) | 1 | |
| Lanner Falcon (Falco biarmicus) | 3 | |
| Peregrine Falcon (Falco peregrinus) | 10 | |
| Honey Buzzard (Pernis apivorus) | 13 | |
| Flamingo (Phoenicopterus ruber) | 6 | |
| Seagull (Larus ridibundus) | 15 | |
| Royal Seagull (Larus michahellis) | 20 | |
| Little Egret (Egretta garzetta) | 2 | |
| Magpie (Pica piza) | 2 | |
| Kestrel (Falco tinnunculus) | 27 | |
| Jay (Garrulus glandarius) | 2 | |
| Gryphon (Gyps fulvus) | 1 | |
| Bee-eater (Merops apiaster) | 1 | |
| Owl (Asio otus) | 1 | |
| Royal Kite (Milvus milvus) | 1 | |
| Buzzard (Buteo buteo) | 52 | |
| Squacco Heron (Ardeola ralloides) | 1 | |
| Great Crested Grebe (Podiceps cristatus) | 2 | |
| Little bittern (Ixobrychus minutus) | 1 | |
| Mallard (Anas platyrhynchos) | 1 | |
| Mammals | Wild boar (Sus scrofa) | 23 |
| Rabbit (Oryctolagus cuniculus) | 84 | |
| Deer (Dama dama) | 1 | |
| Porcupine (Hystrix cristata) | 2 | |
| Marten (Martes martes) | 1 | |
| Hedgehog (Erinaceus europaeus) | 4 | |
| Fox (Vulpes vulpes) | 3 | |
| Bat (Tadarita teniodis) | 1 | |
| Mallard (Anas platyrhynchos) | 1 | |
| Reptiles | Hermann’s tortoise (Testudo Hermanni) | 29 |
| Greek tortoise (Testudo graeca) | 4 | |
| Marginated tortoise (Testudo marginata) | 2 | |
| Pond slider (Trachemys scripta) | 19 | |
| Sicilian Pond Turtle (Emys trinacris) | 2 |
| Bacterial Genera | Number of Isolated Bacteria | Prevalence (%) | ||
|---|---|---|---|---|
| Birds | Mammals | Reptiles | ||
| Escherichia spp. | 114 | 23.3 | 52.9 | 10.7 |
| Clostridium spp. | 50 | 14 | 18.5 | 1.8 |
| Citrobacter spp. | 37 | 0.5 | 1.7 | 60.7 |
| Aeromonas spp. | 32 | 9.3 | 5.9 | 12.5 |
| Staphylococcus spp. | 27 | 11.4 | 1.7 | 5.4 |
| Pasteurella spp. | 15 | 12.6 | ||
| Streptococcus spp. | 12 | 3.1 | 4.2 | 1.8 |
| Enterobacter spp. | 8 | 2.1 | 3.4 | |
| Klebsiella spp. | 6 | 2.1 | 3.6 | |
| Pseudomonas spp. | 5 | 2.1 | 0.8 | |
| Salmonella spp. | 4 | 0.5 | 5.4 | |
| Campylobacter spp. | 3 | 0.5 | 3.6 | |
| Animal Species | Id | Bacterial Species | AMP | AMC | FUR | CRO | C | ENR | CIP | CN | AK | DA | SXT | TE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Birds (26 individuals) | ||||||||||||||
| Mallard | 1 | Escherichia coli | R | S | R | I | S | S | S | S | S | R a | S | I |
| Peregrine falcon | 4 | Staphylococcus aureus | R | S | S | S | S | R | R | S | S | S | S | S |
| Escherichia fergusonii | R | S | R | S | S | S | S | S | S | R a | S | I | ||
| Golden Eagle | 5 | Staphylococcus aureus | R | S | S | S | S | R | R | S | S | S | S | S |
| Escherichia coli | R | S | S | S | S | R | R | S | S | S b | S | S | ||
| Kestrel | 6 | Escherichia coli | R | S | S | S | S | S | S | S | S | R a | S | S |
| Honey Buzzard | 7 | Escherichia coli | R | S | S | S | S | S | S | S | S | R a | S | S |
| Buzzard | 8 | Staphylococcuschromogenes | S | S | S | S | S | S | S | S | S | I | S | S |
| European scops owl | 12 | Escherichia coli | R | S | S | S | S | S | S | S | I | R a | S | S |
| Owl | 13 | Yersinia nurmii | R | S | S | S | S | S | S | S | S | R a | S | S |
| Escherichia coli | R | S | S | S | S | S | S | S | I | R a | R | R | ||
| Peregrine falcon | 15 | Escherichia coli | R | I | S | S | S | S | S | S | S | R a | S | S |
| Barn owl | 16 | Clostridium perfringens | S | S | S | S | S | S | S | R | R | S | S | S |
| Kestrel | 17 | Escherichia coli | R | S | I | S | S | I | I | I | I | R a | S | S |
| Buzzard | 18 | Clostridium perfringens | S | S | S | S | S | S | S | R | R | S | S | S |
| Escherichia coli | I | S | I | S | S | S | S | S | S | R a | S | S | ||
| Tawny owl | 19 | Clostridium perfringens | S | S | S | S | S | S | S | R | R | S | S | I |
| Hafnia alvei | I | R | S | S | S | S | S | S | S | R a | S | R | ||
| Escherichia coli | R | S | I | S | S | S | S | I | S | R a | S | S | ||
| Peregrine falcon | 20 | Escherichia coli | I | R | S | S | S | S | S | S | S | R a | S | S |
| Hafnia alvei | I | S | S | S | S | S | S | S | S | R a | S | S | ||
| Klebsiella oxytoca | R a | R | S | S | S | S | S | S | S | R a | S | S | ||
| Kestrel | 23 | Clostridium perfringens | S | S | S | S | S | S | S | R | R | S | S | I |
| Kestrel | 24 | Pseudomonas aeruginosa | R a | R a | S | I b | I b | R | R | S | S | R a | R | I b |
| Flamingo | 26 | Escherichia coli | R | S | I | S | S | I | S | S | S | R a | S | R |
| Kestrel | 28 | Escherichia albertii | R | S | S | S | S | S | S | S | S | R a | S | S |
| Escherichia coli | R | S | R | S | S | S | S | I | R | R a | S | S | ||
| Staphylococcus warneri | S | S | S | S | S | S | S | S | S | S | S | S | ||
| Gray Heron | 29 | Salmonella enterica | R | S | I | S | S | S | S | S | S | R a | S | S |
| Crow | 30 | Staphylococcus aureus | I | S | I | S | S | S | S | S | S | S | S | S |
| Enterococcus faecium | S | S | R a | I b | S | S | S | S | S | S | Ra | S | ||
| Campylobacter jejuni | R | S | R | I | S | S | S | S | S | S | R | S | ||
| Peregrine falcon | 32 | Staphylococcus simulans | R | S | S | S | R | R | I | S | S | R | S | R |
| Peregrine falcon | 33 | Escherichia coli | R | S | I | S | S | S | S | I | R | Ra | S | S |
| Staphylococcuschromogenes | S | S | I | S | S | R | I | S | S | I | S | S | ||
| Pseudomonas aeruginosa | R a | R a | R | I b | S b | I | I | S | S | R | R | I b | ||
| Peregrine falcon | 34 | Escherichia fergusonii | R | S | I | S | S | S | S | I | R | R a | S | S |
| Pseudomonas aeruginosa | R a | R a | I | I b | S b | I | I | S | S | I b | R | I b | ||
| Bee-eater | 35 | Escherichia coli | R | S | I | S | S | S | S | S | I | R a | S | I |
| Royal Seagull | 38 | Streptococcus bovis | S | S | S | S | S | I | I | I b | R a | I | R | I |
| Royal Seagull | 39 | Escherichia coli | I | S | I | S | S | S | S | I | S | R a | S | S |
| Total resistant strains on 41 isolates | 22 | 3 | 5 | 0 | 1 | 6 | 3 | 4 | 7 | 2 | 6 | 4 | ||
| Mammals (9 individuals) | ||||||||||||||
| Deer | 2 | Escherichia coli | R | S | S | S | S | S | S | S | S | R a | S | S |
| Rabbit | 3 | Escherichia coli | R | S | S | S | S | S | S | S | I | R a | S | S |
| Fox | 9 | Klebsiella oxytoca | R a | S | S | S | S | S | S | S | S | R a | S | S |
| Rabbit | 14 | Escherichia coli | R | S | I | S | S | S | S | I | R | R a | S | S |
| Marten | 21 | Escherichia coli | R | S | S | S | S | S | S | S | S | R a | S | S |
| Rabbit | 22 | Escherichia coli | R | S | I | S | S | S | S | I | I | R a | S | S |
| Rabbit | 25 | Enterobacter cloacae | R | I | R | S | S | S | S | S | I | R a | R | R |
| Hedgehog | 27 | Citrobacter freundii | R a | R a | S | S | S | I | S | S | S | R a | S | S |
| Streptococcus dysgalactiae | S | S | S | S | S | I | I | S b | R a | I | S | S | ||
| Staphylococcus simulans | S | S | S | S | S | S | S | S | S | S | S | S | ||
| Wild boar | 31 | Escherichia fergusonii | R | S | I | S | S | S | S | S | S | R a | S | S |
| Citrobacter freundii | R a | S b | I | S | S | S | S | S | S | R a | S | S | ||
| Total resistant strains on 12 isolates | 7 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | ||
| Reptiles (4 individuals) | ||||||||||||||
| Herman’s tortoise | 10 | Escherichia coli | R | S | S | S | S | S | S | S | S | R a | R | R |
| Citrobacter braakii | R a | S b | S | S | S | S | S | S | S | R a | S | S | ||
| Salmonella tennessee | R | S | I | S | S | S | S | S | S | R a | S | S | ||
| Greek tortoise | 11 | Corynebacterium xerosis | S | S | S | S | S | S | S | S | S | I | S | S |
| Escherichia coli | R | I | R | S | S | S | S | S | S | R a | S | I | ||
| Sicilian Pond Turtle | 36 | Providencia rustigianii | S | S | S | S | S | S | S | S | S | R a | S | R |
| Klebsiella oxytoca | R a | S | I | S | S | S | S | S | I | R a | S | S | ||
| Sicilian Pond Turtle | 37 | Klebsiella oxytoca | R a | S | I | S | S | S | S | S | I | R a | S | S |
| Total resistant strains on 8 isolates | 3 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | ||
| Animal Species | Id | Bacterial Species | Resistance Genes | AMP | AMC | FUR | CRO | C | ENR | CIP | CN | AK | DA | SXT | TE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Golden Eagle | 5 | Escherichia coli | blaTEM, sullII, int1 | R | S | S | S | S | R | R | S | S | S b | S | S |
| Owl | 13 | Escherichia coli | tetA, blaTEM, sullII, int1 | R | S | S | S | S | S | S | S | I | R a | R | R |
| Peregrine falcon | 15 | Escherichia coli | blaTEM | R | I | S | S | S | S | S | S | S | R a | S | S |
| Flamingo | 26 | Escherichia coli | tetA, blaTEM, sullII | R | S | I | S | S | I | S | S | S | R a | S | R |
| Peregrine falcon | 33 | Escherichia coli | blaTEM | R | S | I | S | S | S | S | I | R | R a | S | S |
| Rabbit | 25 | Enterobacter cloacae | tetA, sullII, int1 | R | I | R | S | S | S | S | S | I | R a | R | R |
| Herman’s tortoise | 10 | Escherichia coli | blaTEM | R | S | S | S | S | S | S | S | S | R a | R | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambino, D.; Vicari, D.; Vitale, M.; Schirò, G.; Mira, F.; Giglia, M.L.; Riccardi, A.; Gentile, A.; Giardina, S.; Carrozzo, A.; et al. Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy. Microorganisms 2021, 9, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010203
Gambino D, Vicari D, Vitale M, Schirò G, Mira F, Giglia ML, Riccardi A, Gentile A, Giardina S, Carrozzo A, et al. Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy. Microorganisms. 2021; 9(1):203. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010203
Chicago/Turabian StyleGambino, Delia, Domenico Vicari, Maria Vitale, Giorgia Schirò, Francesco Mira, Maria La Giglia, Alessandra Riccardi, Antonino Gentile, Susanna Giardina, Anna Carrozzo, and et al. 2021. "Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy" Microorganisms 9, no. 1: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010203