
Propionibacterium acnes-Derived Circulating Immune Complexes in Sarcoidosis Patients

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Plasma Sample Pretreatment Procedures
2.3. Sandwich ELISA for Detecting Plasma PLTA before or after the Pretreatment
2.4. Indirect ELISA for Detecting Plasma Anti-PLTA Antibodies
2.5. Statistical Analyses
3. Results
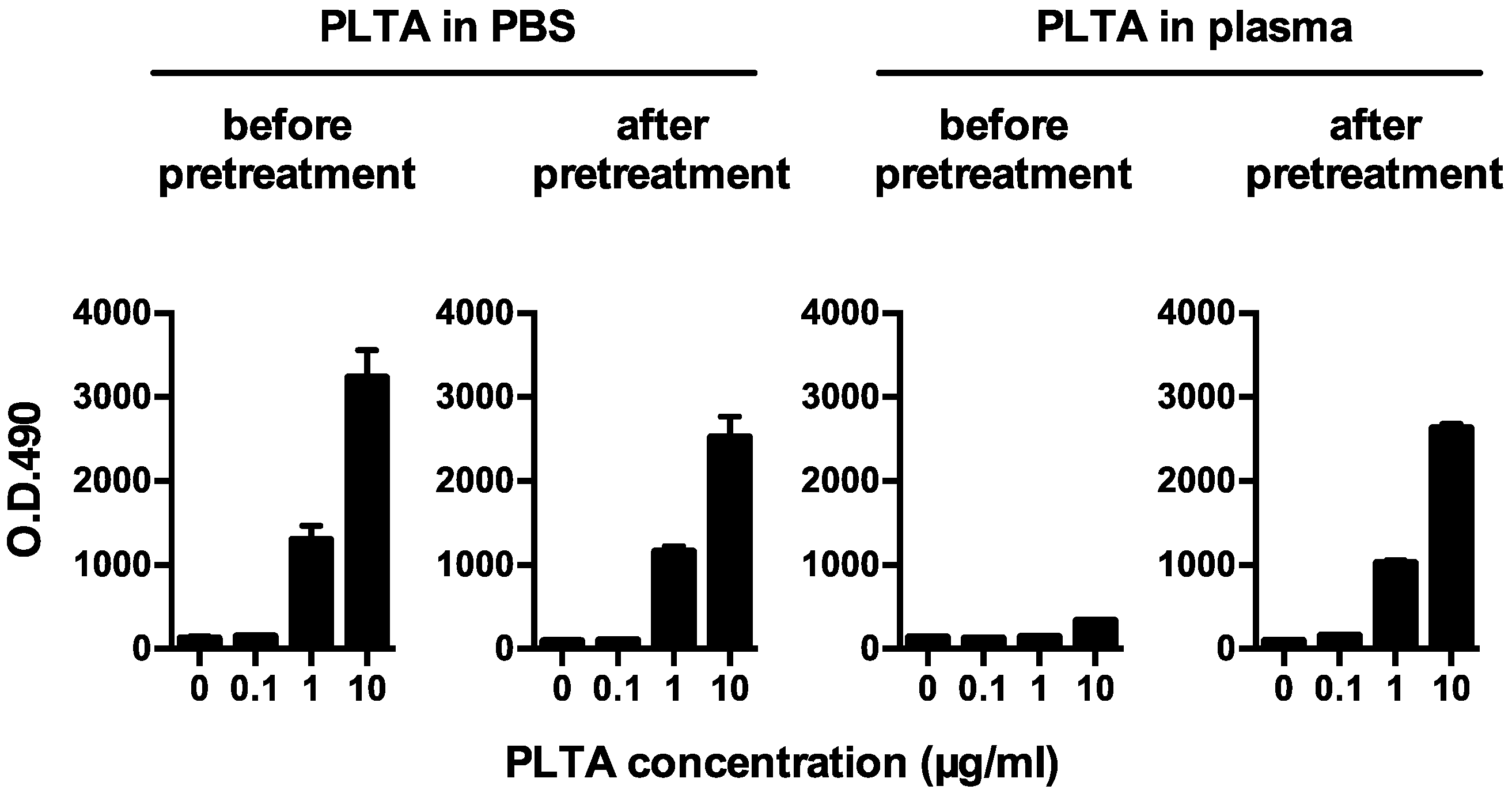
3.1. PLTA Detection Recovered by the Pretreatment
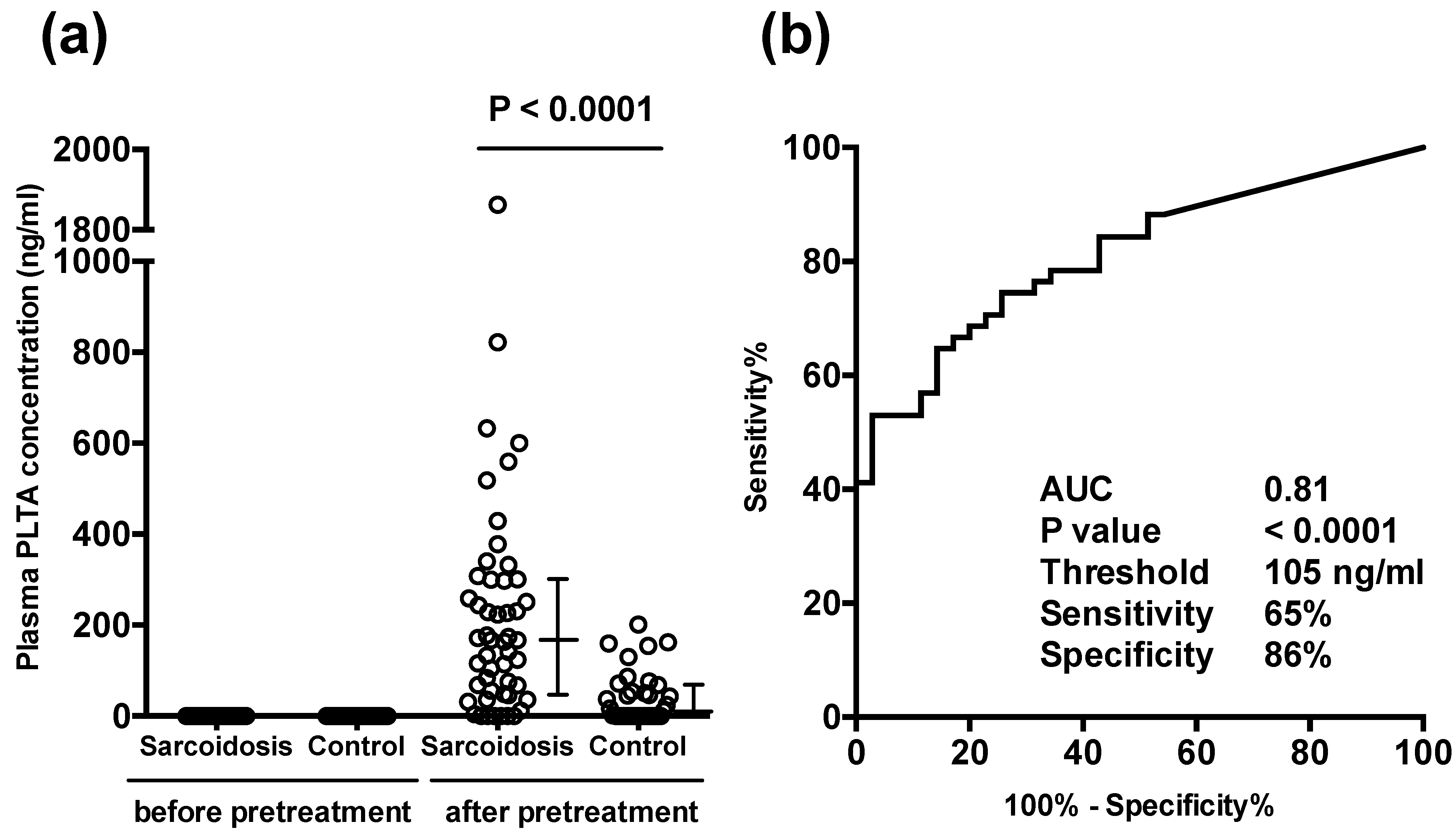
3.2. Plasma PLTA Concentrations before and after the Pretreatment
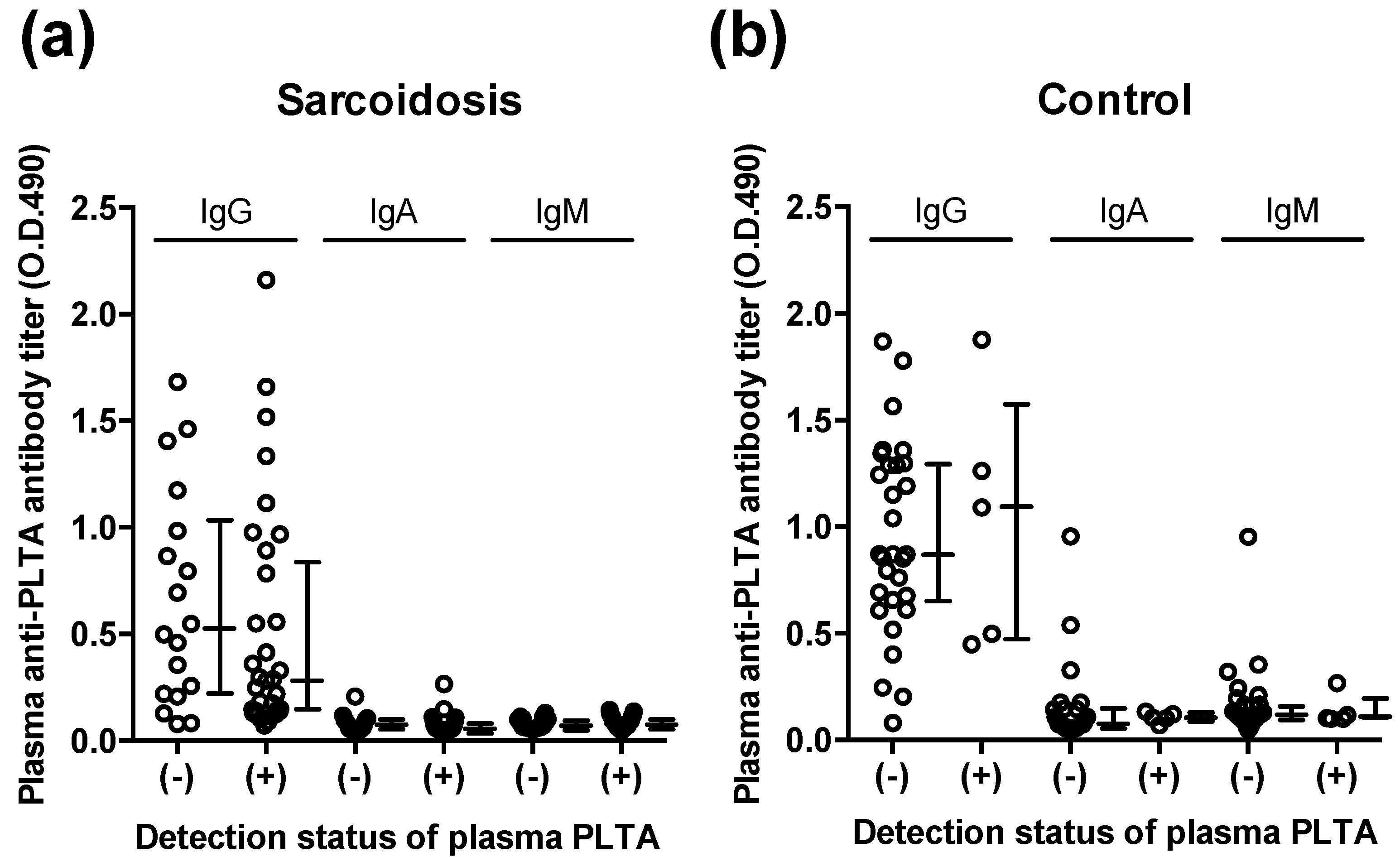
3.3. Plasma Anti-PLTA Antibody Titers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valeyre, D.; Prasse, A.; Nunes, H.; Uzunhan, Y.; Brillet, P.-Y.; Müller-Quernheim, J. Sarcoidosis. Lancet 2014, 383, 1155–1167. [Google Scholar] [CrossRef]
- Drent, M.; Crouser, E.D.; Grunewald, J. Challenges of sarcoidosis and its management. N. Engl. J. Med. 2021, 385, 1018–1032. [Google Scholar] [CrossRef]
- Tchernev, G.; Cardoso, J.C.; Chokoeva, A.A.; Verma, S.B.; Tana, C.; Ananiev, J.; Gulubova, M.; Philipov, S.; Kanazawa, N.; Nenoff, P.; et al. The “mystery” of cutaneous sarcoidosis: Facts and controversies. Int. J. Immunopathol. Pharmacol. 2014, 27, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Greaves, S.A.; Ravindran, A.; Santos, R.G.; Chen, L.; Falta, M.T.; Wang, Y.; Mitchell, A.M.; Atif, S.M.; Mack, D.G.; Tinega, A.N.; et al. CD4+ T cells in the lungs of acute sarcoidosis patients recognize an Aspergillus nidulans epitope. J. Exp. Med. 2021, 218, e20210785. [Google Scholar] [CrossRef]
- Alexeyev, O.A.; Dekio, I.; Layton, A.M.; Li, H.; Hughes, H.; Morris, T.; Zouboulis, C.C.; Patrick, S. Why we continue to use the name Propionibacterium acnes. Br. J. Dermatol. 2018, 179, 1227. [Google Scholar] [CrossRef]
- Abe, C.; Iwai, K.; Mikami, R.; Hosoda, Y. Frequent isolation of Propionibacterium acnes from sarcoidosis lymph nodes. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1984, 256, 541–547. [Google Scholar] [CrossRef]
- Homma, J.Y.; Abe, C.; Chosa, H.; Ueda, K.; Saegusa, J.; Nakayama, M.; Homma, H.; Washizaki, M.; Okano, H. Bacteriological investigation on biopsy specimens from patients with sarcoidosis. Jpn. J. Exp. Med. 1978, 48, 251–255. [Google Scholar]
- Eishi, Y.; Suga, M.; Ishige, I.; Kobayashi, D.; Yamada, T.; Takemura, T.; Takizawa, T.; Koike, M.; Kudoh, S.; Costabel, U.; et al. Quantitative analysis of mycobacterial and propionibacterial DNA in lymph nodes of Japanese and European patients with sarcoidosis. J. Clin. Microbiol. 2002, 40, 198–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishige, I.; Usui, Y.; Takemura, T.; Eishi, Y. Quantitative PCR of mycobacterial and propionibacterial DNA in lymph nodes of Japanese patients with sarcoidosis. Lancet 1999, 354, 120–123. [Google Scholar] [CrossRef]
- Yamada, T.; Eishi, Y.; Ikeda, S.; Ishige, I.; Suzuki, T.; Takemura, T.; Takizawa, T.; Koike, M. In situ localization of Propionibacterium acnes DNA in lymph nodes from sarcoidosis patients by signal amplification with catalysed reporter deposition. J. Pathol. 2002, 198, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Negi, M.; Takemura, T.; Guzman, J.; Uchida, K.; Furukawa, A.; Suzuki, Y.; Iida, T.; Ishige, I.; Minami, J.; Yamada, T.; et al. Localization of Propionibacterium acnes in granulomas supports a possible etiologic link between sarcoidosis and the bacterium. Mod. Pathol. 2012, 25, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Isshiki, T.; Homma, S.; Eishi, Y.; Yabe, M.; Koyama, K.; Nishioka, Y.; Yamaguchi, T.; Uchida, K.; Yamamoto, K.; Ohashi, K.; et al. Immunohistochemical Detection of Propionibacterium acnes in Granulomas for Differentiating Sarcoidosis from Other Granulomatous Diseases Utilizing an Automated System with a Commercially Available PAB Antibody. Microorganisms 2021, 9, 1668. [Google Scholar] [CrossRef]
- Ebe, Y.; Ikushima, S.; Yamaguchi, T.; Kohno, K.; Azuma, A.; Sato, K.; Ishige, I.; Usui, Y.; Takemura, T.; Eishi, Y. Proliferative response of peripheral blood mononuclear cells and levels of antibody to recombinant protein from Propionibacterium acnes DNA expression library in Japanese patients with sarcoidosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2000, 17, 256–265. [Google Scholar]
- Furusawa, H.; Suzuki, Y.; Miyazaki, Y.; Inase, N.; Eishi, Y. Th1 and Th17 immune responses to viable Propionibacterium acnes in patients with sarcoidosis. Respir. Investig. 2012, 50, 104–109. [Google Scholar] [CrossRef]
- Yorozu, P.; Furukawa, A.; Uchida, K.; Akashi, T.; Kakegawa, T.; Ogawa, T.; Minami, J.; Suzuki, Y.; Awano, N.; Furusawa, H.; et al. Propionibacterium acnes catalase induces increased Th1 immune response in sarcoidosis patients. Respir. Investig. 2015, 53, 161–169. [Google Scholar] [CrossRef]
- Schupp, J.C.; Tchaptchet, S.; Lützen, N.; Engelhard, P.; Müller-Quernheim, J.; Freudenberg, M.A.; Prasse, A. Immune response to Propionibacterium acnes in patients with sarcoidosis—In vivo and in vitro. BMC Pulm. Med. 2015, 15, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eishi, Y. Etiologic aspect of sarcoidosis as an allergic endogenous infection caused by Propionibacterium acnes. Biomed. Res. Int. 2013, 2013, 935289. [Google Scholar] [CrossRef] [Green Version]
- Eishi, Y. Etiologic link between sarcoidosis and Propionibacterium acnes. Respir. Investig. 2013, 51, 56–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Costabel, U.; McDowell, A.; Guzman, J.; Uchida, K.; Ohashi, K.; Eishi, Y. Immunohistochemical Detection of Potential Microbial Antigens in Granulomas in the Diagnosis of Sarcoidosis. J. Clin. Med. 2021, 10, 983. [Google Scholar] [CrossRef]
- Jones, J.V.; Cumming, R.H.; Asplin, C.M. Evidence for circulating immune complexes in erythema nodosum and early sarcoidosis. Ann. N. Y. Acad. Sci. 1976, 278, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.M.; NcNicol, M.W.; Burton-Kee, J.E.; Mowbray, J.F. Circulating immune complexes in sarcoidosis. Thorax 1980, 35, 286–289. [Google Scholar] [CrossRef] [Green Version]
- Hedfors, E.; Norberg, R. Evidence for circulating immune complexes in sarcoidosis. Clin. Exp. Immunol. 1974, 16, 493–496. [Google Scholar]
- Gupta, R.C.; Kueppers, F.; DeRemee, R.A.; Huston, K.A.; McDuffie, F.C. Pulmonary and extrapulmonary sarcoidosis in relation to circulating immune complexes: A quantification of immune complexes by two radioimmunoassays. Am. Rev. Respir. Dis. 1977, 116, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Daniele, R.P.; McMillan, L.J.; Dauber, J.H.; Rossman, M.D. Immune complexes in sarcoidosis: A correlation with activity and duration of disease. Chest 1978, 74, 261–264. [Google Scholar] [CrossRef] [PubMed]
- James, D.G.; Neville, E.; Walker, A. Immunology of sarcoidosis. Am. J. Med. 1975, 59, 388–394. [Google Scholar] [CrossRef]
- Eishi, Y.; Takemura, T.; Matsui, Y.; Hatakeyama, S. Pathogenesis of Granuloma Formation in Lymph Nodes with Sarcoidosis. In Sarcoidosis; Grassi, C., Rizzato, G., Pozzi, E., Eds.; Elsevier Science Publisher B.V.: Amsterdam, The Netherlands, 1988; pp. 143–146. [Google Scholar]
- Suzuki, Y.; Uchida, K.; Takemura, T.; Sekine, M.; Tamura, T.; Furukawa, A.; Hebisawa, A.; Sakakibara, Y.; Awano, N.; Amano, T.; et al. Propionibacterium acnes-derived insoluble immune complexes in sinus macrophages of lymph nodes affected by sarcoidosis. PLoS ONE 2018, 13, e0192408. [Google Scholar] [CrossRef] [PubMed]
- Hunninghake, G.W.; Costabel, U.; Ando, M.; Baughman, R.; Cordier, J.F.; du Bois, R.; Eklund, A.; Kitaichi, M.; Lynch, J.; Rizzato, G.; et al. ATS/ERS/WASOG statement on sarcoidosis. American Thoracic Society/European Respiratory Society/World Association of Sarcoidosis and other Granulomatous Disorders. Sarcoidosis Vasc. Diffus. Lung Dis. 1999, 16, 149–173. [Google Scholar]
- Bae, Y.; Ito, T.; Iida, T.; Uchida, K.; Sekine, M.; Nakajima, Y.; Kumagai, J.; Yokoyama, T.; Kawachi, H.; Akashi, T.; et al. Intracellular Propionibacterium acnes infection in glandular epithelium and stromal macrophages of the prostate with or without cancer. PLoS ONE 2014, 9, e90324. [Google Scholar] [CrossRef]
- Fischer, W. Bacterial Phosphoglycolipids and Lipoteichoic Acids. In Handbook of Lipid Research 6: Glycolipids, Phosphoglycolipids and Sulfoglycolipids; Kates, M., Ed.; Plenum Press: New York, NY, USA, 1990. [Google Scholar]
- Knox, K.W.; Wicken, A.J. Immunological properties of teichoic acids. Bacteriol. Rev. 1973, 37, 215–257. [Google Scholar] [CrossRef]
- Ishige, I.; Eishi, Y.; Takemura, T.; Kobayashi, I.; Nakata, K.; Tanaka, I.; Nagaoka, S.; Iwai, K.; Watanabe, K.; Takizawa, T.; et al. Propionibacterium acnes is the most common bacterium commensal in peripheral lung tissue and mediastinal lymph nodes from subjects without sarcoidosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2005, 22, 33–42. [Google Scholar]
- Fischer, N.; Mak, T.N.; Shinohara, D.B.; Sfanos, K.S.; Meyer, T.F.; Brüggemann, H. Deciphering the Intracellular Fate of Propionibacterium acnes in Macrophages. Biomed. Res. Int. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, K.; Eishi, Y.; Tahara, N.; Asakura, M.; Sakamoto, N.; Nakamura, K.; Takaya, Y.; Nakamura, T.; Yazaki, Y.; Yamaguchi, T.; et al. Japanese Antibacterial Drug Management for Cardiac Sarcoidosis (J-ACNES ): A multicenter, open-label, randomized, controlled study. J. Arrhythmia 2018, 34, 520–526. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | |
|---|---|
| Number | 51 |
| Subjects (male/female) | 20/31 |
| Age, years | 51 ± 16 |
| Chest X-ray Stage (0/I/II/III/IV/NA) | 2/18/24/3/1/3 |
| Angiotensin converting enzyme (U/I/37 °C) a | 22.8 ± 10.7 |
| Lysozyme (µg/mL) b | 12.6 ± 9.4 |
| Soluble interleukin-2 receptor (U/mL) c | 1235 ± 841 |
| Pulmonary function test | |
| % Forced vital capacity (%) d | 105 ± 14 |
| Forced expiratory volume in 1 s (%) e | 80 ± 10 |
| Bronchoalveolar lavage analysis | |
| total cell (105 cell/mL) | 2.7 ± 1.6 |
| macrophage (%) | 66 ± 20 |
| lymphocytes (%) | 32 ± 20 |
| CD4/CD8 ratio f | 5.9 ± 3.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchida, K.; Furukawa, A.; Yoneyama, A.; Furusawa, H.; Kobayashi, D.; Ito, T.; Yamamoto, K.; Sekine, M.; Miura, K.; Akashi, T.; et al. Propionibacterium acnes-Derived Circulating Immune Complexes in Sarcoidosis Patients. Microorganisms 2021, 9, 2194. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112194
Uchida K, Furukawa A, Yoneyama A, Furusawa H, Kobayashi D, Ito T, Yamamoto K, Sekine M, Miura K, Akashi T, et al. Propionibacterium acnes-Derived Circulating Immune Complexes in Sarcoidosis Patients. Microorganisms. 2021; 9(11):2194. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112194
Chicago/Turabian StyleUchida, Keisuke, Asuka Furukawa, Akiko Yoneyama, Haruhiko Furusawa, Daisuke Kobayashi, Takashi Ito, Kurara Yamamoto, Masaki Sekine, Keiko Miura, Takumi Akashi, and et al. 2021. "Propionibacterium acnes-Derived Circulating Immune Complexes in Sarcoidosis Patients" Microorganisms 9, no. 11: 2194. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112194