
Diversity of the Tryptophanase Gene and Its Evolutionary Implications in Living Organisms


School of Chemical Engineering, Yeungnam University, 280 Daehak-Ro, Gyeongsan 38541, Korea
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(10), 2156; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102156
Submission received: 31 August 2021
/
Revised: 9 October 2021
/
Accepted: 12 October 2021
/
Published: 15 October 2021
(This article belongs to the Section Systems Microbiology)
Abstract
:Tryptophanase encoded by the gene tnaA is a pyridoxal phosphate-dependent enzyme that catalyses the conversion of tryptophan to indole, which is commonly used as an intra- and interspecies signalling molecule, particularly by microbes. However, the production of indole is rare in eukaryotic organisms. A nucleotide and protein database search revealed tnaA is commonly reported in various Gram-negative bacteria, but that only a few Gram-positive bacteria and archaea possess the gene. The presence of tnaA in eukaryotes, particularly protozoans and marine organisms, demonstrates the importance of this gene in the animal kingdom. Here, we document the distribution of tnaA and its acquisition and expansion among different taxonomic groups, many of which are usually categorized as non-indole producers. This study provides an opportunity to understand the intriguing role played by tnaA, and its distribution among various types of organisms.
1. Introduction
Tryptophanase (TnaA) is a pyridoxal 5′phosphate-dependent enzyme that catalyses the hydrolytic β-elimination of tryptophan to indole, pyruvate, and ammonia, which all play unique roles within organisms and the environment [1,2,3]. Consequently, it is an important enzyme from the perspective of amino acid and nitrogen metabolism. Pyruvate is a key molecule that links carbon and nitrogen metabolic pathways, drives metabolic flux according to environmental conditions and helps to regenerate reducing equivalents under aerobic and anaerobic environments [1]. Re-routing of pyruvate toward gluconeogenesis is associated with the maintenance of enterohaemorrhagic Escherichia coli O157:H7 in bovine intestines [1]. The degradation of tryptophan, like other amino acids, results in the production of ammonia as a by-product, which in turn may participate in other biochemical reactions such as nitrogen cycle post excretion into the environment.
Indole is a unique compound with wide-ranging effects on many organisms [4,5] and its involvement as an effector molecule in prokaryotes and eukaryotes is intriguing. In prokaryotes, it serves as a signalling molecule that modulates cell division, virulence, and biofilm formation [5]. Antibiotic tolerance is a major menace in clinical medicine [6,7] and indole contributes to the resistance acting as a signalling molecule [5]. In addition, indole-producing bacteria can interfere with quorum sensing, biofilm formation, antibiotic tolerance, pigment production, and predation abilities of non-indole producing bacteria including pathogens [2,3,4,5,8,9].
In eukaryotes, indole and indole-derived compounds display a broader significance. In insects, indole is a cue to identify food, mating partners, and ovipositional sites [10,11,12,13]. Indole also participates in survival dynamics and is utilized in predator–prey interactions and plant defence mechanisms [14,15,16]. Fungi and several marine organisms can use indole to produce indole-derived compounds, such as pigments, alkaloids, Tyrian purple, and antifouling agents [17,18,19]. In vertebrates, including humans, tryptophan metabolism leads to the synthesis of indole moieties containing precursors of key mediators such as serotonin, melatonin, kynurenine, and tryptamine [20,21,22].
Thus, available evidence demonstrates that tryptophan metabolism plays vital roles in various living forms [23]. The ability of TnaA to conduct reverse α,β-elimination and β-substitution reactions has also attracted attention because the former can be used to synthesize tryptophan [24]. Furthermore, the ability of TnaA to synthesize tryptophan is important in higher eukaryotic hosts because it is used by gut microbiota to produce the rare amino acid tryptophan [1]. Therefore, we decided to study the lineage and diversity of the tnaA gene to identify the roles of indole and TnaA in organisms and biotechnological applications associated with variations in the active sites of TnaA, especially concerning the biosynthesis of hormones and alkaloids.
2. Materials and Methods
2.1. Sequence Data/Data Mining
The tnaA sequences (>1200 nts) of 221 isolates of 36 eukaryotes (Drosophila, hemichordates and marine organisms including octopus, sea anemone, corals, hermit crab), 41 archaea, 16 fungi, and 128 prokaryotes (Gram-positive and -negative bacteria) were investigated in the present study. These gene sequences were obtained from the NCBI database (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene accessed on 8 October 2020). The following exclusion terms were used: putative protein; TnaA leader peptide (tnaC); tryptophan 2,3-dioxygenase; and hypothetical proteins; record removed; and large eukaryote sequences (with long ‘N’ stretches). Curation of tnaA sequences was performed manually to avoid redundant/partial or non-related sequences.
2.2. Phylogenetic Analyses
For the nucleotide sequence phylogenetic tree, the 221 tnaA nucleotide sequences were assembled and translated to their respective cDNA sequences and aligned using the multiple sequence alignment program ClustalX version 2.0.12. To estimate evolutionary distances, pairwise distances between species were calculated using the MEGA 11 package. A total of 221 tnaA ortholog sequences were analysed. Sequences were aligned with MEGA 11 and alignment errors were corrected using BioEdit 7.2. Evolutionary history was inferred using the Maximum Likelihood method and General Time Reversible model. Initial trees for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (4 categories (+G, parameter = 1.0799)). The rate variation model allowed for some sites to be evolutionarily invariable. The analysis was conducted for 1000 bootstrap replications. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analysis involved 221 nucleotide sequences. There was a total of 32,115 positions in the final dataset. Evolutionary analyses were conducted in MEGA11.
For the amino acid sequence phylogenetic tree, the nucleotide sequences were translated using MEGA 11, assembled and aligned using the multiple sequence alignment program ClustalX version 2.0.12 and alignment errors were corrected using BioEdit 7.2. The evolutionary history was inferred by using the Maximum Likelihood method and Le_Gascuel model [25]. Initial trees for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the JTT model, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (4 categories (+G, parameter = 1.5010)). The rate variation model allowed for some sites to be evolutionarily invariable. There was a total of 8750 positions in the final dataset. Evolutionary analyses were conducted in MEGA 11 software [26,27].
2.3. %GC Content
The %GC content of all the sequences were calculated using Oligo calculator—an online tool of University of California Berkeley accessed at the URL http://mcb.berkeley.edu/labs/krantz/tools/oligocalc.html accessed on 18 November 2020.
3. Results
3.1. Overall Patterns of Horizontal Gene Transfer (HGT)
We analysed 221 tnaA gene sequences of all organisms retrieved from the NCBI database. As the presence of tnaA confers unique abilities to the host (particularly bacteria), we addressed the enduring question regarding the distribution and availability of tnaA across known taxonomic groups. Eighty-five prokaryote species have been reported to possess tnaA and/or its activity [5], and indole was considered to be synthesized exclusively by bacteria. Using a phylogenetic approach, we assessed the evolutionary patterns of tnaA. A total of 190 species, that is, 117 bacteria, 36 archaea, 16 fungi, and 21 eukaryotic species harboured the tnaA gene (Table 1). The nucleotide based trees were constructed to elucidate the diversity of the tnaA gene in the selected groups wherein the tnaA gene is susceptible to point mutations and can differentiate gene diversity in closely related organisms. The phylogenetic tree results were further corroborated using %GC content as a parametric method. Due to high variability in GC content of the prokaryotic genomes, internal factors such as GC parametric comparison can help infer horizontal gene transfers [28,29].
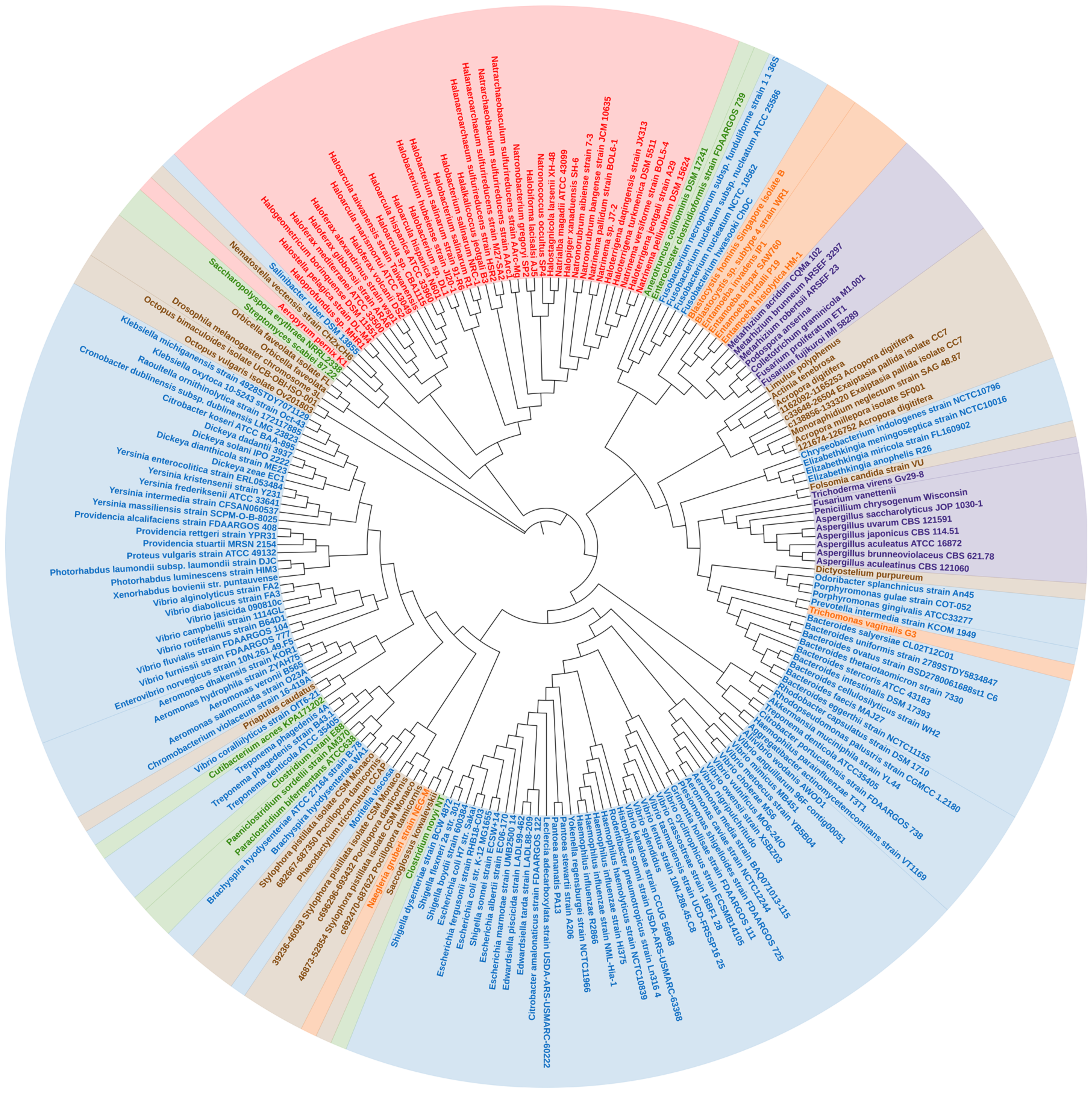
In a phylogenetic reconstruction using a dataset containing the 221 tnaA nucleotides and their respective amino acid orthologs, γ-proteobacteria tnaA formed a statistically well-supported branch in both nucleotide and amino acid trees of Enterobacteriales and Vibrionales. Thus, it appears the order ‘Enterobacterales’, γ–proteobacteria have remained conserved in terms of tnaA gene sharing. We noticed several cases of the “non-vertical” appearance of tnaA in distantly related organisms including fungi and other unicellular eukaryotes (Figure 1). Phylogenetic analysis revealed that HGT of tnaA in eukaryotes was not the result of contamination. A wider distribution of tnaA was found in prokaryotes, particularly γ-proteobacteria. The diversity of tnaA followed phylogenetic relationships already established using unique features of rRNA sequences, according to which archaea, bacteria, and eukaryotes are segregated in isolated clades. However, tnaA based phylogeny revealed some random appearances of closely related gene sequences belonging to taxonomically diverse groups revealing HGTs (Figure 1).
The close sequence similarity of >73% for 99% sequence coverage and similar %GC content (Table S1) between Yokenella and Rodentibacter tnaA indicates a possibility of HGT between two distantly related organisms (Figure 1 and Figure 2), which belong to recently diverged sister orders Enterobacteriales and Pasteurellales of the class γ–proteobacteria, respectively. Previously, glg genes were found to undergo complex gene transfers among γ–proteobacteria and other main bacterial groups [30]. Yokenella regensburgei possess a tnaA gene sequence, which contrasts with reports that the organism is negative for indole production [31,32]. This apparent contradiction may be due to the presence of non-functional tnaA, and therefore needs more investigation. This is also observed in Shigella. sp strains which possess the tnaA gene but mutations in cluster 1 and cluster 3 of the tnaA operon consequently render them indole-negative [33]. On the other hand, Enterococcus faecalis and Lactobacillus reuteri produce indole (in vitro), which is indicative of tnaA activity (Table 1); however, the presence of tnaA within their genomes has yet to be determined. Recently, Lactobacillus sp. isolated from infant faeces were studied for their anti-obesity potential [34]. There is an inverse correlation between indole production and fat content which partially confirms functional presence of tryptophanase in Lactobacillus sp. [13].
3.2. TnaA Gene in Archaea
In archaea, the topology of tnaA phylogeny suggests a distinct but common bacterial ancestor (Figure 1 and Figure 2). Few thermophiles thrive within biofilms by secreting extracellular polymeric substances [35], and few halophiles that produce acyl-homoserine lactone (a quorum-sensing signal molecule) secrete extracellular polymeric substances and form biofilms (Table 2). Relationships among genera remained largely undisturbed. Sequences belonging to archaea—‘Halobacteriales, Haloferacales and Natrialbales’ belonging to the class Haloarchaea and other species formed a distinct clade, while the position of the thermophile ‘Aeropyrum’ shifted slightly (Figure 1) with varying %GC content from the archaeal cluster (Table 3, (a)). The exact benefit of indole production by archaea has yet to be determined. Although archaeal biofilms are poorly studied and characterized, increasing evidence suggests that like prokaryotes, archaea benefit from living in biofilms because they afford protection against environmental stresses, fluctuating pH, and toxic chemicals [36,37,38]. Furthermore, polymicrobial biofilms provide environments that facilitate the transfer of genetic material and enable syntrophic associations [35]. The appearance of Aeropyrum pernix in different clades reveals a distinct yet unknown origin, as it is the only representative thermophile that possesses tnaA. The proximity of the Natrialbales clade with those of Halobacteriales and Haloferacales indicates a common ancestry (Figure 1 and Figure 2). Indeed, Haloterrigena spp., which was previously categorized as a member of the Halobacteriaceae family, shows a close relationship with Natrinema spp. and was proposed to be included in the family Natrialbaceae [39] and supported by similar %GC content (Table S1). Interestingly, haloarchaea retained tnaA and other archaea lost the gene. Salinibacter ruber was found near the archaeal cluster (Figure 1) and has similar %GC content (Table 3, (b)) which further supports a common ancestry with halophilic characteristics [40]. TnaA is one of the genes extensively exchanged among members of the Halobacteriaceae and Salinibacter families. Salinibacter ruber was found to be indole-negative which is a common characteristic in most Halobacteriaceae species (Table 1) which further bolsters the case for common ancestry. Further study is required to decipher the influence of indole on the halophilic lifestyles of microbes and the survival strategies of the indole-negative biofilm-producing archaeal microbes.
3.3. Significance of tnaA in the Eukaryotic Life Cycle
For HGT in cases where a wider distribution of genes is found in donor and recipient lineages (and in other taxa), phylogenetic trees are used to investigate transfer polarity [41]. When a gene is only found in donor and recipient groups/taxa, it is generally assumed that the source of gene transfer must be the taxon displaying the most diverse representation of that gene, since the possibility of gene transfer in several organisms from a single species at the same time is unlikely [42]. HGT is an essential evolutionary tool and was thought to be limited to prokaryotes, but over the past decade, increasing evidence indicates genetic materials are exchanged between prokaryotes and eukaryotes (both endosymbionts and free-living organisms) [43,44,45]. The absence of tnaA in the genomes of vertebrates, including mammals, indicates the need for tryptophan degradation has reduced and that the function is largely provided by gut microbes as a result of co-evolution in a holobiont [46]. However, vertebrates have also adopted other means of tryptophan degradation via the kynurenine pathway by cleaving the indole moiety with tryptophan 2,3-dioxygenase in the liver [47].
It has been reported that about 145 genes in man originated from bacteria, including a few involved in amino-acid metabolism acquired by HGT [46]. For instance, Maximum Likelihood and Bayesian phylogenetic methods showed that eukaryotic pyruvate formate lyase, a homolog of the firmicutes gene, may have been acquired through HGT [48]. Similarly, firmicutes (Anaerotruncus, Clostridium, Paraclostridium and Enterocloster) were found within the cluster accommodating fungi and/or unicellular eukaryotes in the nucleotide sequence tree (Figure 1 and Figure 2). TnaA appears to have transferred from bacteria into fungi by two independent HGT events as evidenced by the formation of a distinct clade by members of Sordariomycetes and Eurotiomycetes (Figure 1 and Figure 2) corroborated by their similar %GC content (Table 3, (c)). Sordariomycetes (a clade of fungi—Colletotrichum, Fusarium, Podospora, and Metarhizium) appeared as a sister group of a Blastocystis branch (with high statistical support—based on bootstrap values) (Figure 1 and Figure 2) and similar %GC content (Table 3, (d)). Other related fungi belonging to the Ascomycota group (Aspergillus, Penicillium, and Trichoderma) also had a similar origin as their tnaA sequences were also found to be associated with a clade containing a few protozoa Naegleria and Dictyostelium purpureum (Figure 1). The abundance of several bacterial tnaA genes indicates that the origin of tnaA could be interleaved with members of different bacterial phyla, and endosymbiosis of tnaA possessing bacteria might be the reason for their common ancestry [43]. E. histolytica tnaA might be a result of HGT from a relative of the anaerobic bacterium Fusobacterium (Figure 1 and Figure 2) seconded with similar %GC content (Table 3, (e)). In contrast, the T. vaginalis appears to acquire the gene from a separate origin by HGT from a relative of the anaerobic Bacteroides group (Figure 1 and Figure 2) and similar %GC content (Table 3, (f)). Only three species of anaerobic protists, Trichomonas vaginalis, Tritrichomonas foetus and E. histolytica have been reported to produce indole [49,50].
HGT might have played a role in the evolution and symbiotic adaptation of various unicellular organisms, including eukaryotes. Most of the unicellular organisms (>30%) we studied are pathogenic, and in these organisms, tnaA provides unique advantages for living within a host [20,23]. Previously, Pfam-based analysis led to the identification of 18 HGT events from bacteria to Dictyostelium, which possesses two β-eliminating lyases [51]. Thus, it is evident that the transfer of genetic material occurs between prokaryotes and unicellular eukaryotes, and that these eukaryotes gain a competitive advantage. Indeed, genes of prokaryotic origin are commonly transferred multiple times between protists [43].
HGT events can be classified into two broad types; those that maintain pre-existing functions (maintenance transfers) and those that add new functionality, such as host interaction, defence and adaptation to extreme environments, to the recipient (innovative transfers) [52]. Of note, many lineages (e.g., amoebae, ciliates, dinoflagellates, and non-parasitic excavates) lack reports of HGT events, probably because of sampling bias. Fungi, especially Ascomycota, are prolific producers of indole alkaloids, many of which display potent biological activities [53]. However, the function of indole and its metabolites are still largely undetermined in fungi. The NCBI-BLAST sequence similarities between Fusarium (along with Penicillium, Aspergillus, and Trichoderma) and Porphyromonas, Prevotella, and Trichomonas exceed 60% with e-values of <3 × 10−21 and similar %GC content (Table S1), indicating a possible HGT from a similar prokaryotic ancestor of the order ‘Bacteroidales’.
Pyruvate can be produced during amino acid metabolism and is specifically required for the regeneration of NAD+ during anaerobic fermentation, whereas aerobic fermentation increases ATP generation via the TCA cycle [1,54,55]. The inclusion of tnaA in a genome broadens metabolic ability especially concerning carbon (gluconeogenesis), amino acid, and nitrogen metabolism [1]. The decomposition and grazing activities of protozoans contribute toward the nitrogen cycle via ammonia generation. Pyruvate and ATP may also be produced via other metabolic pathways and thus the addition of tryptophan conversion may merely result in HGT being considered a ‘maintenance transfer’. Bacteria and protists with such alternative pathways harbouring tnaA thereby gain additional biochemical activity that provides survival and competitive advantages. Tryptophan may also serve as an energy source in S. salmonicida due to the presence of three copies of a bacteria-like TnaA that generate pyruvate, indole, and NH3 from tryptophan [56]. ‘Indole’ plays several other roles in the ecosystem and is considered an ‘archetypical hormone’ as it can regulate the behaviours of prokaryotes [8] and eukaryotes, including higher vertebrates and plants [23]. Therefore, the production of indole by tnaA may provide a means of manipulating a neighbour’s behaviour, which is of particular use in the gut where bacteria dominate, and other taxonomically diverse groups must compete for nutrients. Thus, the ability to produce indole provides distinct advantages, which suggests horizontal tnaA transfer should be considered an ‘innovative transfer’. The transfer of genetic material in this way is considered an indispensable driver of the evolution of fungi dwelling in the gut of higher organisms [57]. In addition, fungi belonging to the Ascomycota phylum produce indole alkaloids, which have high bioactivities, and octopuses and other marine organisms can use indole to synthesize pigments (e.g., Tyrian purple). The muricid mollusc Dicathais orbita produces Tyrian purple with the help of tissue dwelling Vibrio sp. [18]
Tryptophan is an essential amino acid produced by the shikimic acid pathway (by bacteria and plants, but not animals). Indole is an intermediate of this pathway and may serve as a precursor for tryptophan synthesis [58]. Furthermore, the ability of tnaA to work reversibly gives credence to the notion that bacteria can modulate its ability to produce tryptophan or degrade it to pyruvate and indole. Interestingly, fungi and endophytes interact symbiotically with host plants by synthesizing indole acetic acid (a plant growth hormone).

{kind=link}
{kind=link}
Table 1.
Organisms used in the study and their classification and indole production + positive; − negative; n/a—not available; v—varying.
Table 1.
Organisms used in the study and their classification and indole production + positive; − negative; n/a—not available; v—varying.
| Organism | Classification (Class; Order; and Family) | Indole Production | Reference | Organism | Classification (Class; Order; and Family) | Indole Production | Reference |
|---|---|---|---|---|---|---|---|
| Gram−negative bacteria | |||||||
| A. caviae | Gammaproteobacteria; Aeromonadales; Aeromonadaceae | + | [59] | M. viscosa | Gammaproteobacteria; Alteromonadales; Moritellaceae | − | [60] |
| A. dhakensis | + | [61] | O. splanchnicus | Bacteroidia; Bacteroidales; Odoribacteraceae | n/a | ||
| A. hydrophila | + | [59] | P. ananatis | Gammaproteobacteria; Enterobacterales; Erwiniaceae | + | [62] | |
| A. media | + | [59] | P. stewartia | + | [63] | ||
| A. salmonicida | + | [64] | P. laumondii | Gammaproteobacteria; Enterobacterales; Morganellaceae | − | [65] | |
| A. veronii | + | [59] | P. luminescens | − | [65] | ||
| A. actinomycetemcomitans | Gammaproteobacteria; Pasteurellales; Pasteurellaceae | n/a | P. shigelloides | Gammaproteobacteria; Enterobacterales; Enterobacterales incertae sedis | + | [66] | |
| A. muciniphila | Verrucomicrobiae; Verrucomicrobiales; Akkermansiaceae | n/a | P. gingivalis | Bacteroidia; Bacteroidales; Porphyromonadaceae | + | [67] | |
| A. wodanis | Gammaproteobacteria; Vibrionales; Vibrionaceae | + | [68] | P. gulae | + | [69] | |
| B. cellulosilyticus | Bacteroidia; Bacteroidales; Bacteroidaceae | n/a | P. intermedia | Bacteroidia; Bacteroidales; Prevotellaceae | + | [70] | |
| B. eggerthii | + | [71] | P. vulgaris | Gammaproteobacteria; Enterobacterales; Morganellaceae | + | [72] | |
| B. faecis | + | [73] | P. alcalifaciens | Gammaproteobacteria; Enterobacterales; Morganellaceae | + | [74] | |
| B. intestinalis | + | [75] | P. rettgeri | + | [76] | ||
| B. ovatus | + | [73] | P. stuartii | + | [76] | ||
| B. salyersiae | + | [73] | R. ornithinolytica | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | + | [77] | |
| B. stercoris | + | [78] | R. capsulatus | Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae | n/a | ||
| B. thetaiotaomicron | + | [73] | R. palustris | Alphaproteobacteria; Rhizobiales; Bradyrhizobiaceae | n/a | ||
| B. uniformis | + | [71] | R. pneumotropicus | Gammaproteobacteria; Pasteurellales; Pasteurellaceae | + | [79] | |
| B. hyodysenteriae | Spirochaetia; Brachyspirales; Brachyspiraceae | + | [80] | S. ruber | Bacteroidetes Order II. Incertae sedis; Rhodothermaceae | − | [81] |
| C. violaceum | Betaproteobacteria; Neisseriales; Chromobacteriaceae | + | [82] | S. boydii | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | − | [83] |
| C. indologenes | Flavobacteriia; Flavobacteriales; Weeksellaceae | − | [84] | S. dysenteriae | − | [33] | |
| C. amalonaticus | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | n/a | S. flexneri | − | [33] | ||
| C. koseri | + | [85] | S. sonnei | − | [33] | ||
| C. portucalensis | − | [86] | V. alginolyticus | Gammaproteobacteria; Vibrionales; Vibrionaceae | + | [68] | |
| C.dublinensis | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | + | [87] | V. anguillarum | v | [68] | |
| D. dadantii | Gammaproteobacteria; Enterobacterales; Pectobacteriaceae | + | [88] | V. campbellii | + | [68] | |
| D. dianthicola | + | [89] | V. cholerae | + | [90] | ||
| D. solani | + | [91] | V. coralliilyticus | + | [92] | ||
| D. zeae | + | [89] | V. crassostreae | + | [93] | ||
| E. piscicida | Gammaproteobacteria; Enterobacterales; Hafniaceae | + | [94] | V. cyclitrophicus | − | [68] | |
| E. tarda | + | [94] | V. diabolicus | + | [95] | ||
| E. anophelis | Flavobacteriia; Flavobacteriales; Weeksellaceae | + | [96] | V. fluvialis | + | [68] | |
| E. meningoseptica | + | [97] | V. furnissii | v | [98] | ||
| E. miricola | + | [97] | V. jasicida | + | [99] | ||
| E. norvegicus | Gammaproteobacteria; Vibrionales; Vibrionaceae | + | [100] | V. kanaloae | + | [93] | |
| E. albertii | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | + | [101] | V. lentus | + | [68] | |
| E. coli | + | [102] | V. metoecus | + | [103] | ||
| E. fergusonii | + | [104] | V. mimicus | + | [68] | ||
| E. marmotae | − | [105] | V. nigripulchritudo | + | [68] | ||
| F. hwasookii | Fusobacteria; Fusobacteriales; Fusobacteriaceae | + | [106] | V. owensii | + | [99] | |
| F. necrophorum | + | [107] | V. rotiferianus | + | [99] | ||
| F. nucleatum | + | [106] | V. splendidus | + | [68] | ||
| F. nucleatum | + | [106] | V. tasmaniensis | + | [68] | ||
| G. hollisae | Gammaproteobacteria; Vibrionales; Vibrionaceae | + | [108] | V. vulnificus | + | [68] | |
| H. haemolyticus | Gammaproteobacteria; Pasteurellales; Pasteurellaceae | + | [109] | X. bovienii | Gammaproteobacteria; Enterobacterales; Morganellaceae | n/a | |
| H. influenzae | + | [109] | Y. enterocolitica | Gammaproteobacteria; Enterobacterales; Yersiniaceae | + | [110] | |
| H. parainfluenzae | + | [111] | Y. frederiksenii | + | [112] | ||
| H. somni | Gammaproteobacteria; Pasteurellales; Pasteurellaceae | + | [113] | Y. intermedia | + | [114] | |
| K. michiganensis | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | + | [115] | Y. kristensenii | v | [116] | |
| K. oxytoca | + | [117] | Y. massiliensis | + | [118] | ||
| Ladecarboxylata | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | + | [119] | Y. regensburgei | Gammaproteobacteria; Enterobacterales; Enterobacteriaceae | − | [31] |
| Gram−positive bacteria | |||||||
| A. colihominis | Clostridia; Clostridiales; Ruminococcaceae | + | [120] | P. bifermentans | Clostridia; Clostridiales; Peptostreptococcaceae | + | [121] |
| C. novyi | Clostridia; Clostridiales; Clostridiaceae | + | [122] | S. erythraea | Actinobacteria; Pseudonocardiales; Pseudonocardiaceae | n/a | |
| C. tetani | + | [123] | S. scabiei | Actinobacteria; Streptomycetales; Streptomycetaceae | n/a | ||
| C. acnes | Actinobacteria; Propionibacteriales; Propionibacteriaceae | − | [124] | T. denticola | Spirochaetia; Spirochaetales; Spirochaetaceae | + | [125] |
| E. clostridioformis | Clostridia; Clostridiales; Lachnospiraceae | n/a | T. phagedenis | + | [126] | ||
| P. sordellii | Clostridia; Clostridiales; Peptostreptococcaceae | n/a | |||||
| Archaea | |||||||
| A. pernix | Thermoprotei; Desulfurococcales; Desulfurococcaceae | n/a | H. larsenii | Halobacteria; Halobacteriales; Halobacteriaceae | n/a | ||
| H. jeotgali | Halobacteria; Halobacteriales; Halobacteriaceae | v | [127] | H. pelagica | Halobacteria; Halobacteriales; Halobacteriaceae | + | [128] |
| H. sulfurireducens | Halobacteria; Halobacteriales; Halobacteriaceae | n/a | H. daqingensis | Halobacteria; Natrialbales; Natrialbaceae | − | [129] | |
| H. hispanica | Halobacteria; Halobacteriales; Haloarculaceae | v | [130] | H. jeotgali | Halobacteria; Natrialbales; Natrialbaceae Halobacteria; Natrialbales; Natrialbaceae | + | [131] |
| H. marismortui | − | [132] | H. turkmenica | + | [133] | ||
| H. taiwanensis | − | [134] | Natrarchaeobaculum sulfurireducens | − | [135] | ||
| Haloarcula sp. | − | [134] | N. magadii | Halobacteria; Natrialbales; Natrialbaceae | n/a | ||
| H. hubeiense | Halobacteria; Halobacteriales; Halobacteriaceae | n/a | N. pallidum | Halobacteria; Natrialbales; Natrialbaceae | − | [136] | |
| H. salinarum | v | [137] | N. pellirubrum | Halobacteria; Natrialbales; Natrialbaceae Halobacteria; Natrialbales; Natrialbaceae | − | [136] | |
| Halobacterium sp. | n/a | N. versiforme | + | [138] | |||
| H. lacisalsi | Halobacteria; Natrialbales; Natrialbaceae | − | [139] | Natrinema sp. | v | ||
| H. alexandrinus | Halobacteria; Haloferacales; Haloferacaceae | + | [140] | N. gregoryi | n/a | ||
| H. gibbonsii | + | [130] | N. occultus | Halobacteria; Natrialbales; Natrialbaceae | n/a | ||
| H. mediterranei | + | [141] | N. aibiense | Halobacteria; Natrialbales; Natrialbaceae | + | [142] | |
| H. volcanii | + | [143] | N. bangense | + | [144] | ||
| H. borinquense | Halobacteria; Haloferacales; Haloferacaceae | + | [145] | ||||
| H. xanaduensis | Halobacteria; Halobacteriales; Halobacteriaceae | − | [146] | ||||
| Eukaryotes | |||||||
| Protists | |||||||
| A. aculeatinus | Eurotiomycetes; Eurotiales; Aspergillaceae | n/a | N. gruberi | Heterolobosea; Schizopyrenida; Vahlkampfiidae | n/a | ||
| A. aculeatus | n/a | T. vaginalis | --; Trichomonadida; Trichomonadidae | + | [49] | ||
| A. brunneoviolaceus | n/a | E. dispar | Amoebozoa; Mastigamoebida; Entamoebidae Amoebozoa; Mastigamoebida; Entamoebidae | n/a | |||
| A. japonicus | n/a | E. histolytica | + | [50] | |||
| A. saccharolyticus | n/a | E. invadens | n/a | ||||
| A. uvarum | n/a | E. nuttalli | n/a | ||||
| C. graminicola | Sordariomycetes; Glomerellales; Glomerellaceae | n/a | |||||
| F. fujikuroi | Sordariomycetes; Hypocreales; Nectriaceae | n/a | Higher eukaryotes | ||||
| F. proliferatum | n/a | O. bimaculoides | Cephalopoda; Octopoda; Octopodidae Cephalopoda; Octopoda; Octopodidae Anthozoa; Scleractinia; Acroporidae | n/a | |||
| F. vanettenii | n/a | O. vulgaris | n/a | ||||
| M. acridum | Sordariomycetes; Hypocreales; Clavicipitaceae | n/a | A. digitifera | Anthozoa; Scleractinia; Acroporidae Anthozoa; Scleractinia; Pocilloporidae Anthozoa; Scleractinia; Acroporidae Anthozoa; Scleractinia; Pocilloporidae | n/a | ||
| M. brunneum | n/a | A. millepora | n/a | ||||
| M. robertsii | n/a | S. pistillata | Anthozoa; Actiniaria; Aiptasiidae | n/a | |||
| P. chrysogenum | Eurotiomycetes; Eurotiales; Aspergillaceae | n/a | E. pallida | Anthozoa; Actiniaria; Aiptasiidae | n/a | ||
| P. anserina | Sordariomycetes; Sordariales; Chaetomiaceae | n/a | A. tenebrosa | Anthozoa; Actiniaria; Actiniidae | n/a | ||
| T. virens | Sordariomycetes; Hypocreales; Hypocreaceae | n/a | N. vectensis | Anthozoa; Actiniaria; Edwardsiidae | n/a | ||
| O. faveolata | Anthozoa; Scleractinia; Merulinidae | n/a | |||||
| M. neglectum | Chlorophyceae; Sphaeropleales; Selenastraceae | n/a | P. damicornis | Anthozoa; Scleractinia; Pocilloporidae | n/a | ||
| D. purpureum | Eumycetozoa; Dictyosteliales; Dictyosteliaceae | n/a | S. kowalevskii | Enteropneusta; --; Harrimaniidae [Hemichordata] | n/a | ||
| B. hominis, | Bigyra; Opalinata; Blastocystidae | n/a | L. polyphemus | Merostomata; Xiphosura; Limulidae | n/a | ||
| Blastocystis sp. | n/a | F. candida | Collembola; Entomobryomorpha; Isotomidae | n/a | |||
| P. tricornutum | Bacillariophyceae; Naviculales; Phaeodactylaceae | n/a | D. melanogaster | Insecta; Diptera; Drosophilidae | n/a | ||
| P. caudatus | Priapulimorpha; Priapulimorphida; Priapulidae | n/a | |||||
Table 2.
List of archaea possessing the tnaA gene and/or indole producing activity.
| Organism | Unique Growth Requirements | Indole | Biofilm (EPS or QS Signal) | References | |
|---|---|---|---|---|---|
| Gene (NCBI) | Production | ||||
| Aeropyrum pernix K1 | Thermophile, 90 °C | + | − | − | [147] |
| Halalkalicoccus jeotgali B3 | 1.70–5.1 M NaCl | + | + | − | [131] |
| Halanaeroarchaeum sulfurireducens strain HSR2 | 3–5 M NaCl | + | NR | − | [148] |
| Halanaeroarchaeum sulfurireducens strain M27-SA2 | 3–5 M NaCl | + | NR | − | [149] |
| Haloarcula hispanica ATCC 33960 | 1.70–5.1 M NaCl | + | + | EPS | [132] [150] |
| Haloarcula hispanica N601 | + | +/− | EPS | ||
| Haloarcula marismortui ATCC 43049 | + | − | EPS | ||
| Haloarcula taiwanensis strain Taiwanensis | + | NR | NR | ||
| Haloarcula sp. CBA1115 | + | ||||
| Halobacterium hubeiense strain JI20-1 | + | ||||
| Halobacterium salinarum NRC-1 | 3.5–5 M NaCl | + | NR | + | |
| Halobacterium salinarum R1 | 4.2 M NaCl | + | NR | EPS/AHL | |
| Halobiforma lacisalsi AJ5 | >1.7 M NaCl | + | − | − | |
| Haloferax alexandrinus strain wsp1 | 4.3 M NaCl | + | + | − | [140] |
| Haloferax gibbonsii strain ARA6 | 1.7–4.3 M NaCl | + | + | EPS | |
| Haloferax mediterranei ATCC 33500 | 1–5.2 M NaCl | + | + | EPS | |
| Haloferax volcanii DS2 | 2.6–4.3 M NaCl | + | NR | +/AHLs | |
| Halogeometricum borinquense DSM 11551 | 3.4–4.3 M NaCl | + | + | +/AHLs | |
| Halopiger xanaduensis SH-6 | 4.3 M NaCl | + | − | − | |
| Haloprofundus sp. MHR1 | 0.9–4.8 M NaCl | + | − | − | |
| Halostagnicola larsenii XH-48 | 2.5–5.0 M NaCl | + | − | − | |
| Halostella pelagica strain DL-M4 | 2.6 M NaCl | + | NA | − | |
| Haloterrigena daqingensis strain JX313 | 1.7–5.5 M NaCl | + | − | − | |
| Haloterrigena jeotgali strain A29 | 2.6–3.4 M NaCl | + | + | − | |
| Haloterrigena turkmenica DSM 5511 | 2.6–3.4 M NaCl | + | NA | NA | |
| Natrarchaeobaculum sulfurireducens strain AArc1 | 3–5 M NaCl | + | − | NA | |
| Natrialba magadii ATCC 43099 | 3.4 M NaCl | + | NR | NA | |
| Natrinema pallidum strain BOL6-1 | 3.4–4.3 M NaCl | + | − | NA | |
| Natrinema pellirubrum DSM 15624 | 3.4–4.3 M NaCl | + | − | NA | |
| Natrinema versiforme strain BOL5-4 | 3.4–4.3 M NaCl | + | + | NA | |
| Natronobacterium gregoryi SP2 | 2.0–5 M NaCl | + | NR | NA | |
| Natronococcus occultus SP4 | 2 M NaCl | + | NR | AHL/Biofilm | |
| Natronorubrum aibiense strain 7-3 | 2.0–4.3 M NaCl | + | + | NA | [142] |
| Natronorubrum bangense strain JCM 10635 | 2.0–4.3 M NaCl | + | + | NA | [144] |
NR: not reported; NA: not available; AHL: Acyl-homoserine lactone; EPS: extracellular polymeric substance. + positive; − negative.
Table 3.
The %GC content comparison of (a) Aeropyrum pernix K1 and archaeal cluster, (b) Salinibacter ruber and archaeal cluster, (c) Sadriomycetes and Euratiomycetes cluster, (d) Sadriomycetes and Blastocystis.sp branch, (e) Fusobacterium sp. branch and Entamoeba histolytica HM-1 and (f) Bacteroides.sp branch and Trichomonas vaginalis G3.
Table 3.
The %GC content comparison of (a) Aeropyrum pernix K1 and archaeal cluster, (b) Salinibacter ruber and archaeal cluster, (c) Sadriomycetes and Euratiomycetes cluster, (d) Sadriomycetes and Blastocystis.sp branch, (e) Fusobacterium sp. branch and Entamoeba histolytica HM-1 and (f) Bacteroides.sp branch and Trichomonas vaginalis G3.
| (a) | (b) | (c) | |||
| Organism | %GC content | Organism | %GC content | Organism | %GC content |
| N. occultus SP4 | 68 | N. occultus SP4 | 68 | A. aculeatinus CBS 121060 | 65 |
| N. aibiense strain 7-3 | 66 | N. aibiense strain 7-3 | 66 | A. aculeatus ATCC 16872 | 63 |
| N. gregoryi SP2 | 65 | N. gregoryi SP2 | 65 | A. brunneoviolaceus CBS 621.78 | 65 |
| N. pallidum strain BOL6-1 | 66 | N. pallidum strain BOL6-1 | 66 | A. japonicus CBS 114.51 | 62 |
| N. magadii ATCC 43099 | 66 | N. magadii ATCC 43099 | 66 | A. saccharolyticus JOP 1030-1 | 65 |
| N. sulfurireducens strain AArc1 | 67 | N. sulfurireducens strain AArc1 | 67 | A. uvarum CBS 121591 | 64 |
| H. borinquense DSM 11551 | 63 | H. borinquense DSM 11551 | 63 | P. anserina S mat+ | 56 |
| H. gibbonsii strain ARA6 | 68 | H. gibbonsii strain ARA6 | 68 | M. acridum CQMa 102 | 53 |
| H. jeotgali strain A29 | 68 | H. jeotgali strain A29 | 68 | M. brunneum ARSEF 3297 | 52 |
| H. pelagica strain DL-M4 | 67 | H. pelagica strain DL-M4 | 67 | M. robertsii ARSEF 23 | 53 |
| H. larsenii XH-48 | 65 | H. larsenii XH-48 | 65 | F. fujikuroi IMI 58289 | 50 |
| H. xanaduensis SH-6 | 69 | H. xanaduensis SH-6 | 69 | F. proliferatum ET1 | 50 |
| H. lacisalsi AJ5 | 69 | H. lacisalsi AJ5 | 69 | F. vanettenii | 58 |
| Halobacterium sp. DL1 | 71 | Halobacterium sp. DL1 | 71 | C. graminicola M1.001 | 56 |
| H. salinarum NRC-1 | 71 | H. salinarum NRC-1 | 71 | ||
| H. sulfurireducens strain HSR2 | 67 | H. sulfurireducens strain HSR2 | 67 | ||
| H. jeotgali B3 | 65 | H.jeotgali B3 | 65 | ||
| A. pernix K1 | 56 | S. ruber DSM 13855 | 67 | ||
| (d) | (e) | (f) | |||
| Organism | %GC content | Organism | %GC content | Organism | %GC content |
| P. anserina S mat+ | 56 | F. hwasookii ChDC | 34 | B. cellulosilyticus strain WH2 | 46 |
| M. acridum CQMa 102 | 53 | F. hwasookii ChDC F300 | 31 | B. eggerthii strain NCTC11155 | 45 |
| M. brunneum ARSEF 3297 | 52 | F. necrophorum strain 1_1_36S | 40 | B. faecis MAJ27 | 46 |
| M. robertsii ARSEF 23 | 53 | F. nucleatum ATCC 25586 | 33 | B. intestinalis DSM 17393 | 46 |
| F. fujikuroi IMI 58289 | 50 | F. nucleatum strain NCTC10562 | 33 | B. ovatus strainBSD2780061688st1_C6 | 47 |
| F. proliferatum ET1 | 50 | E. histolytica HM-1 | 33 | B. salyersiae CL02T12C01 | 46 |
| F. vanettenii | 58 | B. stercoris ATCC 43183 | 44 | ||
| C. graminicola M1.001 | 56 | B. thetaiotaomicron strain 7330 | 48 | ||
| B. hominis, Singapore isolate B | 56 | T. vaginalis G3 | 48 | ||
| Blastocystis sp. strain WR1 | 57 |
4. Conclusions
Among prokaryotes, we observed a wider distribution of tnaA in γ-proteobacteria, which share similar copies of the gene among themselves and therefore grouped. Our phylogenetic analysis suggests HGT has played a crucial role in tnaA gene transfer and provided ‘maintenance and innovative gain’ to recipients. These results are quite relevant from the host–parasite interaction perspective. Acquired tnaA has probably enabled unicellular eukaryotes to regulate the gut environment and live in synergy with microbiota and other enteric bacteria. We suggest that the adaptation of the intestinal protozoan Blastocystis sp. to the gut resulted in the lateral acquisition of the tnaA gene and enabled the supply of additional metabolites to the pre-existing metabolic systems.
Similarly, other unicellular eukaryotes such as Dictyostelium, Entamoeba, and Trichomonas may have inherited tnaA from a eukaryotic ancestor. However, the sparse presence of tnaA in higher eukaryotes (e.g., sea anemone, corals, octopuses, drosophila, and nematodes) is insufficient to reveal its origins. On the other hand, the common occurrence of tnaA in archaea belonging to the class Haloarchaea is intriguing and warrants further investigation of the role of indole in adaptation to halophilic environments. Collectively, these results reveal new avenues of research directed toward improved understanding of the roles of indole in various organisms including eukaryotes.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/microorganisms9102156/s1, Table S1. % GC content of all the species sequences. Figure S1. Nucleotide sequence phylogenetic tree (arbitrarily rooted maximum likelihood tree) of the tnaA gene representing a total of 221 nucleotide sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values >= 70 (1000 replications). Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown). Figure S2. Amino acid sequence phylogenetic tree (arbitrarily rooted maximum likelihood tree) of the tnaA gene representing a total of 221 amino acid sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values >= 70 (1000 replications). Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown).
Author Contributions
Conceptualization, P.K. and J.L.; methodology, P.K. and B.R.B.; software, P.K. and B.R.B.; validation, P.K. and B.R.B.; formal analysis P.K. and B.R.B.; investigation, P.K. and B.R.B.; resources, J.-H.L. and J.L.; data curation, P.K. and B.R.B.; writing—original draft preparation, P.K. and B.R.B.; writing—review and editing, P.K., B.R.B., J.-H.L. and J.L.; visualization, P.K. and B.R.B.; supervision, J.-H.L. and J.L.; project administration, J.L.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from the National Research Foundation of Korea (NRF) funded by the Korean government (MSIT) (2021R1A2C1008368) and by the Priority Research Center Program through the NRF funded by the Ministry of Education (2014R1A6A1031189).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article (and/or) its Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bertin, Y.; Deval, C.; de la Foye, A.; Masson, L.; Gannon, V.; Harel, J.; Martin, C.; Desvaux, M.; Forano, E. The gluconeogenesis pathway is involved in maintenance of enterohaemorrhagic Escherichia coli O157:H7 in bovine intestinal content. PLoS ONE 2014, 9, e98367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwidar, M.; Nam, D.; Mitchell, R.J. Indole negatively impacts predation by Bdellovibrio bacteriovorus and its release from the bdelloplast. Environ. Microbiol. 2015, 17, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Romano, B.; Gollihar, J.; Brown, S.A.; Whiteley, M.; Valenzuela, E.; Kaplan, H.B.; Wood, T.K.; McLean, R.J.C. Indole inhibition of N-acylated homoserine lactone-mediated quorum signalling is widespread in Gram-negative bacteria. Microbiology 2014, 160, 2464–2473. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.G.; Baek, K.H.; Cho, M.H.; Lee, J. The multifaceted roles of the interspecies signalling molecule indole in Agrobacterium tumefaciens. Environ. Microbiol. 2015, 17, 1234–1244. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, J. Indole as an intercellular signal in microbial communities. FEMS Microbiol. Rev. 2010, 34, 426–444. [Google Scholar] [CrossRef]
- Lee, C.-R.; Cho, I.H.; Jeong, B.C.; Lee, S.H. Strategies to minimize antibiotic resistance. Int. J. Environ. Res. Public Health 2013, 10, 4274–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.Y.; Kim, D.M.; Jo, Y.; Kim, J.M.; Yoo, S.M. DNA Microarray-based detection of bacteria in samples containing antibiotics: Effect of antibiotics on the performance of pathogen detection assays. Biotechnol. Bioprocess Eng. 2021, 26, 447–455. [Google Scholar] [CrossRef]
- Lee, J.; Jayaraman, A.; Wood, T.K. Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol. 2007, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Zhang, X.-S.; Hegde, M.; Bentley, W.E.; Jayaraman, A.; Wood, T.K. Indole cell signaling occurs primarily at low temperatures in Escherichia coli. ISME J. 2008, 2, 1007–1023. [Google Scholar] [CrossRef]
- Dekel, A.; Yakir, E.; Bohbot, J.D. The evolutionarily conserved indolergic receptors of the non-hematophagous elephant mosquito Toxorhynchites amboinensis. Insect Biochem. Mol. Biol. 2019, 110, 45–51. [Google Scholar] [CrossRef]
- Jürgens, A.; Dötterl, S.; Meve, U. The chemical nature of fetid floral odours in stapeliads (Apocynaceae-Asclepiadoideae-Ceropegieae). N. Phytol. 2006, 172, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Urech, R.; Green, P.E.; Rice, M.J.; Brown, G.W.; Duncalfe, F.; Webb, P. Composition of chemical attractants affects trap catches of the australian sheep blowfly, Lucilia cuprina, and Other Blowflies. J. Chem. Ecol. 2004, 30, 851–866. [Google Scholar] [CrossRef]
- Kumar, P.; Lee, J.-H.; Lee, J. Diverse roles of microbial indole compounds in eukaryotic systems. Biol. Rev. 2021. [Google Scholar] [CrossRef]
- Bogner, C.W.; Kamdem, R.S.; Sichtermann, G.; Matthaus, C.; Holscher, D.; Popp, J.; Proksch, P.; Grundler, F.M.; Schouten, A. Bioactive secondary metabolites with multiple activities from a fungal endophyte. Microb. Biotechnol. 2017, 10, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, C.; Li, X.; Tang, Y.; Wang, Y.; Chen, S.; Yan, S. RNA-Seq identification of candidate defense genes targeted by endophytic Bacillus cereus-mediated induced systemic resistance against Meloidogyne incognita in tomato. Pest. Manag. Sci. 2018, 74, 2793–2805. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.G.; Kim, M.; Kim, E.; Choi, H.; Kim, Y.; Lee, J. Indole-associated predator-prey interactions between the nematode Caenorhabditis elegans and bacteria. Environ. Microbiol. 2017, 19, 1776–1790. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhang, X.; Qu, Y. Biodegradation and biotransformation of indole: Advances and perspectives. Front. Microbiol. 2018, 9, 2625. [Google Scholar] [CrossRef] [PubMed]
- Ngangbam, A.K.; Baten, A.; Waters, D.L.E.; Whalan, S.; Benkendorff, K. Characterization of bacterial communities associated with the tyrian purple producing gland in a marine gastropod. PLoS ONE 2015, 10, e0140725. [Google Scholar] [CrossRef]
- Wang, X.; Huang, Y.; Sheng, Y.; Su, P.; Qiu, Y.; Ke, C.; Feng, D. Antifouling activity towards mussel by small-molecule compounds from a strain of Vibrio alginolyticus bacterium associated with sea anemone Haliplanella sp. J. Microbiol. Biotechnol. 2017, 27, 460–470. [Google Scholar] [CrossRef] [Green Version]
- Hendrikx, T.; Schnabl, B. Indoles: Metabolites produced by intestinal bacteria capable of controlling liver disease manifestation. J. Intern. Med. 2019, 286, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Huć, T.; Nowinski, A.; Drapala, A.; Konopelski, P.; Ufnal, M. Indole and indoxyl sulfate, gut bacteria metabolites of tryptophan, change arterial blood pressure via peripheral and central mechanisms in rats. Pharmacol. Res. 2018, 130, 172–179. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [Green Version]
- Costantini, C.; Bellet, M.M.; Renga, G.; Stincardini, C.; Borghi, M.; Pariano, M.; Cellini, B.; Keller, N.; Romani, L.; Zelante, T. Tryptophan co-metabolism at the host-pathogen interface. Front. Immunol. 2020, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuidate, T.; Tansila, N.; Chomchuen, P.; Phattaranit, P.; Eangchuan, S.; Vuddhakul, V. Characterization of tryptophanase from Vibrio cholerae. Appl. Biochem. Biotechnol. 2015, 175, 243–252. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jain, R.; Rivera, M.C.; Moore, J.E.; Lake, J.A. Horizontal gene transfer accelerates genome innovation and evolution. Mol. Biol. Evol. 2003, 20, 1598–1602. [Google Scholar] [CrossRef] [PubMed]
- Noon, J.; Baum, T. Horizontal gene transfer of acetyltransferases, invertases and chorismate mutases from different bacteria to diverse recipients. BMC Evol. Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro, G.; Viale, A.M.; Montero, M.; Rahimpour, M.; Munoz, F.J.; Baroja-Fernandez, E.; Bahaji, A.; Zuniga, M.; Gonzalez-Candelas, F.; Pozueta-Romero, J. Comparative genomic and phylogenetic analyses of Gammaproteobacterial glg genes traced the origin of the Escherichia coli glycogen glgBXCAP operon to the last common ancestor of the sister orders Enterobacteriales and Pasteurellales. PLoS ONE 2015, 10, e0115516. [Google Scholar] [CrossRef] [Green Version]
- Abbott, S.L.; Janda, J.M. Isolation of Yokenella regensburgei (“Koserella trabulsii”) from a patient with transient bacteremia and from a patient with a septic knee. J. Clin. Microbiol. 1994, 32, 2854–2855. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Gaind, R.; Gupta, K.B.; Dawar, R.; Kumar, D.; Paul, P.; Sardana, R.; Deb, M. Yokenella regensburgei infection in India mimicking enteric fever. J. Med. Microbiol. 2013, 62, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Rezwan, F.; Lan, R.; Reeves, P.R. Molecular basis of the indole-negative reaction in Shigella Strains: Extensive damages to the tna operon by insertion sequences. J. Bacteriol. 2004, 186, 7460–7465. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Jeong, Y.; Kim, J.-E.; Kim, Y.; Paek, N.-S.; Kang, C.-H. Anti-obesity potential of Lactobacillus spp. isolated from infant feces. Biotechnol. Bioprocess Eng. 2021, 26, 575–585. [Google Scholar] [CrossRef]
- Van Wolferen, M.; Orell, A.; Albers, S.-V. Archaeal biofilm formation. Nat. Rev. Microbiol. 2018, 16, 699–713. [Google Scholar] [CrossRef]
- Bang, C.; Ehlers, C.; Orell, A.; Prasse, D.; Spinner, M.; Gorb, S.N.; Albers, S.V.; Schmitz, R.A. Biofilm formation of mucosa-associated methanoarchaeal strains. Front. Microbiol. 2014, 5, 353. [Google Scholar] [CrossRef] [PubMed]
- Fröls, S.; Dyall-Smith, M.; Pfeifer, F. Biofilm formation by haloarchaea. Environ. Microbiol. 2012, 14, 3159–3174. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Morales, B.O.; Narváez-Zapata, J.A.; Schmalenberger, A.; Sosa-López, A.; Tebbe, C.C. Biofilms fouling ancient limestone Mayan monuments in Uxmal, Mexico: A cultivation-independent analysis. Biofilms 2004, 1, 79–90. [Google Scholar] [CrossRef]
- Gupta, R.S.; Naushad, S.; Baker, S. Phylogenomic analyses and molecular signatures for the class Halobacteria and its two major clades: A proposal for division of the class Halobacteria into an emended order Halobacteriales and two new orders, Haloferacales ord. nov. and Natrialbales ord. nov., containing the novel families Haloferacaceae fam. nov. and Natrialbaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1050–1069. [Google Scholar] [CrossRef]
- Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhao, J.; Tang, N.; Sun, H.; Huang, J. Horizontal gene transfer from bacteria and plants to the arbuscular mycorrhizal fungus Rhizophagus irregularis. Front. Plant Sci. 2018, 9, 701. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal gene transfer in prokaryotes: Quantification and classification. Annu. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Gentekaki, E.; Curtis, B.; Archibald, J.M.; Roger, A.J. Lateral Gene transfer in the adaptation of the anaerobic parasite Blastocystis to the gut. Curr. Biol. 2017, 27, 807–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imachi, H.; Nobu, M.K.; Nakahara, N.; Morono, Y.; Ogawara, M.; Takaki, Y.; Takano, Y.; Uematsu, K.; Ikuta, T.; Ito, M.; et al. Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 2020, 577, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution: Do mothers matter most? BJOG 2020, 127, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, H.J.; Ball, H.J. Efficient tryptophan-catabolizing activity is consistently conserved through evolution of TDO enzymes, but not IDO enzymes. J. Exp. Zool. B Mol. Dev. Evol. 2015, 324, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Stairs, C.W.; Roger, A.J.; Hampl, V. Eukaryotic pyruvate formate lyase and its activating enzyme were acquired laterally from a firmicute. Mol. Biol. Evol. 2011, 28, 2087–2099. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, D.; Lauritsen, F.R.; Degn, H. The parasitic flagellates Trichomonas vaginalis and Tritrichomonas foetus produce indole and dimethyl disulphide: Direct characterization by membrane inlet tandem mass spectrometry. Microbiology 1991, 137, 1743–1747. [Google Scholar] [CrossRef] [Green Version]
- Nývltová, E.; Šut’ák, R.; Žárský, V.; Harant, K.; Hrdý, I.; Tachezy, J. Lateral gene transfer of p-cresol- and indole-producing enzymes from environmental bacteria to Mastigamoeba balamuthi. Environ. Microbiol. 2017, 19, 1091–1102. [Google Scholar] [CrossRef]
- Eichinger, L.; Pachebat, J.A.; Glockner, G.; Rajandream, M.A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67–79. [Google Scholar] [CrossRef]
- Xu, W.; Gavia, D.J.; Tang, Y. Biosynthesis of fungal indole alkaloids. Nat. Prod. Rep. 2014, 31, 1474–1487. [Google Scholar] [CrossRef] [Green Version]
- Halsey, C.R.; Lei, S.; Wax, J.K.; Lehman, M.K.; Nuxoll, A.S.; Steinke, L.; Sadykov, M.; Powers, R.; Fey, P.D. Amino acid catabolism in Staphylococcus aureus and the function of carbon catabolite repression. mBio 2017, 8, e01434-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Mentel, M.; van Hellemond, J.J.; Henze, K.; Woehle, C.; Gould, S.B.; Yu, R.Y.; van der Giezen, M.; Tielens, A.G.; Martin, W.F. Biochemistry and evolution of anaerobic energy metabolism in eukaryotes. Microbiol. Mol. Biol. Rev. 2012, 76, 444–495. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Jerlström-Hultqvist, J.; Einarsson, E.; Astvaldsson, A.; Svärd, S.G.; Andersson, J.O. The genome of Spironucleus salmonicida highlights a fish pathogen adapted to fluctuating environments. PLoS Genet. 2014, 10, e1004053. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.L.; Youssef, N.H.; Hanafy, R.A.; Couger, M.B.; Stajich, J.E.; Wang, Y.; Baker, K.; Dagar, S.S.; Griffith, G.W.; Farag, I.F.; et al. Horizontal gene transfer as an indispensable driver for evolution of neocallimastigomycota into a distinct gut-dwelling fungal lineage. Appl. Environ. Microbiol. 2019, 85, e00988-19. [Google Scholar] [CrossRef] [Green Version]
- Bunsangiam, S.; Sakpuntoon, V.; Srisuk, N.; Ohashi, T.; Fujiyama, K.; Limtong, S. Biosynthetic Pathway of Indole-3-Acetic Acid in Basidiomycetous Yeast Rhodosporidiobolus fluvialis. Mycobiology 2019, 47, 292–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteve, C.; Gutierrez, M.C.; Ventosa, A. Aeromonas encheleia sp. nov., isolated from European eels. Int. J. Syst. Bacteriol. 1995, 45, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Park, S.; Lee, J.M.; Park, S.; Jung, W.; Kang, J.-S.; Joo, H.M.; Seo, K.-W.; Kang, S.-H. Moritella dasanensis sp. nov., a psychrophilic bacterium isolated from the Arctic ocean. Int. J. Syst. Evol. Microbiol. 2008, 58, 817–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harf-Monteil, C.; Flèche, A.L.; Riegel, P.; Prévost, G.; Bermond, D.; Grimont, P.A.D.; Monteil, H. Aeromonas simiae sp. nov., isolated from monkey faeces. Int. J. Syst. Evol. Microbiol. 2004, 54, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Baere, T.D.; Verhelst, R.; Labit, C.; Verschraegen, G.; Wauters, G.; Claeys, G.; Vaneechoutte, M. Bacteremic Infection with Pantoea ananatis. J. Clin. Microbiol. 2004, 42, 4393–4395. [Google Scholar] [CrossRef] [Green Version]
- Pal, N.; Block, C.C.; Gardner, C.A.C. A real-time PCR differentiating Pantoea stewartii subsp. stewartii from P. stewartii subsp. indologenes in Corn Seed. Plant Dis. 2019, 103, 1474–1486. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsdóttir, D.B.K.; Helgason, S.; Høie, S.; Thoresen, O.F.; Wichardt, U.-P.; Wiklund, T. Identification of atypical Aeromonas salmonicida: Inter-laboratory evaluation and harmonization of methods. J. Appl. Microbiol. 1998, 84, 999–1006. [Google Scholar] [CrossRef]
- Hazir, S.; Stackebrandt, E.; Lang, E.; Schumann, P.; Ehlers, R.-U.; Keskin, N. Two new subspecies of photorhabdus luminescens, isolated from heterorhabditis bacteriophora (Nematoda: Heterorhabditidae): Photorhabdus luminescens subsp. kayaii subsp. nov. and Photorhabdus luminescens subsp. thracensis subsp. nov. Syst. Appl. Microbiol. 2004, 27, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Millership, S.E.; Chattopadhyay, B. Methods for the isolation of Aeromonas hydrophila and Plesiomonas shigelloides from faeces. J. Hyg. 1984, 92, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, Y.; Sasaki, T.; Ito, S.; Tamura, H.; Kunimatsu, K.; Kato, H. Identification and molecular characterization of tryptophanase encoded by tnaA in Porphyromonas gingivalis. Microbiology 2009, 155, 968–978. [Google Scholar] [CrossRef] [Green Version]
- Urbanczyk, H.; Ast, J.C.; Higgins, M.J.; Carson, J.; Dunlap, P.V. Reclassification of Vibrio fischeri, Vibrio logei, Vibrio salmonicida and Vibrio wodanis as Aliivibrio fischeri gen. nov., comb. nov., Aliivibrio logei comb. nov., Aliivibrio salmonicida comb. nov. and Aliivibrio wodanis comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2823–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, D.; Mouton, C.; Lapierre, P.; Kato, T.; Okuda, K.; Ménard, C. Porphyromonas gulae sp. nov., an anaerobic, gram-negative coccobacillus from the gingival sulcus of various animal hosts. Int. J. Syst. Evol. Microbiol. 2001, 51, 1179–1189. [Google Scholar] [CrossRef]
- Sasaki-Imamura, T.; Yoshida, Y.; Suwabe, K.; Yoshimura, F.; Kato, H. Molecular basis of indole production catalyzed by tryptophanase in the genus Prevotella. FEMS Microbiol. Lett. 2011, 322, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, S.G.; Birk, R.J.; Zabransky, R.J. Differences in susceptibilities of species of the Bacteroides fragilis group to several beta-lactam antibiotics: Indole production as an indicator of resistance. Antimicrob. Agents Chemother. 1982, 22, 628–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bale, M.J.; Mclaws, S.M.; Matsen, J.M. The spot indole test for identification of swarming proteus. Am. J. Clin. Pathol. 1985, 83, 87–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-S.; Roh, S.W.; Bae, J.-W. Bacteroides faecis sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2010, 60, 2572–2576. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.M.; Wright, J.W. Spot indole test: Evaluation of four reagents. J. Clin. Microbiol. 1982, 15, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakir, M.A.; Kitahara, M.; Sakamoto, M.; Matsumoto, M.; Benno, Y. Bacteroides intestinalis sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2006, 56, 151–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, A.L.; Yeoman, P.; Rasburn, J.W.; Ng, M. Sensitive reagents for detection of indole production by bacteria. Zent. für Bakteriol. Mikrobiol. Hygiene. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1986, 262, 195–202. [Google Scholar] [CrossRef]
- Alves, M.S.; da Silva Dias, R.C.; de Castro, A.C.D.; Riley, L.W.; Moreira, B.M. Identification of clinical isolates of indole-positive and indole-negative Klebsiella spp. J. Clin. Microbiol. 2006, 44, 3640–3646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.L.; Moore, W.E.C.; Moore, L.V.H. Bacteroides caccae sp. nov., Bacteroides merdae sp. nov., and Bacteroides stercoris sp. nov. Isolated from Human Feces. Int. J. Syst. Evol. Microbiol. 1986, 36, 499–501. [Google Scholar] [CrossRef] [Green Version]
- Adhikary, S.; Nicklas, W.; Bisgaard, M.; Boot, R.; Kuhnert, P.; Waberschek, T.; Aalbæk, B.; Korczak, B.; Christensen, H. Rodentibacter gen. nov. including Rodentibacter pneumotropicus comb. nov., Rodentibacter heylii sp. nov., Rodentibacter myodis sp. nov., Rodentibacter ratti sp. nov., Rodentibacter heidelbergensis sp. nov., Rodentibacter trehalosifermentans sp. nov., Rodentibacter rarus sp. nov., Rodentibacter mrazii and two genomospecies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1793–1806. [Google Scholar] [CrossRef] [PubMed]
- Fellström, C.; Karlsson, M.; Pettersson, B.; Zimmerman, U.; Gunnarsson, A.; Aspan, A. Emended descriptions of indole negative and indole positive isolates of Brachyspira (Serpulina) hyodysenteriae. Vet. Microbiol. 1999, 70, 225–238. [Google Scholar] [CrossRef]
- Antón, J.; Oren, A.; Benlloch, S.; Rodriguez-Valera, F.; Amann, R.; Rossello-Mora, R. Salinibacter ruber gen. nov., sp. nov., a novel, extreme halophilic member of the Bacteria from saltern crystallizer ponds. Int. J. Syst. Evol. Microbiol. 2002, 52, 485–491. [Google Scholar] [CrossRef]
- Sebek, O.K.; JÄGer, H. Divergent pathways of indole metabolism in chromobacterium violaceum. Nature 1962, 196, 793–795. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.P. The relationship of the enterobacterium A 12 (Sachs) to Shigella boydii 14. Microbiology 1961, 26, 535–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panpatte, D. Chryseobacterium indologenes a novel root nodule endophyte in vigna radiata. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 836–844. [Google Scholar] [CrossRef]
- Martino, P.D.; Fursy, R.; Bret, L.; Sundararaju, B.; Phillips, R.S. Indole can act as an extracellular signal to regulate biofilm formation of Escherichia coli and other indole-producing bacteria. Can. J. Microbiol. 2003, 49, 443–449. [Google Scholar] [CrossRef]
- Ribeiro, T.G.; Gonçalves, B.R.; da Silva, M.S.; Novais, Â.; Machado, E.; Carriço, J.A.; Peixe, L. Citrobacter portucalensis sp. nov., isolated from an aquatic sample. Int. J. Syst. Evol. Microbiol. 2017, 67, 3513–3517. [Google Scholar] [CrossRef]
- Iversen, C.; Mullane, N.; McCardell, B.; Tall, B.D.; Lehner, A.; Fanning, S.; Stephan, R.; Joosten, H. Cronobacter gen. nov., a new genus to accommodate the biogroups of Enterobacter sakazakii, and proposal of Cronobacter sakazakii gen. nov., comb. nov., Cronobacter malonaticus sp. nov., Cronobacter turicensis sp. nov., Cronobacter muytjensii sp. nov., Cronobacter dublinensis sp. nov., Cronobacter genomospecies 1, and of three subspecies, Cronobacter dublinensis subsp. dublinensis subsp. nov., Cronobacter dublinensis subsp. lausannensis subsp. nov. and Cronobacter dublinensis subsp. lactaridi subsp. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Ngadze, E.; Coutinho, T.A.; van der Waals, J.E. First report of soft rot of potatoes caused by dickeya dadantii in Zimbabwe. Plant Dis. 2010, 94, 1263. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Yasuoka, S.; Aono, Y.; Nakayama, T.; Ohki, T.; Sayama, M.; Maoka, T. Biochemical, physiological, and molecular characterization of Dickeya dianthicola (formerly named Erwinia chrysanthemi) causing potato blackleg disease in Japan. J. Gen. Plant Pathol. 2018, 84, 124–136. [Google Scholar] [CrossRef]
- Nuidate, T.; Tansila, N.; Saengkerdsub, S.; Kongreung, J.; Bakkiyaraj, D.; Vuddhakul, V. Role of indole production on virulence of vibrio cholerae using galleria mellonella larvae model. Indian J. Microbiol. 2016, 56, 368–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Wolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Lojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Ben-Haim, Y.; Thompson, F.L.; Thompson, C.C.; Cnockaert, M.C.; Hoste, B.; Swings, J.; Rosenberg, E. Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral Pocillopora damicornis. Int. J. Syst. Evol. Microbiol. 2003, 53, 309–315. [Google Scholar] [CrossRef]
- Kim, D.; Baik, K.S.; Hwang, Y.S.; Choi, J.-S.; Kwon, J.; Seong, C.N. Vibriohemicentroti sp. nov., an alginate lyase-producing bacterium, isolated from the gut microflora of sea urchin (Hemicentrotus pulcherrimus). Int. J. Syst. Evol. Microbiol. 2013, 63, 3697–3703. [Google Scholar] [CrossRef] [Green Version]
- Buján, N.; Toranzo, A.E.; Magariños, B. Edwardsiella piscicida: A significant bacterial pathogen of cultured fish. Dis. Aquat. Org. 2018, 131, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Raguénès, G.; Christen, R.; Guezennec, J.; Pignet, P.; Barbier, G. Vibrio diabolicus sp. nov., a new polysaccharide-secreting organism isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana. Int. J. Syst. Evol. Microbiol. 1997, 47, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Lopez, D.L. Mini review: New pathogen profiles: Elizabethkingia anophelis. Diagn. Microbiol. Infect. Dis. 2017, 88, 201–205. [Google Scholar] [CrossRef]
- Kim, K.K.; Kim, M.K.; Lim, J.H.; Park, H.Y.; Lee, S.-T. Transfer of Chryseobacterium meningosepticum and Chryseobacterium miricola to Elizabethkingia gen. nov. as Elizabethkingia meningoseptica comb. nov. and Elizabethkingia miricola comb. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1287–1293. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gil, B.; Thompson, F.L.; Thompson, C.C.; Swings, J. Vibrio pacinii sp. nov., from cultured aquatic organisms. Int. J. Syst. Evol. Microbiol. 2003, 53, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, S.; Tsuruya, Y.; Fukui, Y.; Sawabe, T.; Yokota, A.; Kogure, K.; Higgins, M.; Carson, J.; Thompson, F.L. Vibrio jasicida sp. nov., a member of the Harveyi clade, isolated from marine animals (packhorse lobster, abalone and Atlantic salmon). Int. J. Syst. Evol. Microbiol. 2012, 62, 1864–1870. [Google Scholar] [CrossRef]
- Thompson, F.L.; Hoste, B.; Thompson, C.C.; Goris, J.; Gomez-Gil, B.; Huys, L.; De Vos, P.; Swings, J. Enterovibrio norvegicus gen. nov., sp. nov., isolated from the gut of turbot (Scophthalmus maximus) larvae: A new member of the family Vibrionaceae. Int. J. Syst. Evol. Microbiol. 2002, 52, 2015–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Konno, T.; Kashio, H.; Suzuki, S.; Kumagai, Y. Isolation and characterization of Escherichia albertii from environmental water in Akita prefecture, Japan. Jpn. J. Food Microbiol. 2020, 37, 81–86. [Google Scholar] [CrossRef]
- Bueschkens, D.H.; Stiles, M.E. Escherichia coli variants for gas and indole production at elevated incubation temperatures. Appl. Environ. Microbiol. 1984, 48, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Kirchberger, P.C.; Turnsek, M.; Hunt, D.E.; Haley, B.J.; Colwell, R.R.; Polz, M.F.; Tarr, C.L.; Boucher, Y. Vibriometoecus sp. nov., a close relative of Vibrio cholerae isolated from coastal brackish ponds and clinical specimens. Int. J. Syst. Evol. Microbiol. 2014, 64, 3208–3214. [Google Scholar] [CrossRef] [PubMed]
- Farmer, J.J.; Fanning, G.R.; Davis, B.R.; O’Hara, C.M.; Riddle, C.; Hickman-Brenner, F.W.; Asbury, M.A.; Lowery, V.A.; Brenner, D.J. Escherichia fergusonii and Enterobacter taylorae, two new species of Enterobacteriaceae isolated from clinical specimens. J. Clin. Microbiol. 1985, 21, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Jin, D.; Lan, R.; Wang, Y.; Meng, Q.; Dai, H.; Lu, S.; Hu, S.; Xu, J. Escherichia marmotae sp. nov., isolated from faeces of Marmota himalayana. Int. J. Syst. Evol. Microbiol. 2015, 65, 2130–2134. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.; Park, S.-N.; Lim, Y.K.; Shin, Y.; Paek, J.; Hwang, C.H.; Chang, Y.-H.; Kook, J.-K. Fusobacterium hwasookii sp. nov., Isolated from a human periodontitis lesion. Curr. Microbiol. 2015, 70, 169–175. [Google Scholar] [CrossRef]
- Tan, Z.L.; Nagaraja, T.G.; Chengappa, M.M. Selective enumeration of Fusobacterium necrophorum from the bovine rumen. Appl. Environ. Microbiol. 1994, 60, 1387–1389. [Google Scholar] [CrossRef] [Green Version]
- Thompson, F.L.; Hoste, B.; Vandemeulebroecke, K.; Swings, J. Reclassification of Vibrio hollisae as Grimontia hollisae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1615–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCrea, K.W.; Xie, J.; LaCross, N.; Patel, M.; Mukundan, D.; Murphy, T.F.; Marrs, C.F.; Gilsdorf, J.R. Relationships of Nontypeable Haemophilus influenzae strains to hemolytic and nonhemolytic Haemophilus haemolyticus strains. J. Clin. Microbiol. 2008, 46, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Toma, S.; Lafleur, L. Survey on the Incidence of Yersinia enterocolitica infection in Canada. Appl. Microbiol. 1974, 28, 469–473. [Google Scholar] [CrossRef]
- Oberhofer, T.R.; Back, A.E. Biotypes of Haemophilus encountered in clinical laboratories. J. Clin. Microbiol. 1979, 10, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Ursing, J.; Don Brennert, J.; Bercovier, H.; Richard Fanning, G.; Steigerwalt, A.G.; Brault, J.; Mollaret, H.H. Yersinia frederiksenii: A new species of enterobacteriaceae composed of rhamnose-positive strains (formerly called atypicalyersinia enterocolitica or Yersinia enterocolitica-Like). Curr. Microbiol. 1980, 4, 213–217. [Google Scholar] [CrossRef]
- Angen, Ø.; Ahrens, P.; Kuhnert, P.; Christensen, H.; Mutters, R. Proposal of Histophilus somni gen. nov., sp. nov. for the three species incertae sedis ‘Haemophilus somnus’, ‘Haemophilus agni’ and ‘Histophilus ovis’. Int. J. Syst. Evol. Microbiol 2003, 53, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.J.; Bercovier, H.; Ursing, J.; Alonso, J.M.; Steigerwalt, A.G.; Fanning, G.R.; Carter, G.P.; Mollaret, H.H. Yersinia intermedia: A new species of enterobacteriaceae composed of rhamnose-positive, melibiose-positive, raffinose-positive strains (formerly called Yersinia enterocolitica or Yersinia enterocolitica-like). Curr. Microbiol. 1980, 4, 207–212. [Google Scholar] [CrossRef]
- Saha, R.; Farrance, C.E.; Verghese, B.; Hong, S.; Donofrio, R.S. Klebsiella michiganensis sp. nov., A new bacterium isolated from a tooth brush holder. Curr. Microbiol. 2013, 66, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Wauters, G.; Janssens, M.; Steigerwalt, A.G.; Brenner, D.J. Yersinia mollaretii sp. nov. and Yersinia bercovieri sp. nov., Formerly Called Yersinia enterocolitica Biogroups 3A and 3B. Int. J. Syst. Evol. Microbiol. 1988, 38, 424–429. [Google Scholar] [CrossRef] [Green Version]
- Maslow, J.N.; Brecher, S.M.; Adams, K.S.; Durbin, A.; Loring, S.; Arbeit, R.D. Relationship between indole production and differentiation of Klebsiella species: Indole-positive and -negative isolates of Klebsiella determined to be clonal. J. Clin. Microbiol. 1993, 31, 2000–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merhej, V.; Adékambi, T.; Pagnier, I.; Raoult, D.; Drancourt, M. Yersinia massiliensis sp. nov., isolated from fresh water. Int. J. Syst. Evol. Microbiol. 2008, 58, 779–784. [Google Scholar] [CrossRef]
- Tamura, K.; Sakazaki, R.; Kosako, Y.; Yoshizaki, E. Leclercia adecarboxylata gen. Nov., Comb. Nov., formerly known as Escherichia adecarboxylata. Curr. Microbiol. 1986, 13, 179–184. [Google Scholar] [CrossRef]
- Lawson, P.A.; Song, Y.; Liu, C.; Molitoris, D.R.; Vaisanen, M.-L.; Collins, M.D.; Finegold, S.M. Anaerotruncus colihominis gen. nov., sp. nov., from human faeces. Int. J. Syst. Evol. Microbiol. 2004, 54, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Sasi Jyothsna, T.S.; Tushar, L.; Sasikala, C.; Ramana, C.V. Paraclostridium benzoelyticum gen. nov., sp. nov., isolated from marine sediment and reclassification of Clostridium bifermentans as Paraclostridium bifermentans comb. nov. Proposal of a new genus Paeniclostridium gen. nov. to accommodate Clostridium sordellii and Clostridium ghonii. Int. J. Syst. Evol. Microbiol. 2016, 66, 1268–1274. [Google Scholar] [CrossRef]
- Nishida, S.; Nakagawara, G. Isolation of toxigenic strains of clostridium novyi from soil. J. Bacteriol. 1964, 88, 1636–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, G.L.; Dowell, V.R. Comparison of three reagents for detecting indole production by anaerobic bacteria in microtest systems. J. Clin. Microbiol. 1983, 18, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Baek, C.; Yi, H. A report of 22 unrecorded bacterial species in Korea, isolated from Namhangang. J. Species Res. 2018, 7, 114–122. [Google Scholar] [CrossRef]
- Clark, D.T.; Soory, M. The metabolism of cholesterol and certain hormonal steroids by Treponema denticola. Steroids 2006, 71, 352–363. [Google Scholar] [CrossRef]
- Kuhnert, P.; Brodard, I.; Alsaaod, M.; Steiner, A.; Stoffel, M.H.; Jores, J. Treponema phagedenis (ex Noguchi 1912) Brumpt 1922 sp. nov., nom. rev., isolated from bovine digital dermatitis. Int. J. Syst. Evol. Microbiol. 2020, 70, 2115–2123. [Google Scholar] [CrossRef]
- Cui, H.-L.; Qiu, X.-X. Salinarubrum litoreum gen. nov., sp. nov.: A new member of the family Halobacteriaceae isolated from Chinese marine solar salterns. Antonie Van Leeuwenhoek 2014, 105, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Cui, H.-L. Halostella pelagica sp. nov. and Halostella litorea sp. nov., isolated from salted brown alga Laminaria. Int. J. Syst. Evol. Microbiol. 2020, 70, 1969–1976. [Google Scholar] [CrossRef]
- Wang, S.; Yang, Q.; Liu, Z.-H.; Sun, L.; Wei, D.; Zhang, J.-Z.; Song, J.-Z.; Yuan, H.-F. Haloterrigena daqingensis sp. nov., an extremely haloalkaliphilic archaeon isolated from a saline–alkaline soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 2267–2271. [Google Scholar] [CrossRef] [Green Version]
- Juez, G.; Rodriguez-Valera, F.; Ventosa, A.; Kushner, D.J. Haloarcula hispanica spec. nov. and Haloferax gibbonsii spec, nov., Two new species of extremely halophilic archaebacteria. Syst. Appl. Microbiol. 1986, 8, 75–79. [Google Scholar] [CrossRef]
- Roh, S.W.; Nam, Y.-D.; Chang, H.-W.; Kim, K.-H.; Sung, Y.; Kim, M.-S.; Oh, H.-M.; Bae, J.-W. Haloterrigena jeotgali sp. nov., an extremely halophilic archaeon from salt-fermented food. Int. J. Syst. Evol. Microbiol. 2009, 59, 2359–2363. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Ginzburg, M.; Ginzburg, B.Z.; Hochstein, L.I.; Volcani, B.E. Haloarcula marismortui (Volcani) sp. nov., nom. rev., an Extremely Halophilic Bacterium from the Dead Sea. Int. J. Syst. Evol. Microbiol. 1990, 40, 209–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, S.; Hagagy, N. Genome sequence of carboxylesterase, carboxylase and xylose isomerase producing alkaliphilic haloarchaeon Haloterrigena turkmenica WANU15. Genom. Data 2016, 7, 70–72. [Google Scholar] [CrossRef] [Green Version]
- Hezayen, F.F.; Rehm, B.H.; Tindall, B.J.; Steinbüchel, A. Transfer of Natrialba asiatica B1T to Natrialba taiwanensis sp. nov. and description of Natrialba aegyptiaca sp. nov., a novel extremely halophilic, aerobic, non-pigmented member of the Archaea from Egypt that produces extracellular poly(glutamic acid). Int. J. Syst. Evol. Microbiol. 2001, 51, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Merkel, A.Y.; Messina, E.; Yakimov, M.M.; Itoh, T.; Mesbah, N.M.; Wiegel, J.; Oren, A. Reclassification of the genus Natronolimnobius: Proposal of two new genera, Natronolimnohabitans gen. nov. to accommodate Natronolimnobius innermongolicus and Natrarchaeobaculum gen. nov. to accommodate Natronolimnobius aegyptiacus and Natronolimnobius sulfurireducens. Int. J. Syst. Evol. Microbiol. 2020, 70, 3399–3405. [Google Scholar] [CrossRef]
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D. Proposal of a new halobacterial genus Natrinema gen. nov., with two species Natrinema pellirubrum nom. nov. and Natrinema pallidum nom. nov. Int. J. Syst. Evol. Microbiol. 1998, 48, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Hassanshahian, M.; Jafar, M. Isolation and characterization of Halobacterium salinarum from saline lakes in Iran. Jundishapur J. Microbiol. 2011, 4, 59–65. [Google Scholar]
- Xin, H.; Itoh, T.; Zhou, P.; Suzuki, K.; Kamekura, M.; Nakase, T. Natrinema versiforme sp. nov., an extremely halophilic archaeon from Aibi salt lake, Xinjiang, China. Int. J. Syst. Evol. Microbiol. 2000, 50, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-W.; Wu, M.; Zhou, P.-J.; Liu, S.-J. Halobiforma lacisalsi sp. nov., isolated from a salt lake in China. Int. J. Syst. Evol. Microbiol. 2005, 55, 1949–1952. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Haloferax alexandrinus sp. nov., an extremely halophilic canthaxanthin-producing archaeon from a solar saltern in Alexandria (Egypt). Int. J. Syst. Evol. Microbiol. 2002, 52, 729–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akmoussi-Toumi, S.; Khemili-Talbi, S.; Ferioune, I.; Kebbouche-Gana, S. Purification and characterization of an organic solvent-tolerant and detergent-stable lipase from Haloferax mediterranei CNCMM 50101. Int. J. Biol. Macromol. 2018, 116, 817–830. [Google Scholar] [CrossRef]
- Cui, H.-L.; Tohty, D.; Feng, J.; Zhou, P.-J.; Liu, S.-J. Natronorubrum aibiense sp. nov., an extremely halophilic archaeon isolated from Aibi salt lake in Xin-Jiang, China, and emended description of the genus Natronorubrum. Int. J. Syst. Evol. Microbiol. 2006, 56, 1515–1517. [Google Scholar] [CrossRef] [Green Version]
- Franzmann, P.D.; Stackebrandt, E.; Sanderson, K.; Volkman, J.K.; Cameron, D.E.; Stevenson, P.L.; McMeekin, T.A.; Burton, H.R. Halobacterium lacusprofundi sp. nov., a Halophilic Bacterium Isolated from Deep Lake, Antarctica. Syst. Appl. Microbiol. 1988, 11, 20–27. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, P.; Tian, X. Characterization of two novel haloalkaliphilic archaea Natronorubrum bangense gen. nov., sp. nov. and Natronorubrum tibetense gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A.; Montalvo-Rodríguez, R. Halogeometricum. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–9. [Google Scholar]
- Gutiérrez, M.C.; Castillo, A.M.; Kamekura, M.; Xue, Y.; Ma, Y.; Cowan, D.A.; Jones, B.E.; Grant, W.D.; Ventosa, A. Halopiger xanaduensis gen. nov., sp. nov., an extremely halophilic archaeon isolated from saline Lake Shangmatala in Inner Mongolia, China. Int. J. Syst. Evol. Microbiol. 2007, 57, 1402–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sako, Y.; Nomura, N.; Uchida, A.; Ishida, Y.; Morii, H.; Koga, Y.; Hoaki, T.; Maruyama, T. Aeropyrum pernix gen. nov., sp. nov., a novel aerobic hyperthermophilic archaeon growing at temperatures up to 100 degrees C. Int. J. Syst. Bacteriol. 1996, 46, 1070–1077. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Kublanov, I.V.; Gavrilov, S.N.; Rojo, D.; Roman, P.; Golyshin, P.N.; Slepak, V.Z.; Smedile, F.; Ferrer, M.; Messina, E.; et al. Elemental sulfur and acetate can support life of a novel strictly anaerobic haloarchaeon. ISME J. 2016, 10, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorokin, D.Y.; Kublanov, I.V.; Yakimov, M.M.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S. Halanaeroarchaeum sulfurireducens gen. nov., sp. nov., the first obligately anaerobic sulfur-respiring haloarchaeon, isolated from a hypersaline lake. Int. J. Syst. Evol. Microbiol. 2016, 66, 2377–2381. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.C.; Chen, T.W.; Han, Y.A.; Ng, W.V.; Yang, C.S. Complete genome sequence of a new halophilic archaeon, Haloarcula taiwanensis, isolated from a solar saltern in Southern Taiwan. Genome Announc. 2018, 6, e01529-17. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Nucleotide sequence phylogenetic tree (arbitrarily rooted Maximum Likelihood tree) of the tnaA gene representing a total of 221 nucleotide sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values ≥70 (1000 replications) and bootstrap value representation in Figure S1. Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown).
Figure 1.
Nucleotide sequence phylogenetic tree (arbitrarily rooted Maximum Likelihood tree) of the tnaA gene representing a total of 221 nucleotide sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values ≥70 (1000 replications) and bootstrap value representation in Figure S1. Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown).

Figure 2.
Amino acid sequence phylogenetic tree (arbitrarily rooted Maximum Likelihood tree) of the tnaA gene representing a total of 221 amino acid sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values ≥70 (1000 replications) and bootstrap value representation in Figure S2. Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown).
Figure 2.
Amino acid sequence phylogenetic tree (arbitrarily rooted Maximum Likelihood tree) of the tnaA gene representing a total of 221 amino acid sequences belonging to 95 genera of various taxonomic groups. All the nodes represent bootstrap values ≥70 (1000 replications) and bootstrap value representation in Figure S2. Details of the phylogenetic analysis are provided in Methods. A complete list of genera and species and their taxonomic classifications are provided in Table 1. Species names are labelled according to phylogenetic classifications by the following colour codes. Gram-negative bacteria (blue), Gram-positive bacteria (green), archaea (red), fungi (purple), unicellular eukaryotes (orange), higher eukaryotes (brown).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Boya, B.R.; Kumar, P.; Lee, J.-H.; Lee, J. Diversity of the Tryptophanase Gene and Its Evolutionary Implications in Living Organisms. Microorganisms 2021, 9, 2156. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102156
AMA Style
Boya BR, Kumar P, Lee J-H, Lee J. Diversity of the Tryptophanase Gene and Its Evolutionary Implications in Living Organisms. Microorganisms. 2021; 9(10):2156. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102156
Chicago/Turabian StyleBoya, Bharath Reddy, Prasun Kumar, Jin-Hyung Lee, and Jintae Lee. 2021. "Diversity of the Tryptophanase Gene and Its Evolutionary Implications in Living Organisms" Microorganisms 9, no. 10: 2156. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102156
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.