
Yeasts and Lactic Acid Bacteria for Panettone Production: An Assessment of Candidate Strains

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
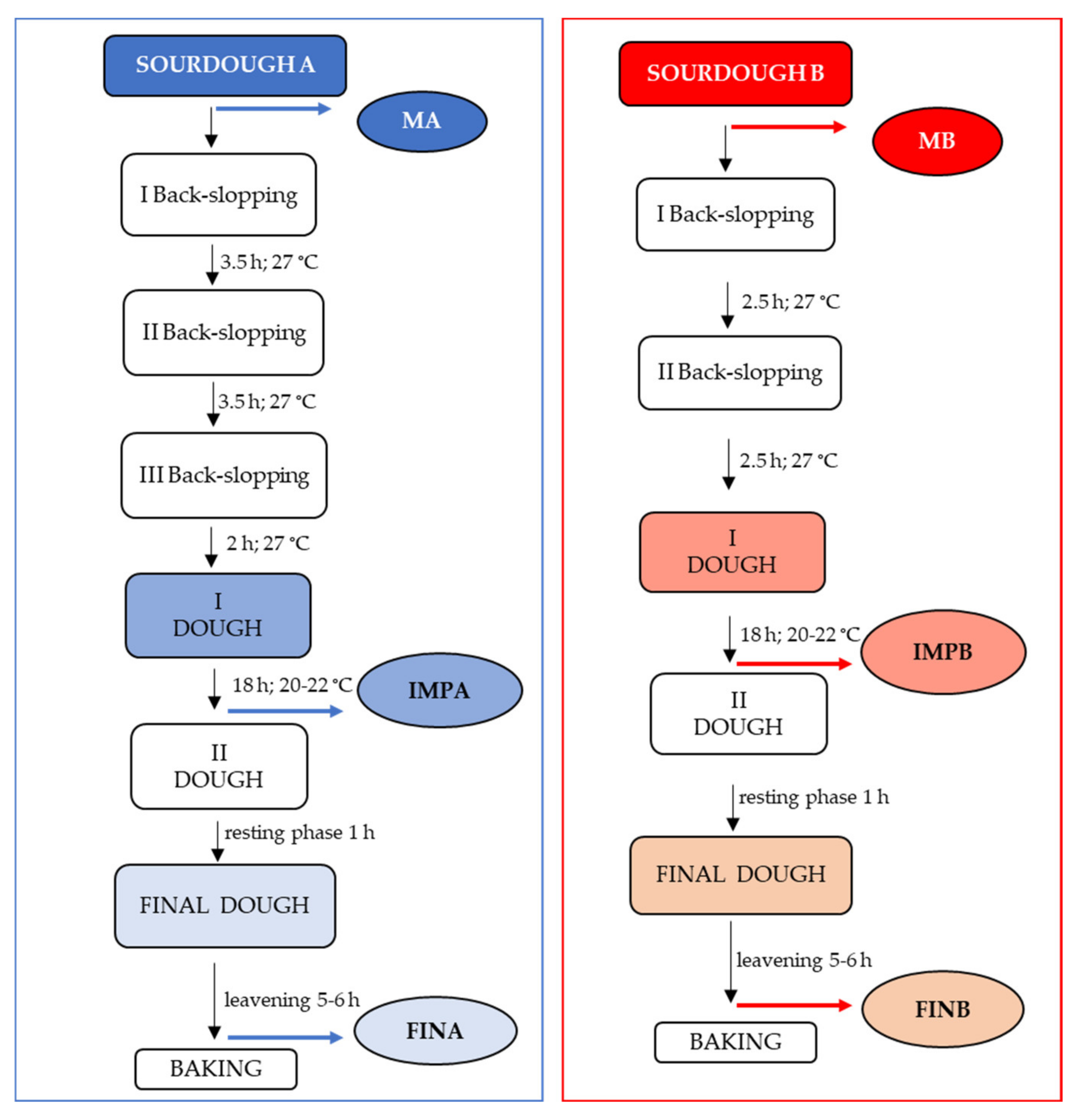
2.1. Panettone Production and Sample Collection
2.2. Determination of Physicochemical Parameters
2.3. Yeasts and LAB Isolation
2.4. DNA Extraction and Molecular Characterization
2.5. Species Assignment and Phylogenetic Clustering
2.6. HS-SPME-GC/MS Profiling of Fermented Products
2.7. Statistical Analysis
3. Results
3.1. Samples Analysis and Microbial Counts
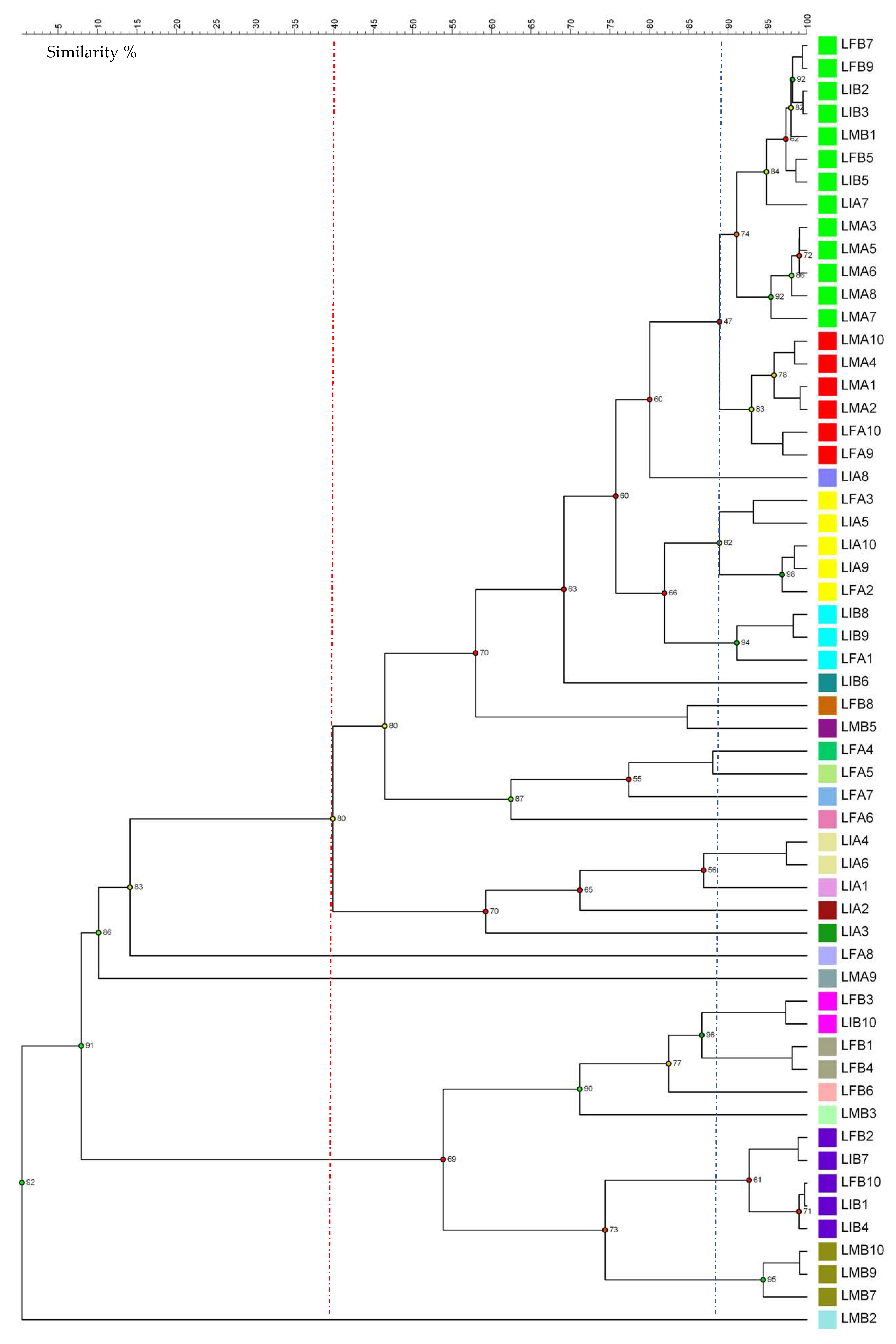
3.2. Yeasts’ Molecular Characterization and Phylogenetic Analysis
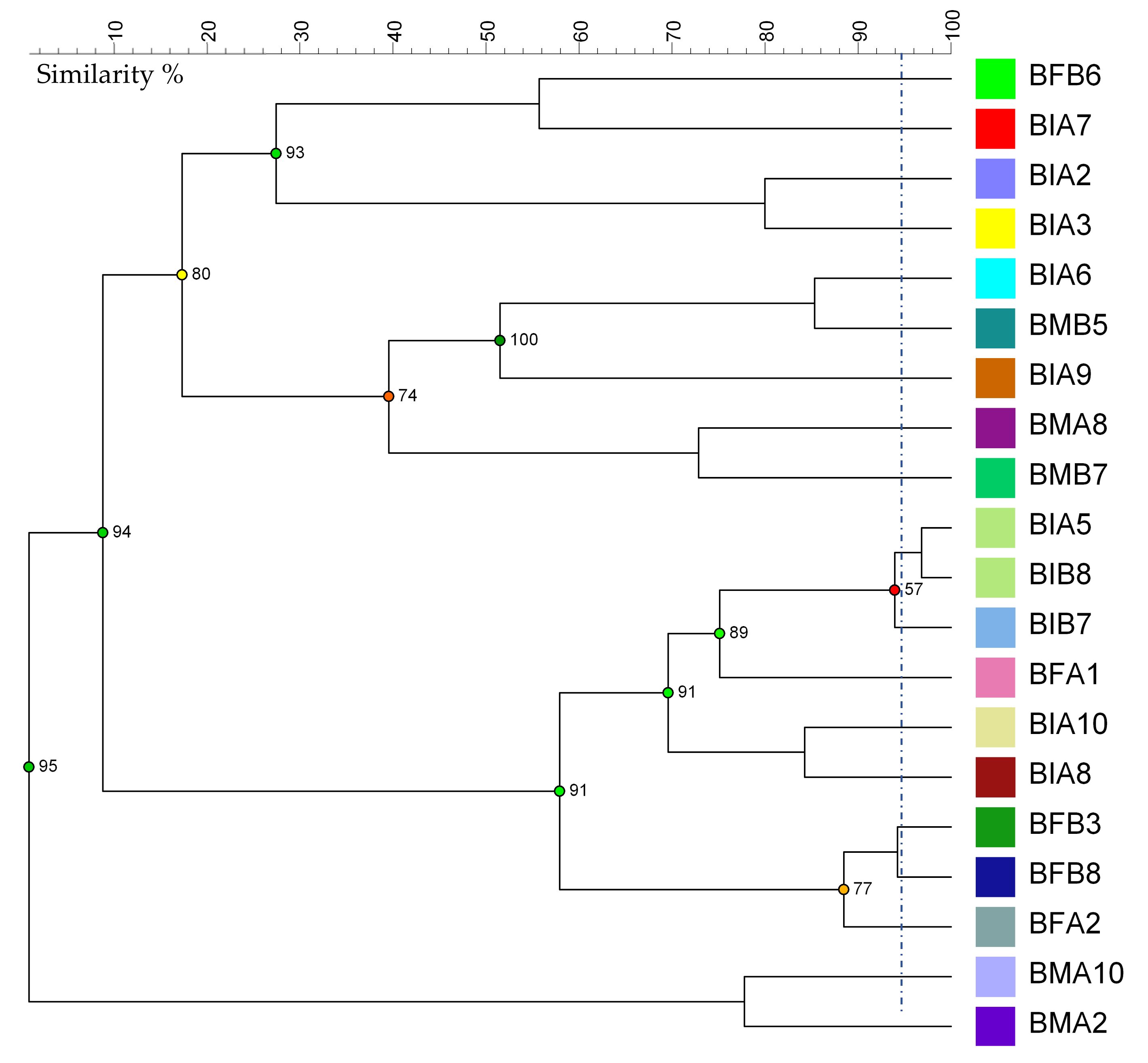
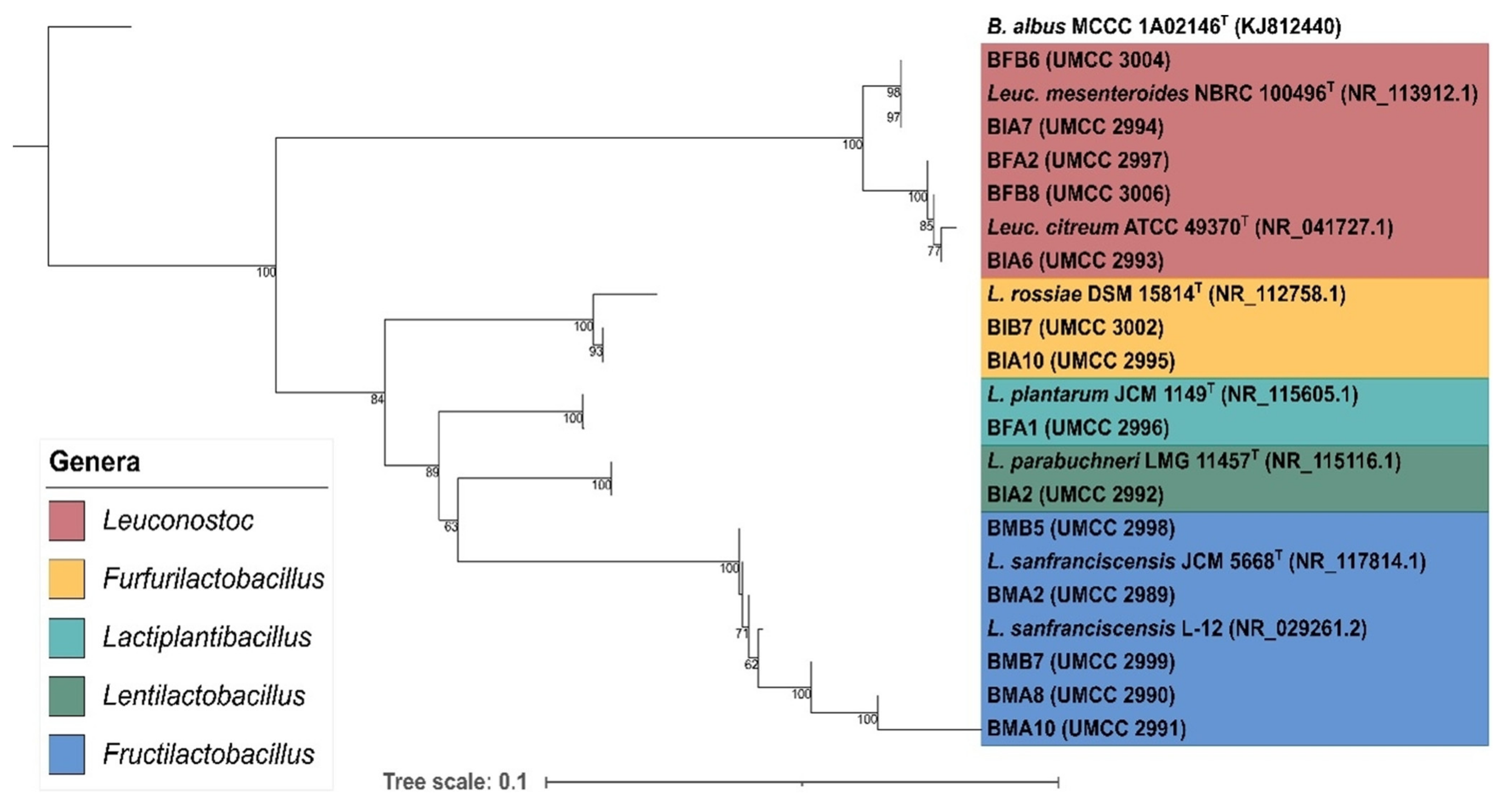
3.3. Bacteria Molecular Characterization and Phylogenetic Analysis
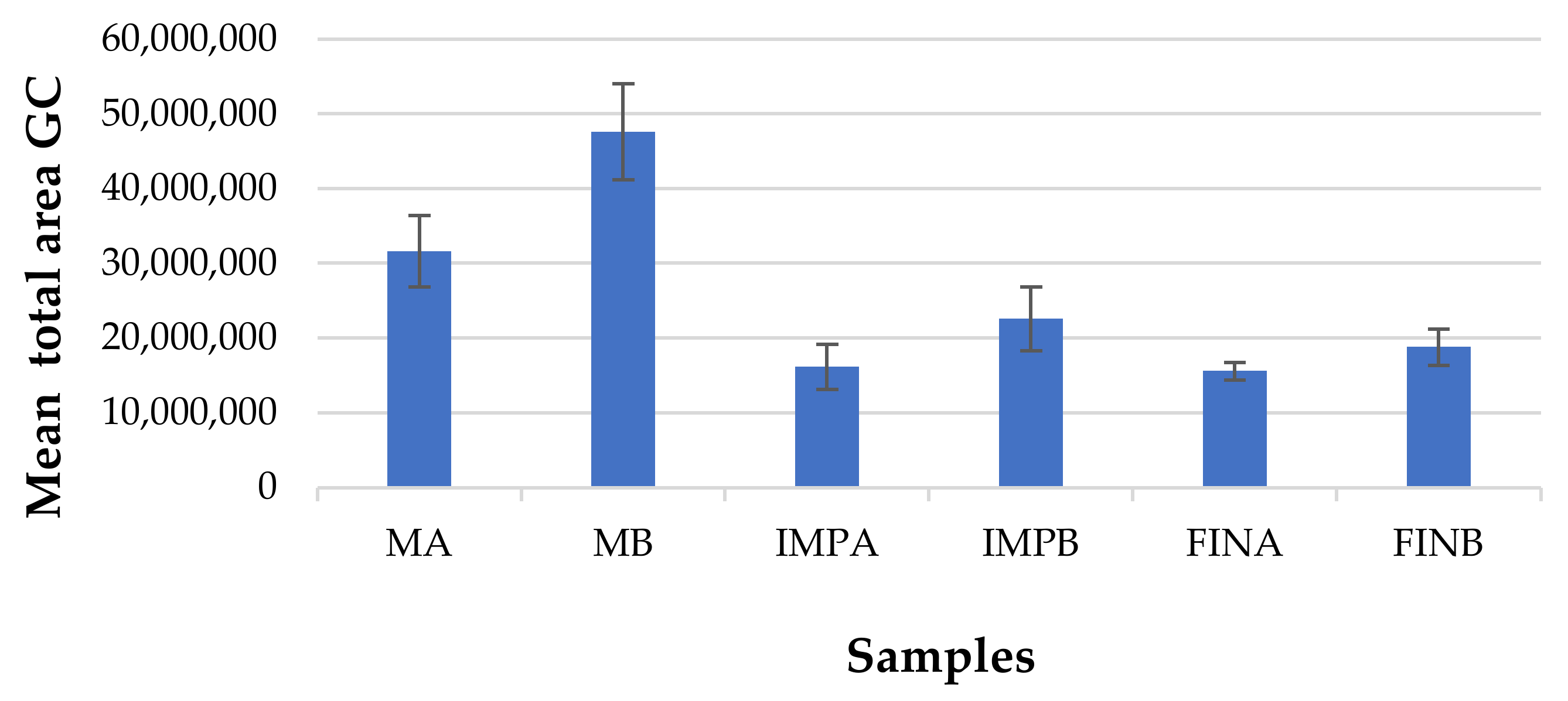
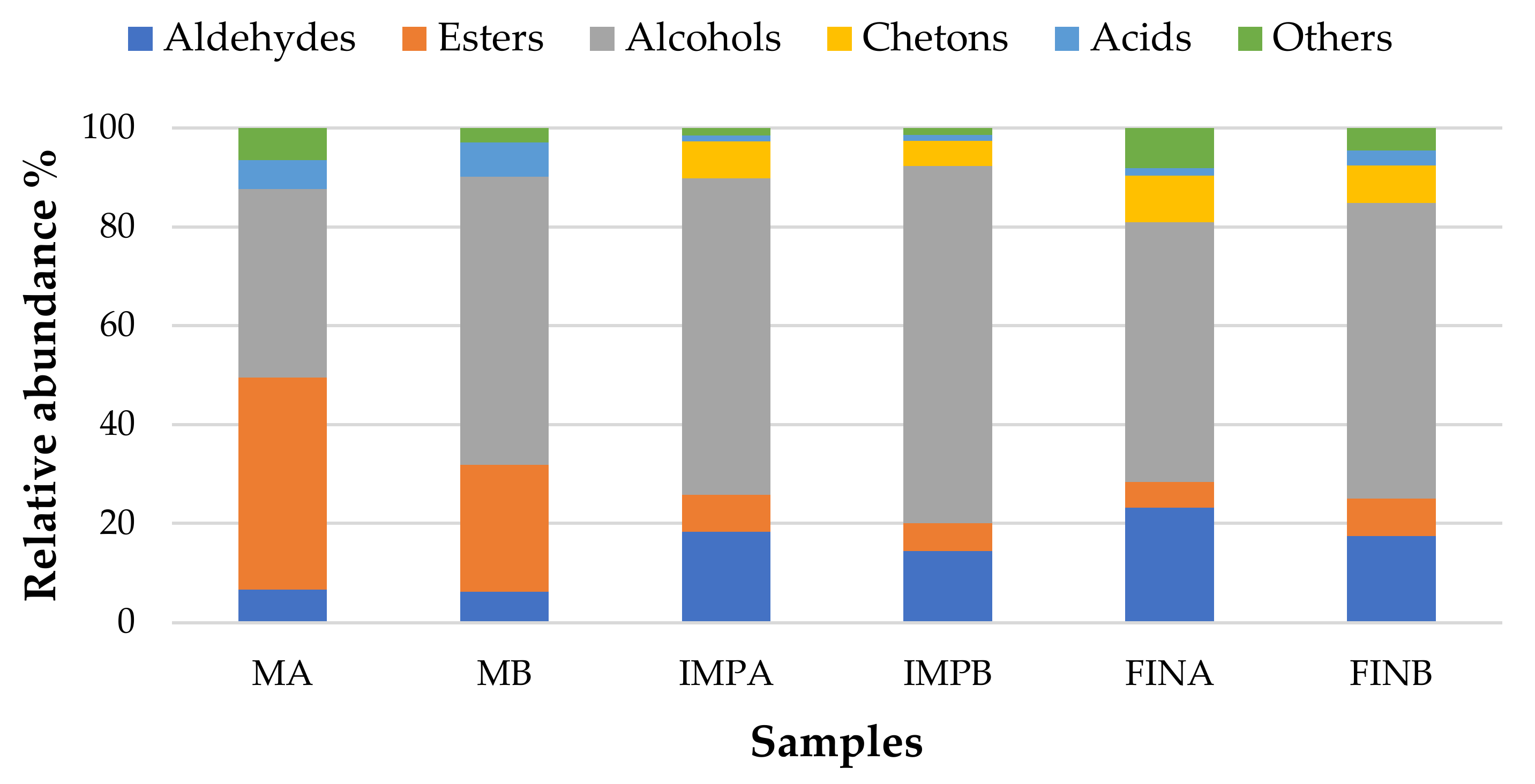
3.4. Volatile Organic Compounds Profiling
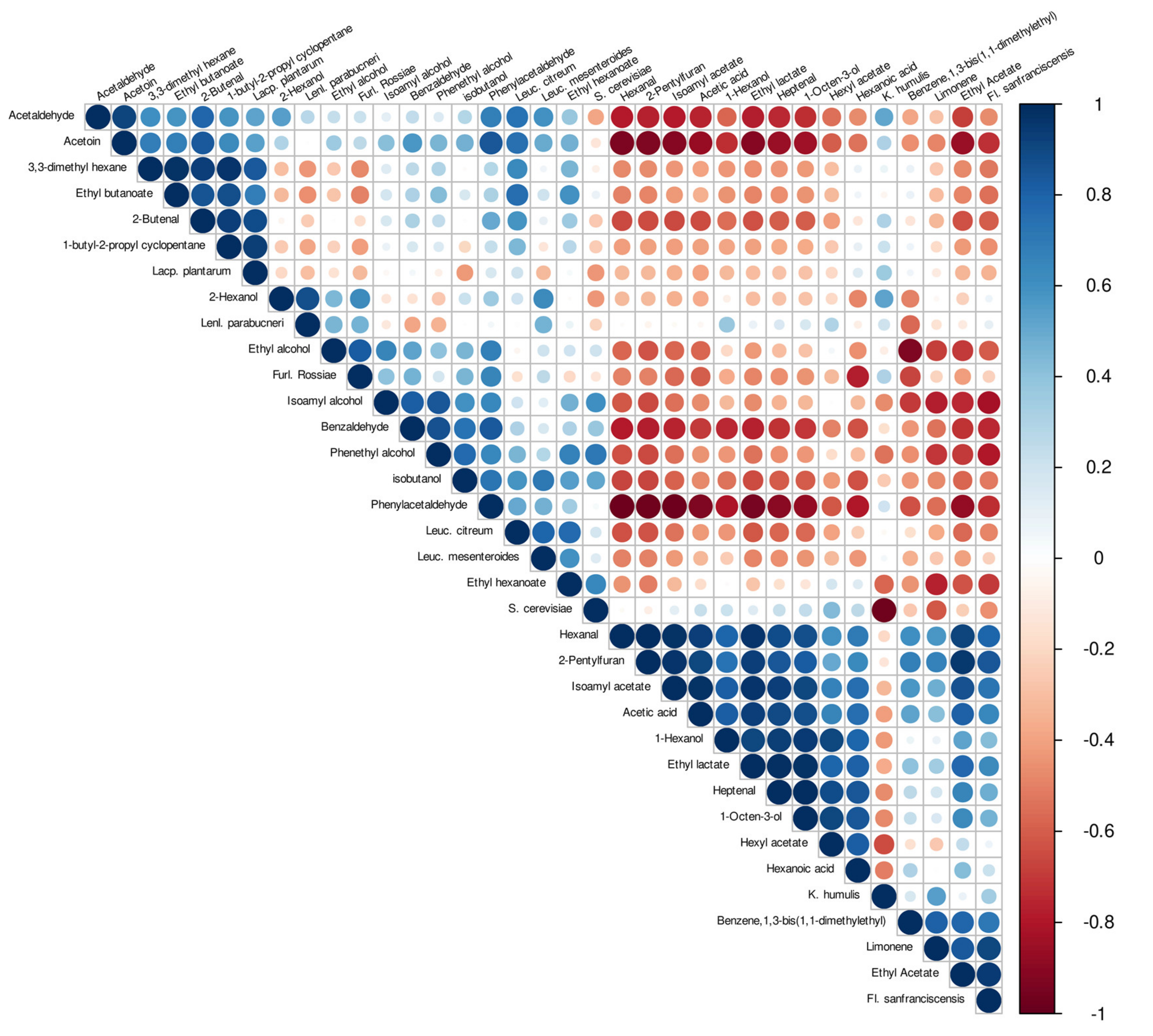
3.5. Correlation of VOCs and Species Detected in Sourdough and Dough Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vernocchi, P.; Valmorri, S.; Gatto, V.; Torriani, S.; Gianotti, A.; Suzzi, G.; Guerzoni, M.E.; Gardini, F. A Survey on yeast microbiota associated with an Italian traditional sweet-leavened baked good fermentation. Food Res. Int. 2004, 37, 469–476. [Google Scholar] [CrossRef]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The lactic acid bacteria and yeast microbiota of eighteen sourdoughs used for the manufacture of traditional Italian sweet leavened baked goods. Int. J. Food Microbiol. 2013, 163, 71–79. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Foschino, R.; Gallina, S.; Andrighetto, C.; Rossetti, L.; Galli, A. Comparison of Cultural Methods for the Identification and Molecular Investigation of Yeasts from Sourdoughs for Italian Sweet Baked Products. FEMS Yeast Res. 2004, 4, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Silvestri, G.; Aquilanti, L.; Clementi, F. PCR-DGGE Analysis of lactic acid bacteria and yeast dynamics during the production processes of three varieties of Panettone. J. Appl. Microbiol. 2008, 105, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, E.; Ehrmann, M.A.; Vogel, R.F. Role of Kazachstania humilis and Saccharomyces cerevisiae in the strain-specific assertiveness of Fructilactobacillus sanfranciscensis strains in rye sourdough. Eur. Food Res. Technol. 2020, 246, 1817–1827. [Google Scholar] [CrossRef]
- Venturi, M.; Guerrini, S.; Vincenzini, M. Stable and non-competitive association of Saccharomyces cerevisiae, Candida milleri and Lactobacillus sanfranciscensis during manufacture of two traditional sourdough baked goods. Food Microbiol. 2012, 31, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Pontonio, E.; Buchin, S.; De Angelis, M.; Lattanzi, A.; Valerio, F.; Gobbetti, M.; Calasso, M. Diversity of the lactic acid bacterium and yeast microbiota in the switch from firm- to liquid-sourdough fermentation. Appl. Environ. Microbiol. 2014, 80, 3161–3172. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckx, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Gobbetti, M.; Corsetti, A.; Rossi, J.; La Rosa, F.; De Vincenzi, S. Identification and clustering of lactic acid bacteria and yeasts from wheat sourdoughs of central Italy. Ital. J. Food. Sci. 1994, 1, 85–94. [Google Scholar]
- Gullo, M.; Romano, A.D.; Pulvirenti, A.; Giudici, P. Candida humilis—Dominant species in sourdoughs for the production of durum wheat bran flour bread. Int. J. Food Microbiol. 2003, 80, 55–59. [Google Scholar] [CrossRef]
- Pulvirenti, A.; Solieri, L.; Gullo, M.; De Vero, L.; Giudici, P. Occurrence and dominance of yeast species in sourdough. Lett. Appl. Microbiol. 2004, 38, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Chavan, R.S.; Chavan, S.R. Sourdough Technology-A Traditional Way for Wholesome Foods: A Review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 169–182. [Google Scholar] [CrossRef]
- Montanari, C.; Bargossi, E.; Lanciotti, R.; Chinnici, F.; Gardini, F.; Tabanelli, G. Effects of two different sourdoughs on the characteristics of Pandoro, a typical Italian sweet leavened baked good. LWT 2014, 59, 289–299. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Reale, A.; Di Renzo, T.; Boscaino, F.; Nazzaro, F.; Fratianni, F.; Aponte, M. Lactic acid bacteria biota and aroma profile of italian traditional sourdoughs from the irpinian area in Italy. Front. Microbiol. 2019, 10, 1621. [Google Scholar] [CrossRef] [Green Version]
- De Vero, L.; Iosca, G.; Gullo, M.; Pulvirenti, A. Functional and Healthy Features of Conventional and Non-Conventional Sourdoughs. Appl. Sci. 2021, 11, 3694. [Google Scholar] [CrossRef]
- Pulvirenti, A.; Rainieri, S.; Boveri, S.; Giudici, P. Optimizing the selection process of yeast starter cultures by preselecting strains dominating spontaneous fermentations. Can. J. Microbiol. 2009, 55, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Reale, A.; Zotta, T.; Ianniello, R.G.; Mamone, G.; Di Renzo, T. Selection criteria of lactic acid bacteria to be used as starter for sweet and salty leavened baked products. LTW 2020, 133, 110092. [Google Scholar] [CrossRef]
- Palla, M.; Blandino, M.; Grassi, A.; Giordano, D.; Sgerri, C.; Quartacci, M.F.; Reyneri, A.; Agnolucci, M.; Giovanetti, M. Characterization and selection of functional yeast strains during sourdough fermentation of different cereal wholegrain flours. Sci. Rep. 2020, 10, 12856. [Google Scholar] [CrossRef]
- Ercolini, D.; Pontonio, E.; De Filippis, F.; Minervini, F.; La Storia, A.; Gobbetti, M.; Di Cagno, R. Microbial ecology dynamics during rye and wheat sourdough preparation. Appl. Environ. Microbiol. 2013, 79, 7827–7836. [Google Scholar] [CrossRef] [Green Version]
- De Vero, L.; Boniotti, M.B.; Budroni, M.; Buzzini, P.; Cassanelli, S.; Comunian, R.; Gullo, M.; Logrieco, A.F.; Mannazzu, I.; Musumeci, R.; et al. Preservation, characterization and exploitation of microbial biodiversity: The perspective of the italian network of culture collections. Microorganisms 2019, 7, 685. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, C.S.; Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology: Preface, Ringbou ed.; John Wiley & Sons Inc: Hoboken, NJ, USA, 2003; ISBN 047150338X. [Google Scholar]
- Valcheva, R.; Kabadjova, P.; Rachman, C.; Ivanova, I.; Onno, B.; Prévost, H.; Dousset, X. A Rapid PCR procedure for the specific identification of Lactobacillus sanfranciscensis, based on the 16S–23S Intergenic Spacer Regions. J. Appl. Microbiol. 2007, 102, 290–302. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- La China, S.; De Vero, L.; Anguluri, K.; Brugnoli, M.; Mamlouk, D.; Gullo, M. Kombucha tea as a reservoir of cellulose producing bacteria: Assessing diversity among Komagataeibacter isolates. Appl. Sci. 2021, 11, 1595. [Google Scholar] [CrossRef]
- Kurtzman, C.P. Identification of food and beverage spoilage yeasts from DNA sequence analyses. Int. J. Food Microbiol. 2015, 213, 71–78. [Google Scholar] [CrossRef]
- Weiss, S.; Samson, F.; Navarro, D.; Casaregola, S. YeastIP: A database for identification and phylogeny of Saccharomycotina yeasts. FEMS Yeast Res. 2013, 13, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Yanagida, F.; Shinohara, T.; Yokotsuka, K. Restriction Fragment Length Polymorphism Analysis of 16S RRNA genes in lactic acid aacteria isolated from Red Wine. J. Biosci. Bioeng. 2000, 90, 335–337. [Google Scholar] [CrossRef]
- Higgins, D.; Thompson, J.; Gibson TThompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taiyun, W.; Viliam, S. R Package “corrplot”: Visualization of a Correlation Matrix: Version 0.84. 2017. Available online: https://github.com/taiyun/corrplot (accessed on 2 April 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 2 April 2021).
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of Yeasts by RFLP Analysis of the 5.8S rRNA Gene and the Two Ribosomal Internal Transcribed Spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subu.Pdf. Antonie Van Leeuwenhoek 1998, 98, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Lachance, M.A.; Wijayanayaka, T.M.; Bundus, J.D.; Wijayanayaka, D.N. Ribosomal DNA sequence polymorphism and the delineation of two ascosporic yeast species, Metschnikowia agaves and Starmerella bombicola. FEMS Yeast Res. 2011, 11, 324–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, N.; Sarilar, V.; Urien, C.; Lopes, M.R.; Morais, C.G.; Uetanabaro, A.P.T.; Tinsley, C.R.; Rosa, C.A.; Sicard, D.; Casaregola, S. Three novel ascomycetous yeast species of the Kazachstania clade, Kazachstania saulgeensis sp. nov., Kazachstania serrabonitensis sp. nov. and Kazachstania australis sp. nov. Reassignment of Candida humilis to Kazachstania humilis f.a. comb. nov. and Candida pseudohumilis to Kazachstania pseudohumilis f.a. comb. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5192–5200. [Google Scholar]
- Coppola, S.; Blaiotta, G.; Ercolini, D.; Moschetti, G. Molecular evaluation of microbial diversity occurring in different types of Mozzarella cheese. J. Appl. Microbiol. 2001, 90, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Rachman, C.N.; Kabadjova, P.; Prevost, H.; Dousset, X. Identification of Lactobacillus alimentarius and Lactobacillus farciminis with 16S-23S rDNA intergenic spacer region polymorphism and PCR amplification using species-specific oligonucleotide. J. Appl. Microbiol. 2003, 95, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Anaya, M.A. Enzymes and bread flavor. J. Agr. Food Chem. 1996, 44, 2469–2480. [Google Scholar] [CrossRef]
- Lund, B.; Hansen, A.; Lewis, M.J. The Influence of Yeast and Sourdough Fermentation on Volatile Compounds in Rye Bread Crumb. In Cereal Science and Technology, Proceedings from 23. Nordic Cereal Congress; Munck, L., Ed.; The Danish Cereal Society: Copenhagen, Denmark, 1987; pp. 185–195. [Google Scholar]
- Damiani, P.; Gobbetti, M.; Cossignani, L.; Corsetti, A.; Simonetti, M.S.; Rossi, J. The sourdough microflora. Characterization of hetero- and homo- fermentative lactic acid bacteria, yeasts and their interactions on the basis of the volatile compounds produced. LWT 1996, 29, 63–70. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast Diversity of Sourdoughs and Associated Metabolic Properties and Functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef]
- Carbonetto, B.; Nidelet, T.; Guezenec, S.; Perez, M.; Segond, D.; Sicard, D. Interactions between Kazachstania humilis yeast species and Lactic Acid Bacteria in Sourdough. Microorganisms 2020, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, A.; Ito, K.; Narushima, N.; Miyamoto, T. Identification of Lactic Acid Bacteria and Yeasts, and characterization of food components of Sourdoughs used in Japanese bakeries. J. Biosci. Bioeng. 2019, 127, 575–581. [Google Scholar] [CrossRef]
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Technol. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- Corsetti, A.; Lavermicocca, P.; Morea, M.; Baruzzi, F.; Tosti, N.; Gobbetti, M. Phenotypic and molecular identification and clustering of lactic acid bacteria and yeasts from wheat (species Triticum durum and Triticum aestivum) sourdoughs of Southern Italy. Int. J. Food Microbiol. 2001, 64, 95–104. [Google Scholar] [CrossRef]
- Rogalski, E.; Ehrmann, M.A.; Vogel, R.F. Strain-specific interaction of Fructilactobacillus sanfranciscensis with yeasts in the sourdough fermentation. Eur. Food Res. Technol. 2021, 247, 1437–1447. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L.; Van Sinderen, D.; Felis, G.E.; Dellaglio, F.; Gobbetti, M. Lactobacillus rossii sp. nov., isolated from Wheat Sourdough. Int. J. Syst. Evol. Microbiol. 2005, 55, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Di Cagno, R.; De Angelis, M.; Gallo, G.; Settanni, L.; Berloco, M.G.; Siragusa, S.; Parente, E. Genotypic and phenotypic diversity of Lactobacillus rossiae strains isolated from Sourdough. J. Appl. Microbiol. 2007, 103, 821–835. [Google Scholar] [CrossRef]
- Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. From Microbial ecology to innovative applications in food quality improvements: The Case of Sourdough as a Model Matrix. J. Multidiscip. Sci. J. 2020, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.; Schieberle, P. Generation of aroma compounds during sourdough fermentation: Applied and fundamental aspects. Trends Food Sci. Technol. 2005, 16, 85–94. [Google Scholar] [CrossRef]
- De Luca, L.; Aiello, A.; Pizzolongo, F.; Blaiotta, G.; Aponte, M.; Romano, R. Volatile Organic Compounds in Breads Prepared with Different Sourdoughs. Appl. Sci. 2021, 11, 1330. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | pH | Water Activity (aw) | Total Titratable Acidity (mL NaOH 0.1N/10 g) |
|---|---|---|---|
| MA | 3.90 ± 0.03 d | 0.0.9855 ± 0.002 d | 10.85 ± 0.45 b |
| IMPA | 5.44 ± 0.05 b | 0.0.9636 ± 0.001 b | 3.35 ± 0.25 a |
| FINA | 5.35 ± 0.02 bc | 0.0.9441 ± 0.002 a | 3.55 ± 0.45 a |
| MB | 3.81 ± 0.02 d | 0.0.9894 ± 0.001 d | 10.45 ± 0.45 b |
| IMPB | 5.30 ± 0.07 ac | 0.0.9562 ± 0.002 c | 3.45 ± 0.30 a |
| FINB | 5.21 ± 0.03 a | 0.0.9411 ± 0.003 a | 4.05 ± 0.05 b |
| Samples | Means Counts * on the Isolation Media | ||
|---|---|---|---|
| YPDA | MRS | SDB | |
| MA | 6.81 ± 0.25 b | 6.15 ± 0.14 ab | 6.17 ± 0.21 b |
| IMPA | 7.81 ± 0.05 a | 5.39 ± 0.12 a | 5.65 ± 0.21 ab |
| FINA | 7.83 ± 0.17 a | 5.46 ± 0.07 a | 5.45 ± 0.19 ab |
| MB | 7.56 ± 0.03 a | 6.87 ± 0.42 b | 6.16 ± 0.15 b |
| IMPB | 8.02 ± 0.01 a | 5.57 ± 0.06 a | 5.27 ± 0.09 a |
| FINB | 7.89 ± 0.17 a | 5.64 ± 0.04 a | 5.64 ± 0.02 ab |
| Sample | Strains * | Size of Amplicon (bp) | Amplicons Digestion with HaeIII (Size Fragments, bp) |
|---|---|---|---|
| MA | LMA2, LMA3 (UMCC 2973), LMA1, LMA4, LMA5 (UMCC 2974), LMA6, LMA7, LMA8, LMA9 (UMCC 2975), LMA10 | 630 | 400–230 |
| IMPA | LIA1 (UMCC 2976), LIA9, LIA10, LIA2, LIA3, LIA4, LIA5 (UMCC 2977), LIA7, LIA6 (UMCC 2978), LIA8 | 630 | 400–230 |
| FINA | LFA1 (UMCC 2979), LFA2, LFA3 (UMCC 2980), LFA4, LFA5, LFA6 (UMCC 2981), LFA7, LFA8, LFA9, LFA10 | 630 | 400–230 |
| MB | LMB1 (UMCC 2982), LMB5 | 630 | 400–230 |
| LMB2, LMB3, LMB7, LMB9 (UMCC 2984), LMB10 | 880 | 320–230–180–150 | |
| IMPB | LIB1, LIB4, LIB7 (UMCC 2986), LIB10 | 880 | 320–230–180–150 |
| LIB2, LIB3, LIB5 (UMCC 2985), LIB6, LIB8, LIB9 | 880 | 400–230 | |
| FINB | LFB1 (UMCC 2987), LFB2, LFB3, LFB4, LFB6, LFB10 (UMCC 2988) | 880 | 320–230–180–150 |
| LFB5, LFB7, LFB8, LFB9 | 630 | 400–230 |
| Sample | Strain * | 16S-23S rDNA ISR Size Fragments (bp) |
|---|---|---|
| MA | BMA2 (UMCC 2989), BMA8 (UMCC 2990), BMA10 (UMCC 2991) | 600–700–800 |
| IMPA | BIA2 (UMCC 2992), BIA3, BIA5, BIA6 (UMCC 2993), BIA7 (UMCC 2994), BIA8, BIA9, BIA10 (UMCC 2995) | 600–800 |
| FINA | BFA1 (UMCC 2996), BFA2 (UMCC 2997) | 600–800 |
| MB | BMB5 (UMCC 2998), BMB7 (UMCC 2999) | 600–700–800 |
| IMPB | BIB7 (UMCC 3002), BIB8 | 600–800 |
| FINB | BFB3, BFB6 (UMCC 3004), BFB8 (UMCC 3006) | 600–800 |
| Chemical Compounds | Samples * | |||||
|---|---|---|---|---|---|---|
| MA | MB | ImpA | ImpB | FinA | FinB | |
| Acetaldehyde | 2.22 ± 0.11 | 1.62 ± 0.13 | 8.43 ± 0.19 | 3.28 ± 0.01 | 8.31 ± 0.10 | 6.27 ± 0.05 |
| 2-Butenal | - | - | 2.44 ± 0.16 | 1.56 ± 0.23 | 8.61 ± 0.65 | 3.80 ± 0.26 |
| Hexanal | 4.13 ± 0.17 | 3.86 ± 0.37 | - | - | - | - |
| Heptenal | 0.31 ± 0.04 | 0.66 ± 0.06 | - | - | - | - |
| Benzaldehyde | - | - | 0.55 ± 0.06 | 1.96 ± 0.12 | 0.86 ± 0.21 | 1.37 ± 0.05 |
| Phenylacetaldehyde | - | - | 6.80 ±0.47 | 7.62 ± 0.18 | 5.52 ± 0.33 | 6.08 ± 0.03 |
| Aldehydes | 6.34 | 6.14 | 18.22 | 14.43 | 23.30 | 17.52 |
| Ethyl Acetate | 39.73 ± 1.51 | 20.78 ± 0.43 | 7.17 ± 0.45 | 5.43 ± 0.23 | 4.08 ± 0.04 | 5.85 ± 0.15 |
| Ethyl butanoate | - | - | - | - | 0.86 ± 0.12 | 0.76 ± 0.13 |
| Isoamyl acetate | 2.03 ± 0.14 | 2.17 ± 0.23 | - | - | - | 0.39 ± 0.12 |
| Ethyl hexanoate | - | 0.38 ± 0.07 | 0.31 ± 0.00 | 0.20 ± 0.03 | 0.32 ± 0.02 | 0.64 ± 0.07 |
| Hexyl acetate | - | 0.26±0.01 | - | - | - | - |
| Ethyl lactate | 1.44 ± 0.04 | 2.14 ± 0.01 | - | - | - | - |
| Esters | 43.19 | 25.74 | 7.48 | 5.64 | 5.26 | 7.26 |
| Ethyl alcohol | 31.19 ± 0.22 | 44.05 ± 0.00 | 50.77 ± 0.66 | 53.19 ± 2.50 | 41.09 ± 2.65 | 40.51 ± 1.46 |
| Isobutanol | - | - | 1.40 ± 0.17 | 1.75 ± 0.11 | - | 2.51 ± 0.05 |
| Isoamyl alcohol | 4.30 ± 0.25 | 6.80 ± 0.32 | 6.42 ± 0.56 | 8.92 ± 1.06 | 6.70 ± 1.32 | 7.77 ± 0.25 |
| 1-Hexanol | 1.96 ± 0.15 | 3.48 ± 0.42 | 1.60 ± 0.23 | 0.95 ± 0.12 | 1.12 ± 0.01 | 1.25 ± 0.06 |
| 2-Hexanol | - | - | 0.64 ± 0.11 | - | - | - |
| 1-Octen-3-ol | 0.30 ± 0.03 | 0.69 ± 0.03 | - | - | - | - |
| Phenethyl alcohol | 0.51 ± 0.01 | 3.27 ± 0.45 | 2.70 ± 0.38 | 7.44 ± 1.09 | 3.90 ± 0.62 | 8.22 ± 1.96 |
| Alcohols | 38.27 | 58.30 | 63.52 | 72.26 | 52.82 | 60.25 |
| Acetic acid | 5.34 ± 1.44 | 5.93 ± 0.09 | 1.23 ± 0.05 | 1.19 ± 0.09 | 1.11 ± 0.37 | 2.76 ± 0.35 |
| Hexanoic acid | 0.43 ± 0.07 | 0.94 ± 0.04 | - | - | 0.45 ± 0.12 | 0.31 ± 0.01 |
| Acids | 5.76 | 6.87 | 1.23 | 1.19 | 1.11 | 2.76 |
| Limonene | 2.49 ± 0.07 | 0.74 ± 0.01 | 1.08 ± 0.24 | 0.75 ± 0.11 | 0.96 ± 0.20 | 0.67 ± 0.06 |
| Acetoin | - | - | 7.42 ± 0.37 | 5.05 ± 0.33 | 9.39 ± 0.26 | 7.64 ±0.91 |
| 3,3-dimethyl hexane | - | - | - | - | 2.48 ± 0.36 | 1.47 ± 0.19 |
| 1-butyl-2-propyl cyclopentane | - | - | - | - | 3.24 ± 0.67 | 1.00 ± 0.40 |
| 2-Pentylfuran | 0.92 ± 0.00 | 0.72 ± 0.02 | - | - | - | - |
| Benzene,1,3-bis(1,1-dimethylethyl) | 3.02 ± 0.28 | 1.03 ± 0.14 | 0.38 ± 0.06 | 0.69 ± 0.12 | 1.44 ± 0.02 | 1.44 ± 0.04 |
| Others | 6.43 | 2.49 | 8.89 | 6.49 | 17.51 | 12.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Vero, L.; Iosca, G.; La China, S.; Licciardello, F.; Gullo, M.; Pulvirenti, A. Yeasts and Lactic Acid Bacteria for Panettone Production: An Assessment of Candidate Strains. Microorganisms 2021, 9, 1093. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051093
De Vero L, Iosca G, La China S, Licciardello F, Gullo M, Pulvirenti A. Yeasts and Lactic Acid Bacteria for Panettone Production: An Assessment of Candidate Strains. Microorganisms. 2021; 9(5):1093. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051093
Chicago/Turabian StyleDe Vero, Luciana, Giovanna Iosca, Salvatore La China, Fabio Licciardello, Maria Gullo, and Andrea Pulvirenti. 2021. "Yeasts and Lactic Acid Bacteria for Panettone Production: An Assessment of Candidate Strains" Microorganisms 9, no. 5: 1093. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051093