
Wastewater from the Edible Oil Industry as a Potential Source of Lipase- and Surfactant-Producing Actinobacteria


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. Physicochemical Analyses
2.3. Culture-Dependent Enumeration of Heterotrophic Bacteria and Isolation of Actinobacteria
2.4. Identification of Actinobacteria
2.5. Amplicon Sequencing
2.6. Genome Sequencing and Analysis
2.7. Lipase Activity and Surfactant Production
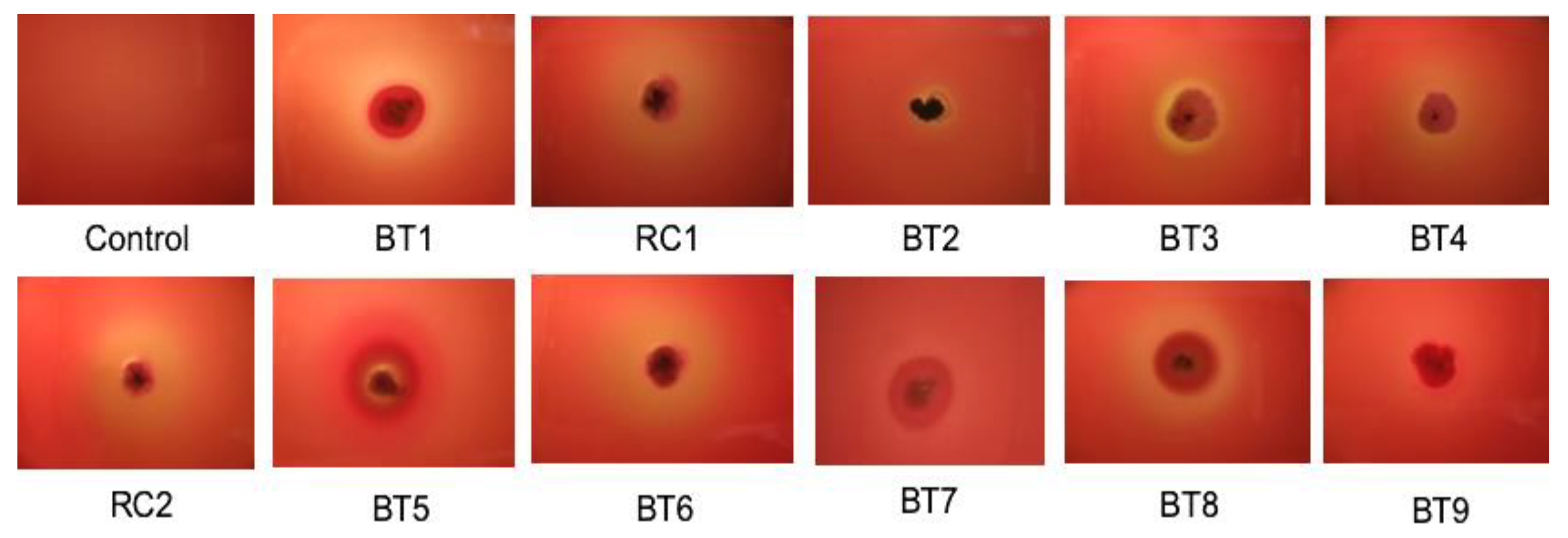
2.7.1. Screening Tests for Surfactant Production
2.7.2. Screening Tests for Lipase Production
2.7.3. Lipase Activity Assays
2.8. Statistical Analyses
3. Results
3.1. Qualitative Analysis of Edible Oil Wastewater Streams
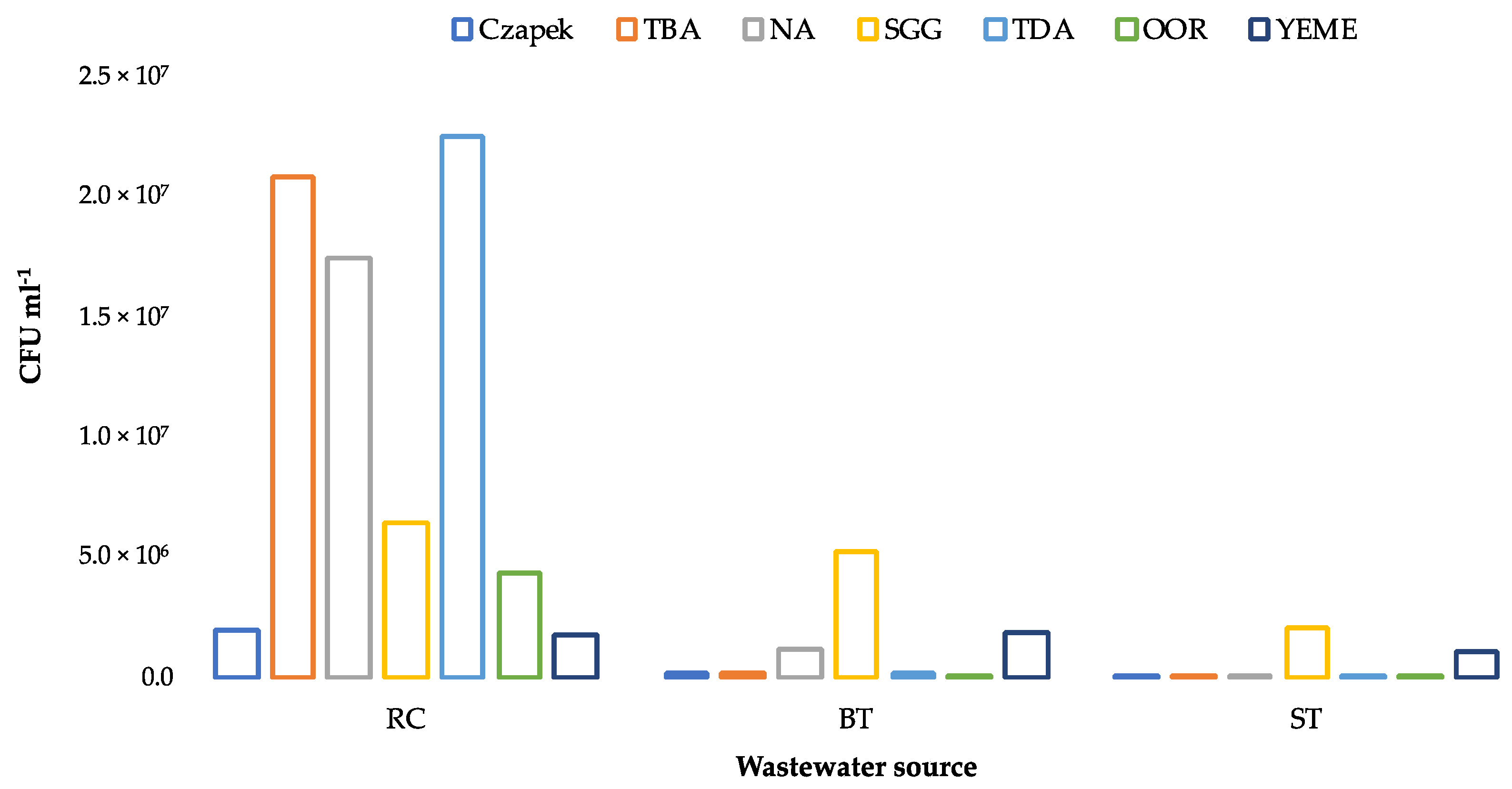
3.2. Enumeration of Culturable Heterotrophic Bacteria
3.3. Identification of Actinobacteria and Screening for Lipase and Biosurfactant Activity
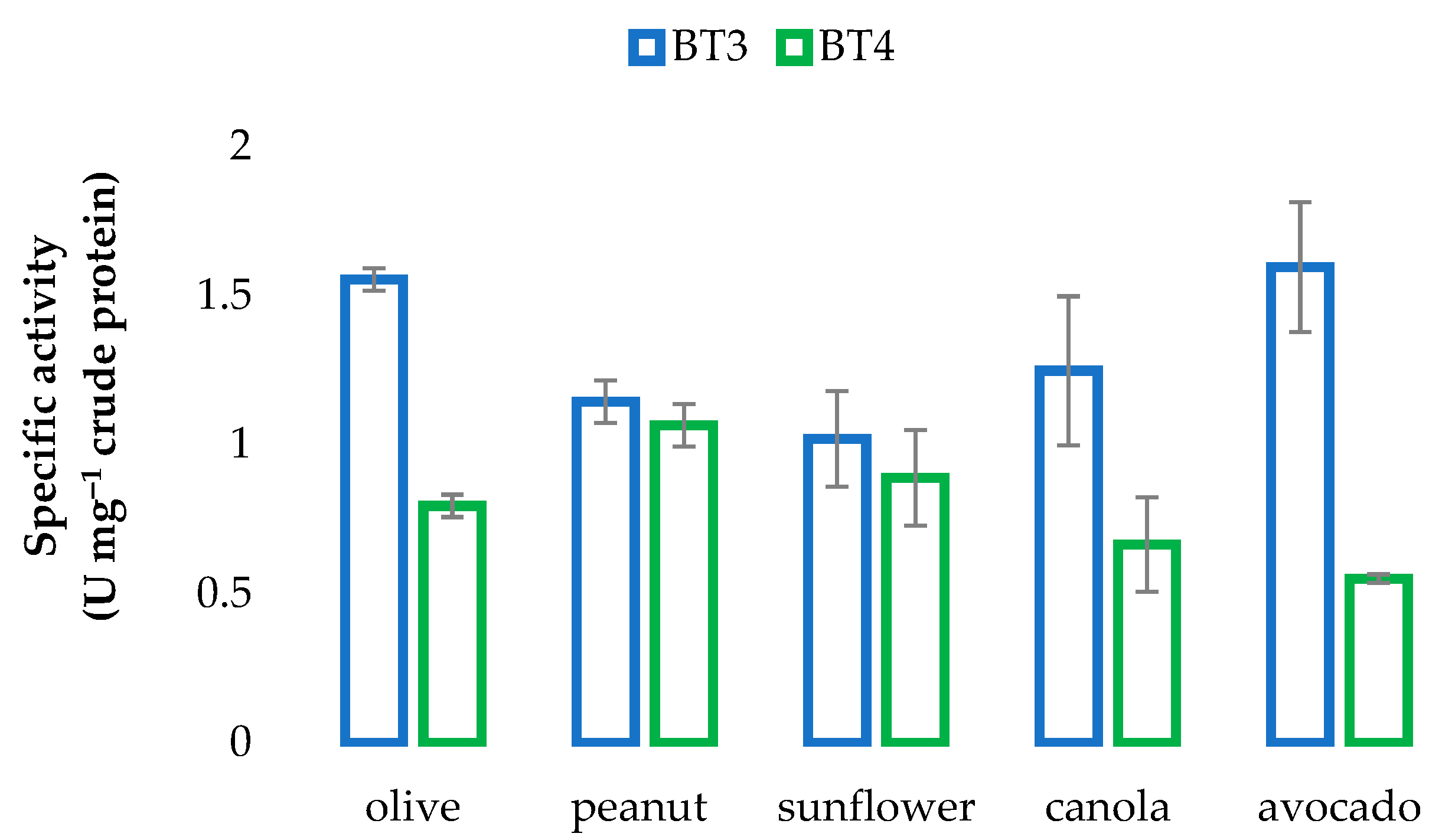
3.4. Determination of Lipase Activity of Actinobacterial Isolates
3.5. Genome Sequencing
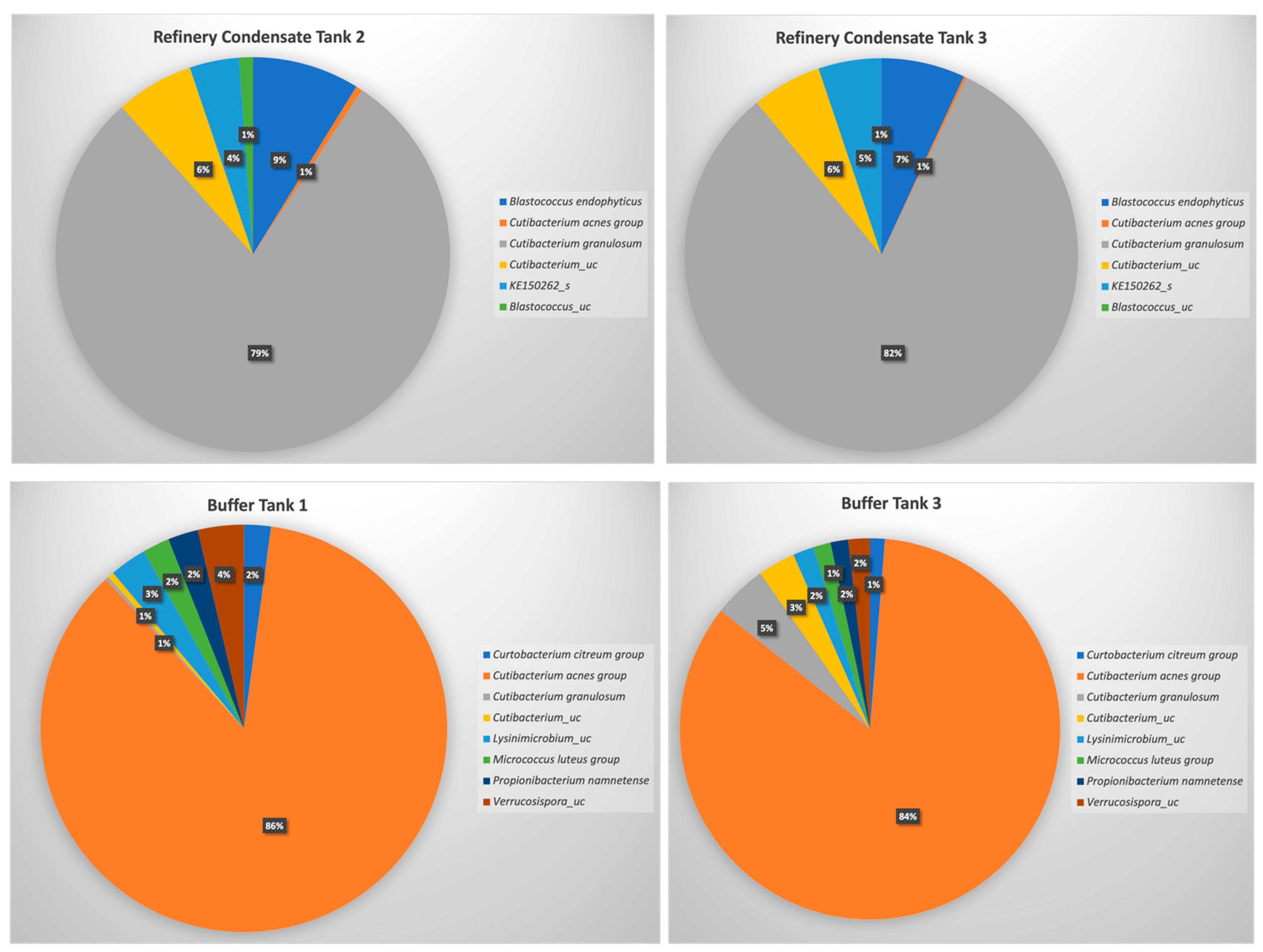
3.6. Microbiota Analysis for Selected Wastewater Streams
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verger, R. ‘Interfacial activation’ of lipases: Facts and artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Kis, A.; Laczi, K.; Zsíros, S.; Rákhely, G.; Perei, K. Biodegradation of animal fats and vegetable oils by Rhodococcus erythropolis PR4. Int. Biodeter. Biodegrad. 2015, 105, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Castilla, A.; Panizza, P.; Rodriguez, D.; Bonino, L.; Díaz, P.; Irazoqui, G.; Giordano, S.R. A novel thermophilic esterase from Janibacter sp. R02, the first member of a new lipase family (Family XVII). Enzym. Microb. Technol. 2017, 98, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Colin, V.L.; Bourguignon, N.; Gómez, J.S.; de Carvalho, K.G.; Ferrero, M.A.; Amorosó, M.J. Production of surface-active compounds by a hydrocarbon-degrading actinobacterium: Presumptive relationship with lipase activity. Water Air Soil Pollut. 2017, 228, 454. [Google Scholar] [CrossRef]
- Furini, G.; Berger, J.S.; Campos, J.A.M.; Van der Sand, S.T.; Germani, J.C. Production of lipolytic enzymes by bacteria isolated from biological effluent treatment systems. Ann. Braz. Acad. Sci. 2018, 90, 2955–2965. [Google Scholar] [CrossRef] [PubMed]
- Ficarra, F.A.; Santecchia, I.; Lagorio, S.H.; Alarcόn, S.; Magni, C.; Espariz, M. Genome mining of lipolytic exoenzymes from Bacillus safensis S9 and Pseudomonas alcaliphilia ED1 from a dairy wastewater lagoon. Arch. Microbiol. 2016, 198, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Silva-Bedoya, L.M.; Sánchez-Pinzón, S.; Cadavid-Restrepo, G.E.; Moreno-Herrera, C.X. Bacterial community analysis of an industrial wastewater treatment plant in Colombia with screening for lipid-degrading microorganisms. Microbiol. Res. 2016, 192, 313–325. [Google Scholar] [CrossRef]
- Sutrisno, A.; Wardani, A.K.; Ratnawati, L. Isolation and identification of lipid-degrading bacteria from fish (Sardinella longiceps) canning wastewater. Int. J. Adv. Sci. Eng. Inf. Technol. 2016, 6, 441–446. [Google Scholar] [CrossRef]
- Bestari, N.C.S. Activity assay and identification of lipolytic bacteria from wastewater fish industry at District Muncar, Banyuwangi Indonesia. Int. J. ChemTech Res. 2015, 8, 377–383. [Google Scholar]
- Petropolos, E.; Dolfing, J.; Yu, Y.; Wade, M.J.; Bowen, E.J.; Davenport, R.J.; Curtis, T.P. Lipolysis of domestic wastewater in anaerobic reactors operating at low temperatures. Environ. Sci. Water Res. Technol. 2018, 4, 1002–1013. [Google Scholar] [CrossRef] [Green Version]
- Garode, A.M.; Sonune, N.A. Screening and identification of lipase bacteria from domestic wastewater. Int. J. Pharma. Biol. Sci. 2014, 5, 265–268. [Google Scholar]
- Remmas, N.; Melidis, P.; Voltsi, C.; Athanasiou, D.; Ntougias, S. Novel hydrolytic halotolerant alkaliphiles from mature landfill leachate with key involvement in maturation process. J. Environ. Sci. Health 2017, 52, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Aktar, L.; Khan, F.I.; Islam, T.; Mitra, S.; Saha, M.L. Isolation and characterisation of indigenous lipase producing bacteria from lipid rich environment. Plant Tissue Cult. Biotechnol. 2016, 26, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Hozzein, W.N.; Wadaan, M.A.M.; Tawab, M.S.A.; Ahmed, M.B. Quantitative screening of some wastewater actinomycetes for production of industrial enzymes. J. Pure Appl. Microbiol. 2013, 7, 483–487. [Google Scholar]
- Welz, P.J. Edible seed oil waste: Status quo and future perspectives. Water Sci. Technol. 2019, 80, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Aslan, S.; Alyuz, B.; Bozkurt, Z.; Bakaoglu, M. Characterisation and biological treatability of edible oil wastewaters. Pol. J. Environ. Stud. 2009, 18, 533–538. [Google Scholar]
- Pintor, A.M.A.; Vilar, V.J.P.; Botleho, C.M.S.; Boaventura, R.A.R. Optimization of a primary graving separation treatment of vegetable oil refinery wastewater. Clean Technol. Environ. 2014, 16, 1725–1734. [Google Scholar] [CrossRef]
- Roux-van der Merwe, M.P.; Badenhorst, J.; Britz, T.J. Fungal treatment of an edible-oil containing industrial effluent. World J. Microbiol. Biotechnol. 2005, 21, 947–953. [Google Scholar] [CrossRef]
- American Public Health Association (APHA); American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA: Denver, CO, USA, 1999; ISBN 0-875533-235-7. [Google Scholar]
- Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl. Environ. Microbiol. 1999, 65, 4715–4724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, A.E.; Meyers, P.R. Rapid identification of filamentous actinomycetes to the genus level using genus-specific 16S rRNA gene restriction fragment patterns. Int. J. Syst. Evol. Microbiol. 2003, 53, 1907–1915. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Schäfer, J.; Jäckel, U.; Kämpfer, P. Development of a new PCR primer system for the selective amplification of Actinobacteria. FEMS Microbiol. Lett. 2010, 311, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandel, M.; Marmur, J. Use of ultraviolet absorbance-temperature profile for determining the guanine plus cytosine content of DNA. In Methods in Enzymology; Grossman, L., Moldave, K., Eds.; Academic Press: Cambridge, MA, USA, 1968; Volume XII, pp. 195–206. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Ha, S.M.; Kim, C.K.; Roh, J.; Byun, J.H.; Yang, S.J.; Choi, S.B.; Chun, J.; Yong, D. Application of the whole genome-based bacterial identification system, TrueBac ID, in clinical isolates which were not identified with three MALDI-TOF M/S systems. Ann. Lab. Med. 2019, 39, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrillo, P.G.; Mardaraz, C.; Pitta-Alvarez, S.J.; Giulietti, A.M. Isolation and selection of biosurfactant-producing bacteria. World J. Microbiol. Biotechnol. 1996, 12, 82–84. [Google Scholar] [CrossRef]
- Youssef, N.H.; Duncan, K.E.; Nagle, D.P.; Savager, K.N.; Knapp, R.M.; McInerney, M.J. Comparison of methods to detect biosurfactant production by diverse microorganisms. J. Microbiol. Meth. 2004, 56, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Morikova, M.; Hirata, Y.; Imanaka, T. A study on the structure–function relationship of the lipopeptide biosurfactants. Biochim. Biophys. Acta 2000, 1488, 211–218. [Google Scholar] [CrossRef]
- Bosch, M.P.; Robert, M.; Mercade, M.E.; Espuny, M.J.; Parra, J.L.; Guinea, J. Surface-active compounds on microbial cultures. Tenside Surfactants Deterg. 1988, 25, 208–211. [Google Scholar] [CrossRef]
- Palacios, D.; Busto, M.D.; Ortega, N. Study of a new spectrophotometric end-point assay for lipase activity determination in aqueous media. LWT—Food Sci. Technol. 2014, 55, 536–542. [Google Scholar] [CrossRef]
- Sakar, J.; Kazy, S.K.; Gupta, A.; Dutta, A.; Mohapatra, B.; Roy, A.; Bera, P.; Mitra, A.; Sar, P. Biostimulation of indigenous microbial community for bioremediation of petroleum refinery sludge. Front. Microbiol. 2016, 7, 1407. [Google Scholar]
- Tchobanoglous, G.; Burton, F.L.; Stensel, H.D. Wastewater Engineering: Treatment and Reuse, 4th ed.; McGraw-Hill Education: New York, NY, USA, 2003. [Google Scholar]
- Dos Santos, J.B.C.; da Silva Cruz, R.G.; Tardioli, P.W. Production of whole cell lipase from Streptomyces clavuligerus in a bench scale bioreactor and its first evaluation as biocatalyst for synthesis in organic medium. Appl. Biochem. Biotech. 2007, 183, 218–240. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-H.; Jin, Y.; Cheng, J.; Park, D.-J.; Kim, C.-J.; Hozzein, W.N.; Wadaan, M.A.M.; Shu, W.-S.; Ding, L.-X.; Li, W.-J. Gordonia jinhuaensis sp. nov., a novel actinobacterium, isolated from a VBNC (viable but non-culturable) state in pharmaceutical wastewater. Antonie Leeuwenhoek 2014, 106, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Abramić, M.; Lescic, I.; Konica, T.; Vitale, L.; Saenger, W.; Pigac, J. Purification and properties of extracellular lipase from Streptomyces rimosus. Enzym. Microb. Technol. 1999, 25, 522–529. [Google Scholar] [CrossRef]
- Bielen, A.; ćetković, H.; Long, P.F.; Schwab, H.; Abramić, M.; Vujaklija, D. The SGNH-hydrolase of Streptomyces coelicolor has (aryl)esterase and a true lipase activity. Biochimie 2009, 91, 390–400. [Google Scholar] [CrossRef]
- Ayaz, B.; Ugur, A.; Boran, R. Purification and characterization of organic solvent-tolerant lipase from Streptomyces sp. OC119-7 for biodiesel production. Biocatal. Agric. Biotechnol. 2015, 4, 103–108. [Google Scholar] [CrossRef]
- Seipke, R.F.; Crossman, L.; Drou, N.; Heavens, D.; Bibb, M.J.; Caccamo, M.; Hutchings, M.I. Draft genome sequence of Streptomyces strain S4, a symbiont of the leaf-cutting ant Acromyrmex octospinosus. J. Bacteriol. 2011, 193, 4270–4271. [Google Scholar] [CrossRef] [Green Version]
- Almeida, E.L.; Margassery, L.M.; Kennedy, J.; Dobson, A.D.W. Draft genome sequence of the antimycin-producing bacterium Streptomyces sp. strain SM8, isolated from the marine sponge Haliclona simulans. Genome Announc. 2018, 6, e01535-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.A.; Borchert, E.; O’Gara, F.; Dobson, A.D.W. Metagenomics for the discovery of novel biosurfactants of environmental interest from marine ecosystems. Curr. Opin. Biotechnol. 2015, 33, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Le Roes-Hill, M.; Durrell, K.A.; Kuegler, J. Biotechnological potential of actinobacterial biosurfactants: What we know and future perspectives. In Microbial Biosurfactants and Their Environmental and Industrial Applications, 1st ed.; Banat, I.M., Thavasi, R., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 174–208. [Google Scholar]
- Welz, P.J.; Ramond, J.-B.; Braun, L.; Vikram, S.; Le Roes-Hill, M. Biological nitrogen fixation in sand bioreactors treating winery wastewater with a high carbon to nitrogen ratio. J. Environ. Manag. 2018, 207, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Boran, R.; Ugur, A.; Sarac, N.; Ceylan, O. Characterisation of Streptomyces violascens OC125-8 lipase for oily wastewater treatment. 3Biotech 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Côté, A.; Shareck, F. Cloning, purification and characterisation of two lipases from Streptomyces coelicolor A3(2). Enzym. Microb. Technol. 2008, 42, 381–388. [Google Scholar] [CrossRef]
- Bhushan, I.; Yadav, A.K.; Parshad, R. Enhancement in the production of lipase from Arthrobacter sp. using fed-batch fermentation strategy. Asiat. J. Biotechnol. Resour. 2011, 2, 522–534. [Google Scholar]
- Sharma, A.; Bardhan, D.; Patel, R. Optimization of physical parameters for lipase production from Arthrobacter sp. BGCC#490. Indian J. Biochem. Biophys. 2009, 46, 178–183. [Google Scholar]
- Trujillo, M.E.; Kroppenstedt, R.M.; Fernández-Molinero, C.; Schumann, P.; Martínez-Molino, E. Micromonospora lupini sp. nov. and Micromonospora saelicesensis sp. nov., isolated from root nodules of Lupinus angustifolius. Int. J. Syst. Evol. Microbiol. 2007, 57, 2799–2804. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, F.R.C.; Cammarota, M.C.; Freire, D.M.G. The combined use of a biosurfactant and an enzyme preparation to treat an effluent with a high fat content. Colloid Surf. B 2012, 95, 241–246. [Google Scholar] [CrossRef]
- Louhasakul, Y.; Cheirsilp, B.; Intasit, R.; Maneerat, S.; Saimmai, A. Enhanced valorization of industrial wastes for biodiesel feedstocks and biocatalyst by lipolytic oleaginous yeast and biosurfactant-producing bacteria. Int. Biodeter Biodegrad. 2020, 148, 104911. [Google Scholar] [CrossRef]
- O’Neill, A.M.; Gallo, R.L. Host-microbiome interactions and recent progress into understanding the biology of acne vulgaris. Microbiome 2018, 6, 177. [Google Scholar] [CrossRef]
- Castillo, D.E.; Nanda, S.; Keri, J.E. Propionibacterium (Cutibacterium) acnes bacteriophage therapy in acne: Current evidence and future perspectives. Dermatol. Ther. 2019, 9, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollerup, S.; Friis-Nielsen, J.; Vinner, L.; Hansen, T.A.; Richter, S.R.; Fridholm, H.; Herrera, J.A.R.; Lund, O.; Brunak, S.; Izarzugaza, J.M.G.; et al. Propionibacterium acnes: Disease-causing agent or common contaminant? Detection in diverse patient samples by next-generation sequencing. J. Clin. Microbiol. 2016, 54, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Campisano, A.; Ometto, L.; Compant, S.; Pancher, M.; Antonielle, L.; Yousaf, S.; Varotto, C.; Anfora, G.; Pertot, I.; Sessitsch, A.; et al. Interkingdom transfer of the acne-causing agent, Propionibacterium acnes, from human to grapevine. Mol. Biol. Evol. 2014, 31, 1059–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousaf, S.; Bulgari, D.; Bergna, A.; Pancher, M.; Quaglino, F.; Casati, P.; Campisano, A. Pyrosequencing detects human and animal pathogenic taxa in the grapevine endosphere. Front. Microbiol. 2014, 5, 327. [Google Scholar] [CrossRef] [PubMed]
- McDowell, A.; Barnard, E.; Liu, J.; Li, H.; Patrick, S. Proposal to reclassify Propionibacterium acnes type I as Propionibacterium acnes subsp.acnes subsp. nov. and Propionibacterium acnes type II as Propionibacterium acnes subsp. defendens subsp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5358–5365. [Google Scholar]
- Dekio, I.; McDowell, A.; Sakamoto, M.; Tomida, S.; Ohkuma, M. Proposal of new combination, Cutibacterium acnes subsp. elongatum comb. nov., and emended descriptions of the genus Cutibacterium, Cutibacterium acnes subsp. acnes and Cutibacterium acnes subsp. defendens. Int. J. Syst. Evol. Microbiol. 2019, 69, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Ertuğrul, S.; Dönmez, G.; Takaç, S. Isolation of lipase producing Bacillus sp. from olive mill wastewater and improving its enzyme activity. J. Hazard Mater. 2007, 149, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Oliveira, F.; Dantas, D.; Gonçalves, C.; Belo, I. Lipase production by Aspergillus ibericus using olive mill wastewater. Bioprocess Biosyst. Eng. 2012, 36, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, V.; Fonseca, C.; Lopes da Silva, T.; Roseiro, J.C.; Eusébio, A. Isolation and identification of Magnusiomyces capitatus as a lipase-producing yeast from olive mill wastewater. Waste Biomass Valorization 2020, 11, 3207–3221. [Google Scholar] [CrossRef] [Green Version]
- Glogauer, A.; Martini, V.P.; Faoro, H.; Couto, G.H.; Müller-Santos, M.; Monteiro, R.A.; Mitchell, D.A.; de Souza, E.M.; Pedrosa, F.O.; Krieger, N. Identification and characterization of a new true lipase isolated through metagenomic approach. Microb. Cell Factories 2011, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HE | AW | RC | BT | ST | |
|---|---|---|---|---|---|
| COD screen (mg L−1) | 45 | ND * | 8400 | 33,200 | 381,000 |
| COD final (mg L−1) | ND | ND | <10 | 86,700 | 19,200 |
| Total N final (mg L−1) | ND | ND | BDL | 4 | 3 |
| Total P final (mg L−1) | ND | ND | BDL | 1290 | 1101 |
| COD:N (ratio) | ND | ND | ND | 21,625:1 | 6400:1 |
| COD:P (ratio) | ND | ND | ND | 67:1 | 17:1 |
| FOG screen (mg L−1) | ND | 110 | 1505 | 41,738 | 264,150 |
| Total fat (%wt.wt) | ND | ND | 0.21 | 1.56 | 0.23 |
| Strain | Nearest Phylogenetic Neighbour (% Sequence Similarity) | TDA | EYA | TBA | OOR | DC | OS (Zone) | E24 |
|---|---|---|---|---|---|---|---|---|
| BT1 | Arthrobacter ruber MDB1-42 (99.12%) | + | + | + | − | + CMO * | + ** (0.5 cm) | − |
| RC1 | Streptomyces werraensis NBRC 13404 (99.50%) | + | + | + | − | + CMO | − | 68% |
| BT2 | Micromonospora fluminis A38 (99.27%) | + | + | + | − | + M | − | − |
| BT3 | Streptomyces albidoflavus DSM 40455 (99.77%) | + | + | + | + | + CM * | − | − |
| BT4 | Streptomyces albidoflavus DSM 40455 (99.64%) | + | + | + | + | + CM * | − | − |
| RC2 | Streptomyces pratensis ch24 (99.49%) | + | + | + | − | + CMO * | − | 62% ** |
| BT5 | Streptomyces setonii NRRL ISP-5322 (99.78%) | + | + | + | + | + M | − | 88% |
| BT6 | Streptomyces colonosanans MUSC 93J (98.00%) | − | + | + | − | + M | − | 60% ** |
| BT7 | Gordonia jinhuaensis ZYR 51 (99.49%) | + | + | + | + | + CMO * | + ** (1 cm) | 60% ** |
| BT8 | Streptomyces griseoincarnatus LMG 19316 (98.28%) | + | − | + | − | + CM * | − | − |
| BT9 | Gordonia jinhauensis ZYR 51 (98.99%) | + | − | + | + | + CMO * | + ** (4 cm) | 60% ** |
| Type of Fat (% of Total Fat in Sample) | Total Fat (%, w/w of Sample) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Saturated | Unsaturated | Poly-Unsaturated | ||||||||||
| Avocado | 20.8 | 68.6 | 10.6 | 83.98 (0.84) * | ||||||||
| Olive | 17.6 | 78.1 | 4.3 | 81.37 (0.81) * | ||||||||
| Peanut | 20.8 | 42.7 | 36.5 | 71.44 (0.71) * | ||||||||
| Sunflower | 13.3 | 33.1 | 53.7 | 73.35 (0.73) * | ||||||||
| Canola | 20.7 | 62.1 | 17.2 | 78.44 (0.78) * | ||||||||
| RC | 28.6 | 47.6 | 23.8 | 0.21 | ||||||||
| BT | 23.7 | 57.1 | 19.2 | 1.56 | ||||||||
| ST | 30.4 | 47.8 | 21.7 | 0.23 | ||||||||
| Fatty acid type (%, w/w of sample) | ||||||||||||
| Palmitic | Stearic | Arachidic | Palmitoleic | Oleic | Linoleic | |||||||
| Avocado | 15.96 | 0.80 | 0.73 | 4.45 | 53.12 | 8.92 | ||||||
| Olive | 10.90 | 3.45 | 0 | 0.76 | 62.75 | 3.51 | ||||||
| Peanut | 11.33 | 3.56 | 0 | 0 | 30.49 | 26.06 | ||||||
| Sunflower | 6.07 | 3.67 | 0 | 0 | 24.25 | 39.36 | ||||||
| Canola | 4.81 | 2.20 | 9.26 | 0 | 48.69 | 13.48 | ||||||
| BT1 | BT2 | RT1 | RT2 | |
|---|---|---|---|---|
| Parameter | ||||
| Total valid reads | 57,271 | 25,339 | 1711 | 1035 |
| Number of reads identified to species level | 30,640 | 17,544 | 1484 | 898 |
| Number of species found (97% cut-off) | 130 | 97 | 21 | 20 |
| Average read lengths (bp) | 237.6 | 236.8 | 228.6 | 229.8 |
| α-diversity | ||||
| Shannon | 2.246 | 2.049 | 1.322 | 1.226 |
| Jackknife | 410 | 258 | 35.0 | 32.0 |
| Chao1 | 373.8 | 236.3 | 32.0 | 29 |
| Simpson | 0.247 | 0.342 | 0.507 | 0.547 |
| Number of OTUs found in sample | 363 | 226 | 29 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Welz, P.; Swanepoel, G.; Weels, S.; Le Roes-Hill, M. Wastewater from the Edible Oil Industry as a Potential Source of Lipase- and Surfactant-Producing Actinobacteria. Microorganisms 2021, 9, 1987. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091987
Welz P, Swanepoel G, Weels S, Le Roes-Hill M. Wastewater from the Edible Oil Industry as a Potential Source of Lipase- and Surfactant-Producing Actinobacteria. Microorganisms. 2021; 9(9):1987. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091987
Chicago/Turabian StyleWelz, Pamela, Gustav Swanepoel, Shandré Weels, and Marilize Le Roes-Hill. 2021. "Wastewater from the Edible Oil Industry as a Potential Source of Lipase- and Surfactant-Producing Actinobacteria" Microorganisms 9, no. 9: 1987. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9091987