
Effects of High-Forage Diets Containing Raw Flaxseeds or Soybean on In Vitro Ruminal Fermentation, Gas Emission, and Microbial Profile

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diet Substrates
2.2. Rumen Fluid Preparation
2.3. Equipment and In Vitro Incubation
2.4. Sampling
2.5. Chemical Analysis and Calculations
2.6. DNA Extraction and Determination
2.7. Statistical Methods
3. Results
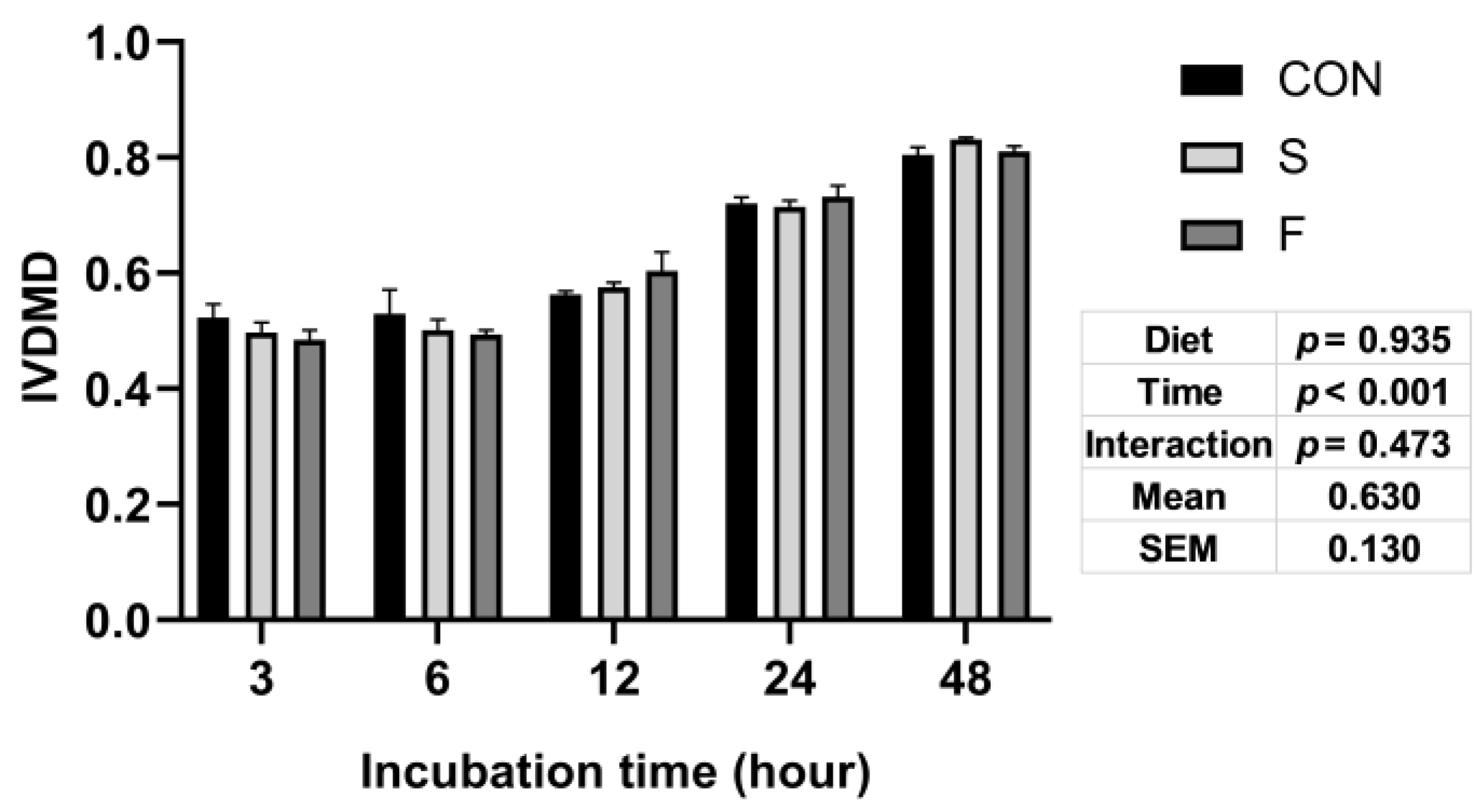
3.1. In Vitro Dry Matter Digestibility
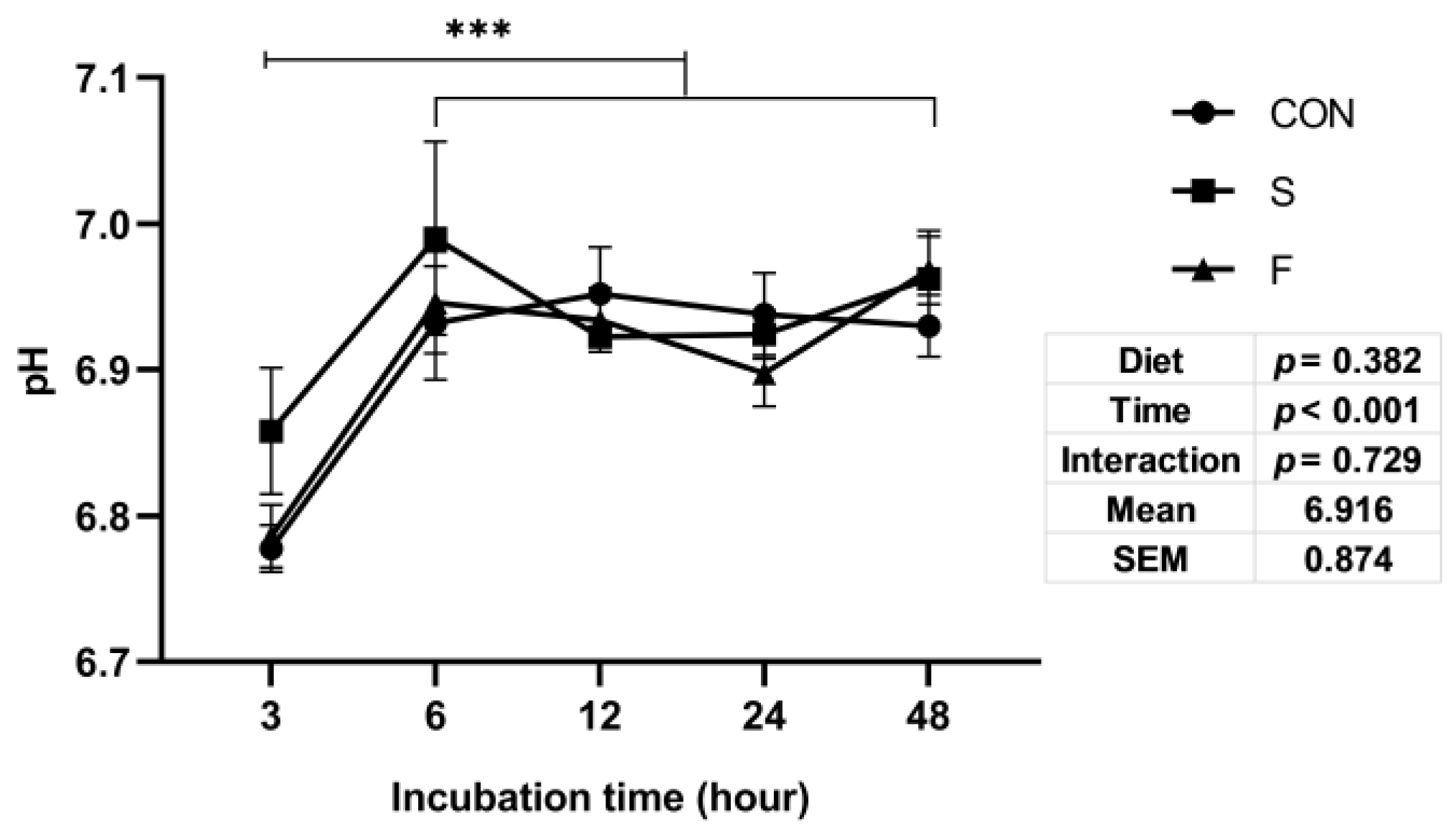
3.2. Culture Fluids pH
3.3. Volatile Fatty Acid Concentration
3.4. Ammonia–N Concentration
3.5. Microbial Protein Concentration
3.6. Gas Production and Kinetic Parameters
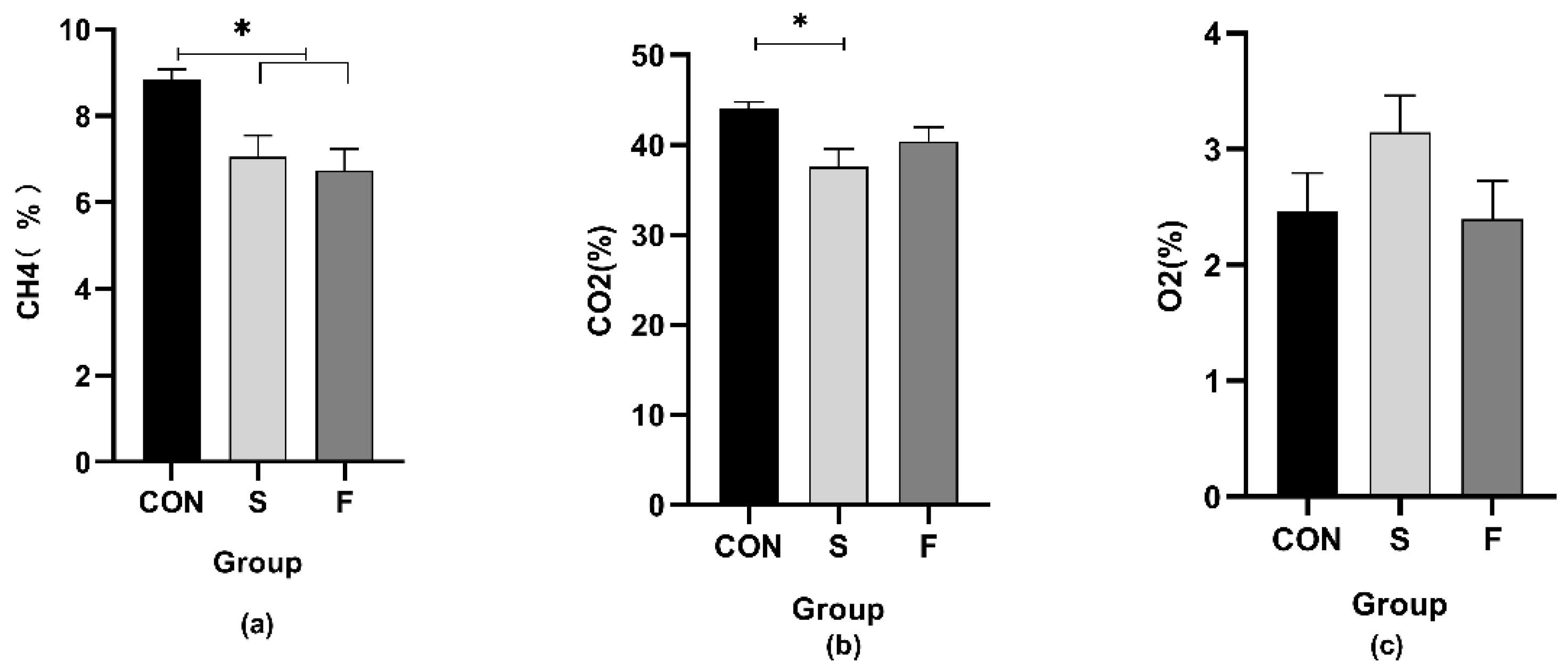
3.7. Gas Composition
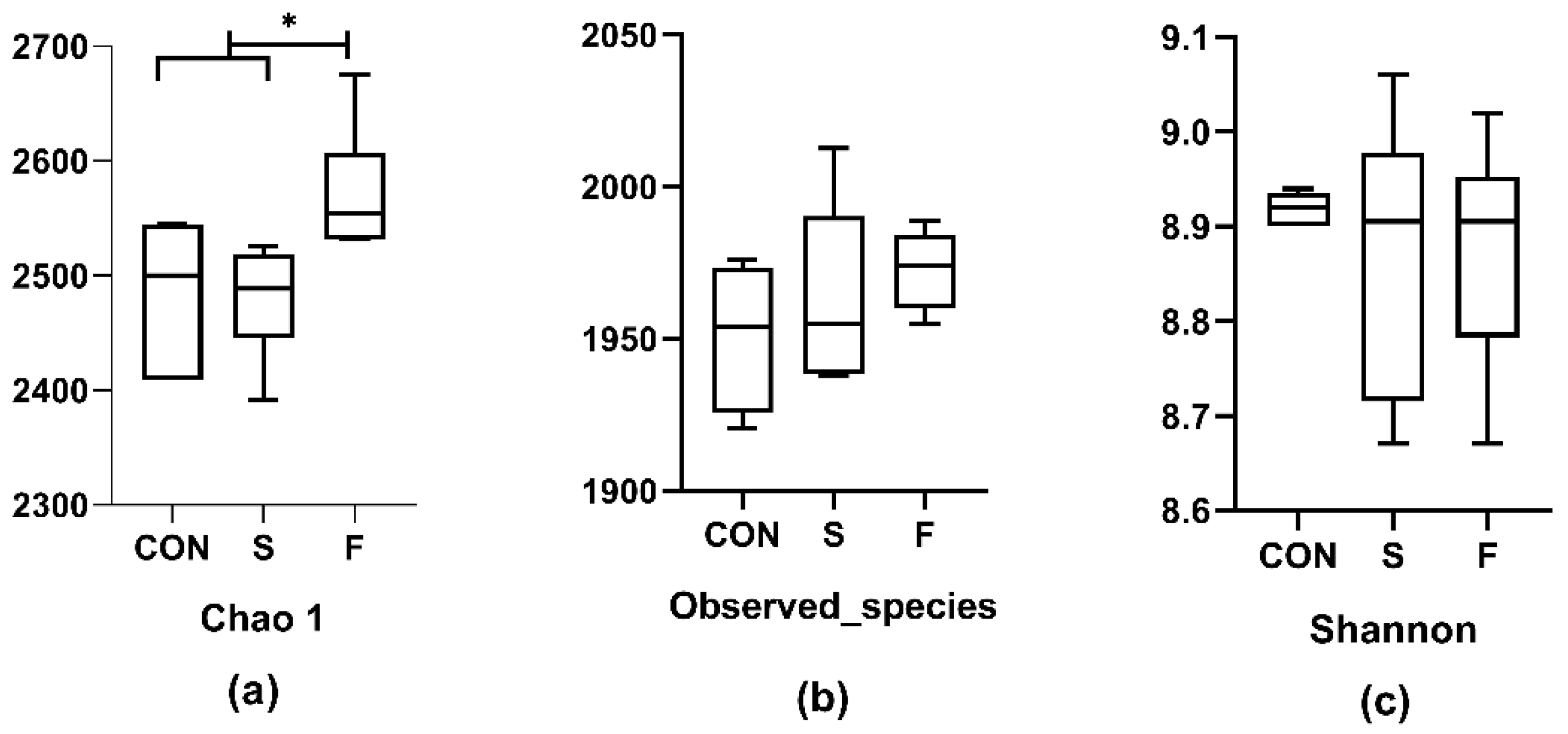
3.8. Rumen Microbe Diversity and Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Savoini, G.; Omodei Zorini, F.; Farina, G.; Agazzi, A.; Cattaneo, D.; Invernizzi, G. Effects of fat supplementation in dairy goats on lipid metabolism and health status. Animals 2019, 9, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huws, S.A.; Lee, M.R.; Muetzel, S.M.; Scott, M.B.; Wallace, R.J.; Scollan, N.D. Forage type and fish oil cause shifts in rumen bacterial diversity. FEMS Microbiol. Ecol. 2010, 73, 396–702. [Google Scholar] [CrossRef] [Green Version]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.D.; Castel, V.; Rosales, M.; Rosales, M.; de Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food & Agriculture Organisation: Rome, Italy, 2006. [Google Scholar]
- Beauchemin, K.A.; McGinn, S.M.; Petit, H.V. Methane abatement strategies for cattle: Lipid supplementation of diets. Can. J. Anim. Sci. 2007, 87, 431–440. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Huws, S.A.; Lee, M.R.; Scollan, N.D. Dietary transformation of lipid in the rumen microbial ecosystem. Asian-Australas. J. Anim. Sci. 2009, 22, 1341–1350. [Google Scholar] [CrossRef]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef] [Green Version]
- Weld, K.; Armentano, L. The effects of adding fat to diets of lactating dairy cows on total-tract neutral detergent fiber digestibility: A meta-analysis. J. Dairy Sci. 2017, 100, 1766–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leduc, M.; Létourneau-Montminy, M.-P.; Gervais, R.; Chouinard, P. Effect of dietary flax seed and oil on milk yield, gross composition, and fatty acid profile in dairy cows: A meta-analysis and meta-regression. J. Dairy Sci. 2017, 100, 8906–8927. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.; Lee, M.R.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 19th ed; Association of Official Analytical Chemists Inc.: Arlington, VA, USA, 2012; pp. 1048–1049. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle: 2001; National Academies Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Zhang, D.; Yang, H. In vitro ruminal methanogenesis of a hay-rich substrate in response to different combination supplements of nitrocompounds; pyromellitic diimide and 2-bromoethanesulphonate. Anim. Feed Sci. Technol. 2011, 163, 20–32. [Google Scholar] [CrossRef]
- Menke, K.H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Menke, K.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Lanzas, C.; Fox, D.; Pell, A. Digestion kinetics of dried cereal grains. Anim. Feed Sci. Technol. 2007, 136, 265–280. [Google Scholar] [CrossRef]
- Verdouw, H.; Van Echteld, C.; Dekkers, E. Ammonia determination based on indophenol formation with sodium salicylate. Water Res. 1978, 12, 399–402. [Google Scholar] [CrossRef]
- Cui, J.; Yang, H.; Yu, C.; Bai, S.; Wu, T.; Song, S.; Sun, W.; Shao, X.; Jiang, L. Effect of urea fertilization on biomass yield, chemical composition, in vitro rumen digestibility and fermentation characteristics of straw of highland barley planted in Tibet. J. Agric. Sci. 2016, 154, 151–164. [Google Scholar] [CrossRef]
- Groot, J.C.; Cone, J.W.; Williams, B.A.; Debersaques, F.M.; Lantinga, E.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Yang, H.; Tamminga, S.; Williams, B.A.; Dijkstra, J.; Boer, H. In vitro gas and volatile fatty acids production profiles of barley and maize and their soluble and washout fractions after feed processing. Anim. Feed Sci. Technol. 2005, 120, 125–140. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, S.; Si, J.; Zhang, J.; Gaowa, N.; Sun, X.; Lv, J.; Liu, G.; He, Y.; Wang, W. Effects of paper mulberry silage on the milk production, apparent digestibility, antioxidant capacity, and fecal bacteria composition in Holstein dairy cows. Animals 2020, 10, 1152. [Google Scholar] [CrossRef]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus subtilis improves immunity and disease resistance in rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Yu, Z. Effects of coconut and fish oils on ruminal methanogenesis, fermentation, and abundance and diversity of microbial populations in vitro. J. Dairy Sci. 2013, 96, 1782–1792. [Google Scholar] [CrossRef]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk fatty acids in dairy cows fed whole crude linseed, extruded linseed, or linseed oil, and their relationship with methane output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef]
- Ueda, K.; Ferlay, A.; Chabrot, J.; Loor, J.; Chilliard, Y.; Doreau, M. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage: Concentrate ratios. J. Dairy Sci. 2003, 86, 3999–4007. [Google Scholar] [CrossRef] [Green Version]
- Jian, G.; Wang, M.-Z.; Jing, Y.-J.; Sun, X.-Z.; Wu, T.-Y.; Shi, L.-F. Impacts of the unsaturation degree of long-chain fatty acids on the volatile fatty acid profiles of rumen microbial fermentation in goats in vitro. J. Integr. Agric. 2016, 15, 2827–2833. [Google Scholar]
- Pi, Y.; Ma, L.; Pierce, K.; Wang, H.; Xu, J.; Bu, D. Rubber seed oil and flaxseed oil supplementation alter digestion, ruminal fermentation and rumen fatty acid profile of dairy cows. Animal 2019, 13, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Scollan, N.D.; Dhanoa, M.; Choi, N.-J.; Maeng, W.; Enser, M.; Wood, J. Biohydrogenation and digestion of long chain fatty acids in steers fed on different sources of lipid. J. Agric. Sci. 2001, 136, 345–355. [Google Scholar] [CrossRef]
- Broudiscou, L.; Pochet, S.; Poncet, C. Effect of linseed oil supplementation on feed degradation and microbial synthesis in the rumen of ciliate-free and refaunated sheep. Anim. Feed Sci. Technol. 1994, 49, 189–202. [Google Scholar] [CrossRef]
- Van Gastelen, S.; Antunes-Fernandes, E.; Hettinga, K.; Klop, G.; Alferink, S.; Hendriks, W.; Dijkstra, J. Enteric methane production, rumen volatile fatty acid concentrations, and milk fatty acid composition in lactating Holstein-Friesian cows fed grass silage-or corn silage-based diets. J. Dairy Sci. 2015, 98, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchemin, K.; McGinn, S.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. In Annales de Zootechnie; EDP Sciences: Les Ulis, France, 2000; pp. 231–253. [Google Scholar]
- Mirzaei-Aghsaghali, A.; Maheri-Sis, N. Factors affecting mitigation of methane emission from ruminants: Microbiology and biotechnology strategies. J. Anim. Behav. Biometeorol. 2020, 4, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 839. [Google Scholar] [CrossRef] [Green Version]
- Sousa, D.Z.; Salvador, A.F.; Ramos, J.; Guedes, A.P.; Barbosa, S.N.; Stams, A.J.; Alves, M.M.; Pereira, M.A. Activity and viability of methanogens in anaerobic digestion of unsaturated and saturated long-chain fatty acids. Appl. Environ. Microbiol. 2013, 79, 4239–4245. [Google Scholar] [CrossRef] [Green Version]
- Belanche, A.; de la Fuente, G.; Newbold, C.J. Study of methanogen communities associated with different rumen protozoal populations. FEMS Microbiol. Ecol. 2014, 90, 663–677. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, J.Q.; Bu, D.P. Ruminal microbe of biohydrogenation of trans-vaccenic acid to stearic acid in vitro. BMC Res. Notes 2012, 5, 97. [Google Scholar] [CrossRef] [Green Version]
- Toral, P.G.; Hervás, G.; Carreño, D.; Leskinen, H.; Belenguer, A.; Shingfield, K.J.; Frutos, P. In vitro response to EPA, DPA, and DHA: Comparison of effects on ruminal fermentation and biohydrogenation of 18-carbon fatty acids in cows and ewes. J. Dairy Sci. 2017, 100, 6187–6198. [Google Scholar] [CrossRef]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Increased abundance of Sutterella spp. and Ruminococcus torques in feces of children with autism spectrum disorder. Mol. Autism 2013, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morissette, B.; Talbot, G.; Beaulieu, C.; Lessard, M. Growth performance of piglets during the first two weeks of lactation affects the development of the intestinal microbiota. J. Anim. Physiol. Anim. Nutr. 2018, 102, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.; López-García, A.; González-Recio, O.; Elcoso, G.; Fàbregas, F.; Chaucheyras-Durand, F.; Castex, M. Changes in the rumen and colon microbiota and effects of live yeast dietary supplementation during the transition from the dry period to lactation of dairy cows. J. Dairy Sci. 2019, 102, 6180–6198. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhu, Y.; Wang, Z.; Yu, X.; Hu, R.; Wang, X.; Cao, G.; Zou, H.; Shah, A.M.; Peng, Q. Comparing the Bacterial Community in the Gastrointestinal Tracts Between Growth-Retarded and Normal Yaks on the Qinghai–Tibetan Plateau. Front. Microbiol. 2020, 11, 3134. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | CON (DM %) | S (DM %) | F (DM %) |
|---|---|---|---|
| Oat hay | 41.80 | 41.49 | 41.87 |
| Corn silage | 29.58 | 27.97 | 28.23 |
| Corn fine | 10.63 | 10.93 | 9.23 |
| Soybean meal | 16.99 | 8.61 | 14.67 |
| Whole soybean | 0.00 | 10.00 | 0.00 |
| Flaxseed | 0.00 | 0.00 | 5.00 |
| Mineral vitamin premix | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrition (DM basis) | |||
| DM (kg) | 12.42 | 12.51 | 12.4 |
| CP (%) | 13.85 | 13.77 | 13.78 |
| NEL (Mcal/kg) | 1.48 | 1.49 | 1.49 |
| NFC (%) | 34.60 | 33.46 | 33.29 |
| NDF (%) | 42.00 | 41.95 | 42.41 |
| EE (%) | 2.60 | 4.01 | 4.01 |
| UFA (%) | 1.47 | 2.74 | 2.73 |
| n-6 (g) | 102.03 | 178.23 | 121.48 |
| n-3 (g) | 45.65 | 58.63 | 150.68 |
| n-6:n-3 | 2.23:1 | 3.04:1 | 0.8:1 |
| Item 1 | Diet 2 | Time 3 | SEM 4 | p-Value 5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | S | F | 3 h | 6 h | 12 h | 24 h | 48 h | Diet | Time | INT | ||
| Acetate (mmol/L) | 27.34 ab | 26.13 b | 27.67 a | 19.82 d | 24.67 c | 23.48 c | 30.17 b | 37.59 a | 6.67 | 0.024 | <0.001 | 0.149 |
| Propionate (mmol/L) | 10.68 | 10.64 | 11.09 | 7.54 d | 9.19 c | 9.04 c | 12.95 b | 15.89 a | 3.16 | 0.094 | <0.001 | 0.100 |
| Butyrate (mmol/L) | 4.21 ab | 4.13 b | 4.41 a | 2.72 d | 4.01 c | 3.96 c | 4.78 b | 6.01 a | 1.14 | 0.035 | <0.001 | 0.087 |
| A:P | 2.54 | 2.50 | 2.53 | 2.63 b | 2.69 a | 2.59 c | 2.34 d | 2.35 d | 0.16 | 0.592 | <0.001 | 0.973 |
| TVFA (mmol/L) | 44.12 ab | 42.28 b | 44.62 a | 30.88 d | 39.24 c | 37.85 c | 49.36 b | 62.03 a | 11.58 | 0.033 | <0.001 | 0.118 |
| NH3-N (mg/dL) | 10.84 | 11.85 | 11.84 | 5.04 e | 6.89 d | 8.11 c | 13.84 b | 23.43 a | 6.93 | 0.081 | <0.001 | 0.659 |
| MCP (μg/L) | 650.70 | 631.58 | 650.71 | 665.26 ab | 721.01 a | 644.74 b | 640.79 b | 549.03 c | 95.54 | 0.792 | <0.001 | 0.660 |
| Item 1 | Diet 2 | SEM 3 | p-Value | ||
|---|---|---|---|---|---|
| CON | S | F | |||
| GP48 (mL/g) | 67.48 b | 75.49 ab | 85.33 a | 13.26 | 0.032 |
| A (mL) | 69.36 b | 78.05 ab | 91.42 a | 15.97 | 0.024 |
| B (h) | 1.67 | 1.41 | 1.51 | 0.24 | 0.142 |
| C (h) | 6.48 | 7.09 | 7.69 | 1.23 | 0.194 |
| TRmaxG (h) | 2.61 | 2.38 | 1.92 | 0.69 | 0.157 |
| RmaxG (h) | 6.75 | 6.81 | 7.73 | 1.08 | 0.167 |
| TRmaxS (h) | 4.68 | 4.36 | 3.55 | 1.15 | 0.164 |
| RmaxS (mL/h) | 1.37 | 1.12 | 1.11 | 0.03 | 0.174 |
| Item | Diet (%) | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | S | F | |||
| Sutterella | 0.173 a | 0.107 b | 0.123 b | 0.033 | 0.007 |
| Prevotellaceae_Ga6A1 | 0.021 | 0.028 | 0.044 | 0.014 | 0.075 |
| Hydrogenoanaerobacterium | 0.006 b | 0.016 a | 0.011 ab | 0.005 | 0.029 |
| Butyrivibrio | 0.527 b | 0.678 a | 0.642 a | 0.078 | 0.015 |
| Succiniclasticum | 7.607 | 9.764 | 7.634 | 1.414 | 0.076 |
| Prevotellaceae_UCG-004 | 0.219 a | 0.325 ab | 0.356 a | 0.076 | 0.038 |
| Methanobrevibacter | 0.021 | 0.027 | 0.006 | 0.015 | 0.078 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.-G.; Wang, Y.; Xie, T.; Yang, Z.-T.; Wang, J.-D.; Zheng, Y.-H.; Guo, C.; Zhang, Y.; Wang, Q.-Q.; Wang, Z.-H.; et al. Effects of High-Forage Diets Containing Raw Flaxseeds or Soybean on In Vitro Ruminal Fermentation, Gas Emission, and Microbial Profile. Microorganisms 2021, 9, 2304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112304
Sun X-G, Wang Y, Xie T, Yang Z-T, Wang J-D, Zheng Y-H, Guo C, Zhang Y, Wang Q-Q, Wang Z-H, et al. Effects of High-Forage Diets Containing Raw Flaxseeds or Soybean on In Vitro Ruminal Fermentation, Gas Emission, and Microbial Profile. Microorganisms. 2021; 9(11):2304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112304
Chicago/Turabian StyleSun, Xiao-Ge, Yue Wang, Tian Xie, Zhan-Tao Yang, Ji-Dong Wang, Yu-Hui Zheng, Cheng Guo, Yan Zhang, Qian-Qian Wang, Zhong-Han Wang, and et al. 2021. "Effects of High-Forage Diets Containing Raw Flaxseeds or Soybean on In Vitro Ruminal Fermentation, Gas Emission, and Microbial Profile" Microorganisms 9, no. 11: 2304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112304