
Role of the Sortase A in the Release of Cell-Wall Proteinase PrtS in the Growth Medium of Streptococcus thermophilus 4F44

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. DNA Extraction, PCR Amplification, Electrophoresis, and Sequencing Conditions
2.3. Mutant Constructions and Natural Transformation
2.4. Detection of Extracellular Proteinase Activity
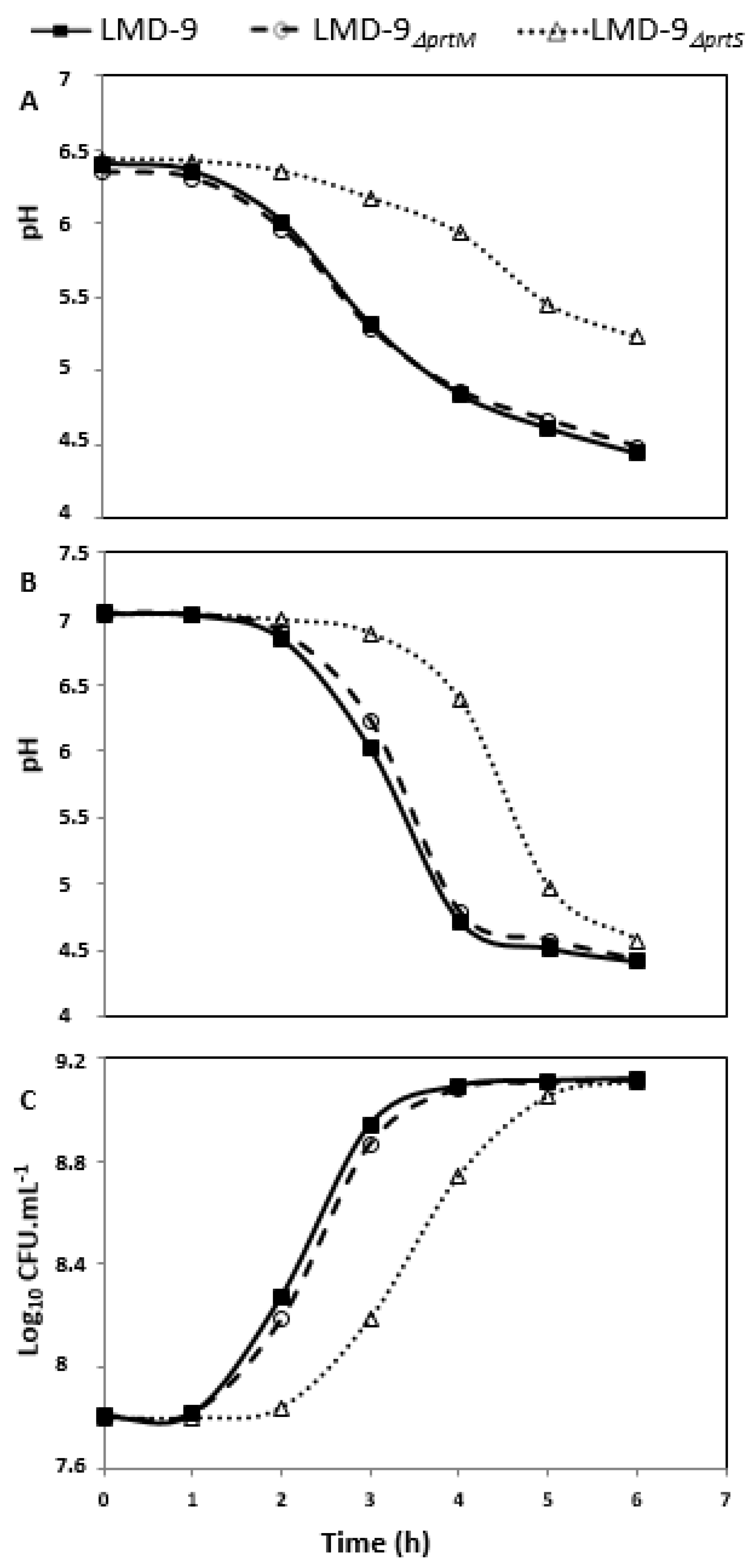
3. Results and Discussion
3.1. The Imperfect Duplication of 32 Amino Acid Residues in the Prodomain of PrtSLMD-9 Is Not Involved in the Anchoring of PrtS to the Cell Wall of S. thermophilus
3.2. Role of Maturases in the Maturation of PrtS in S. thermophilus LMD-9
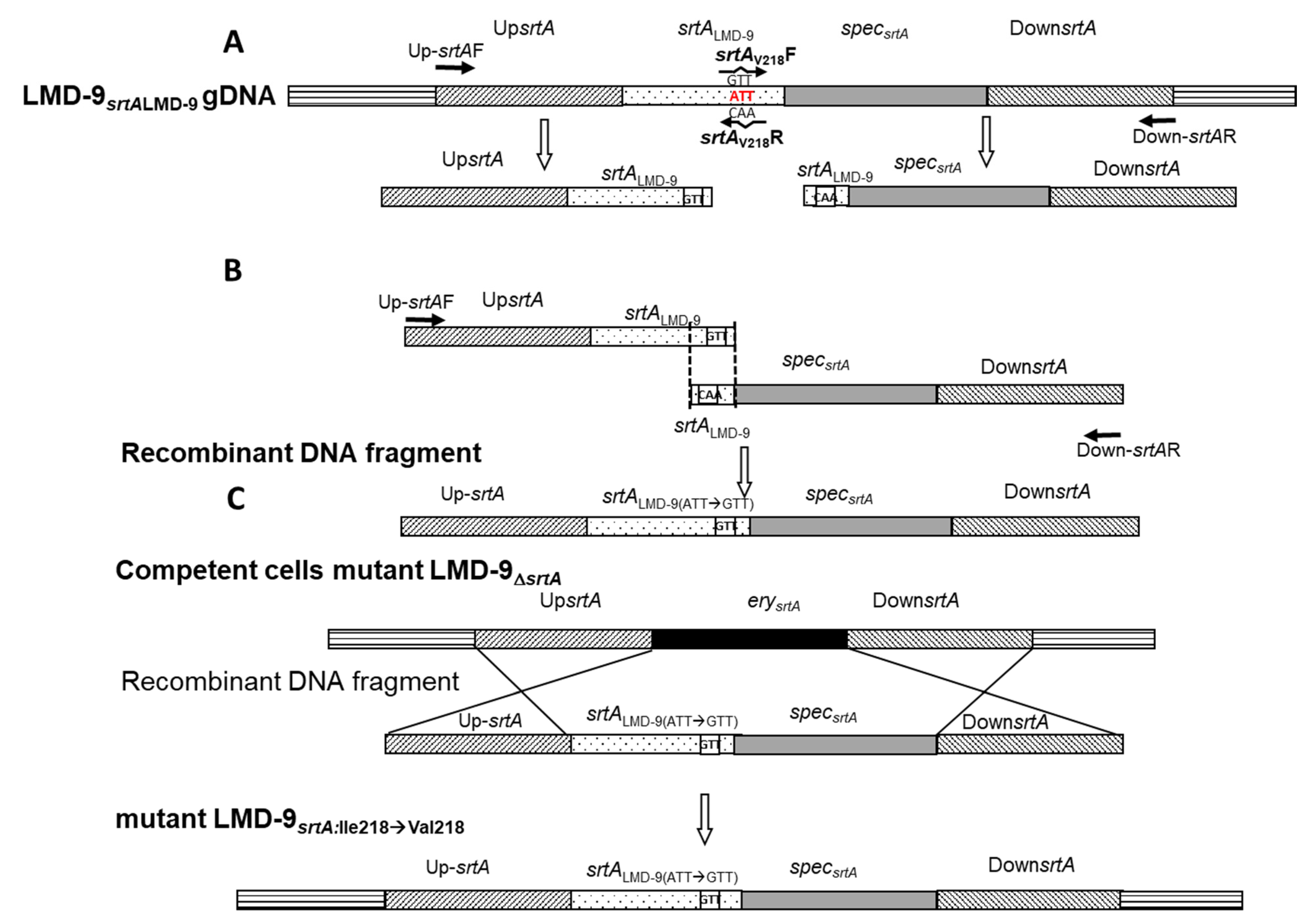
3.3. SrtA Is Responsible for the Anchoring of PrtS to the Cell Wall of S. thermophilus and Is Deficient in Strain 4F44
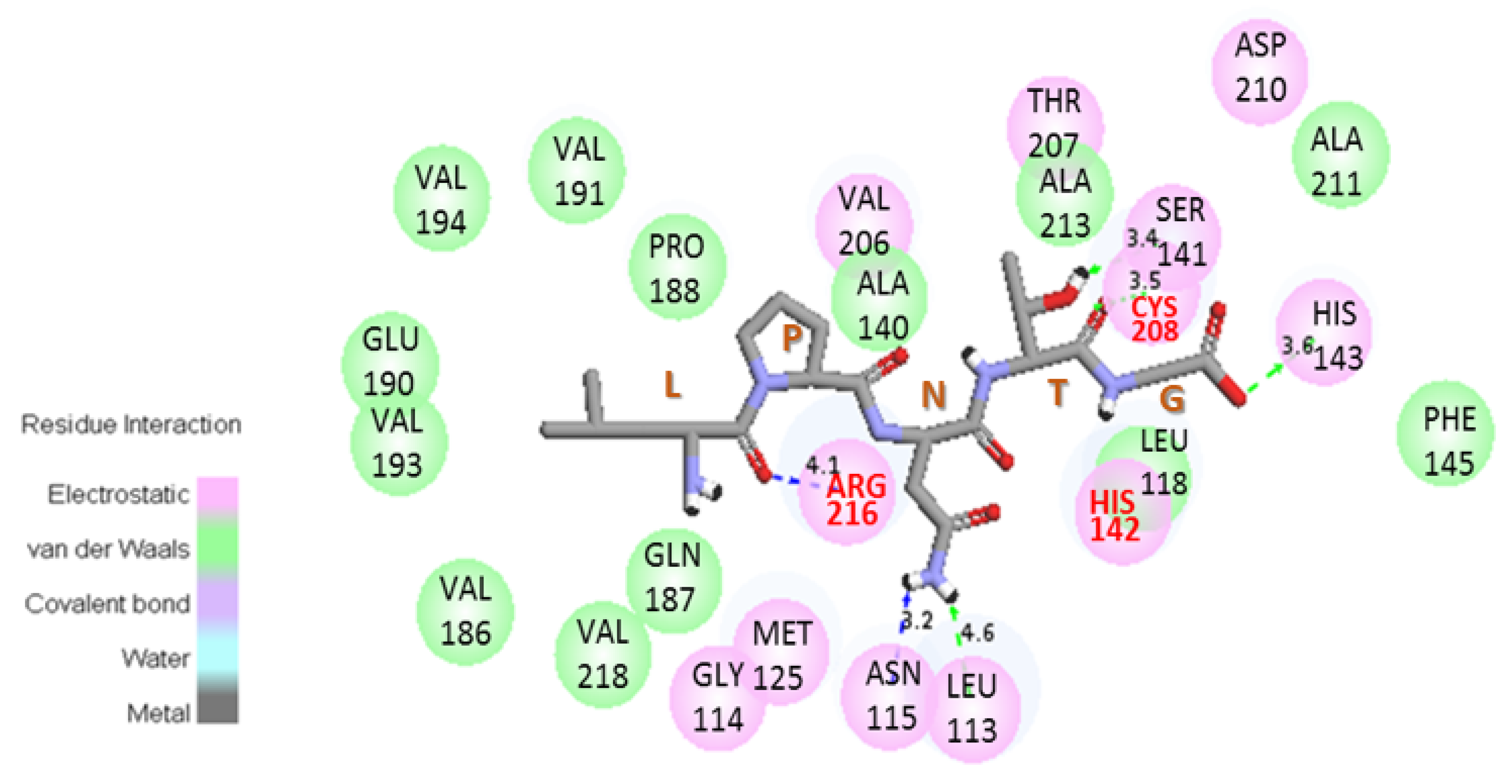
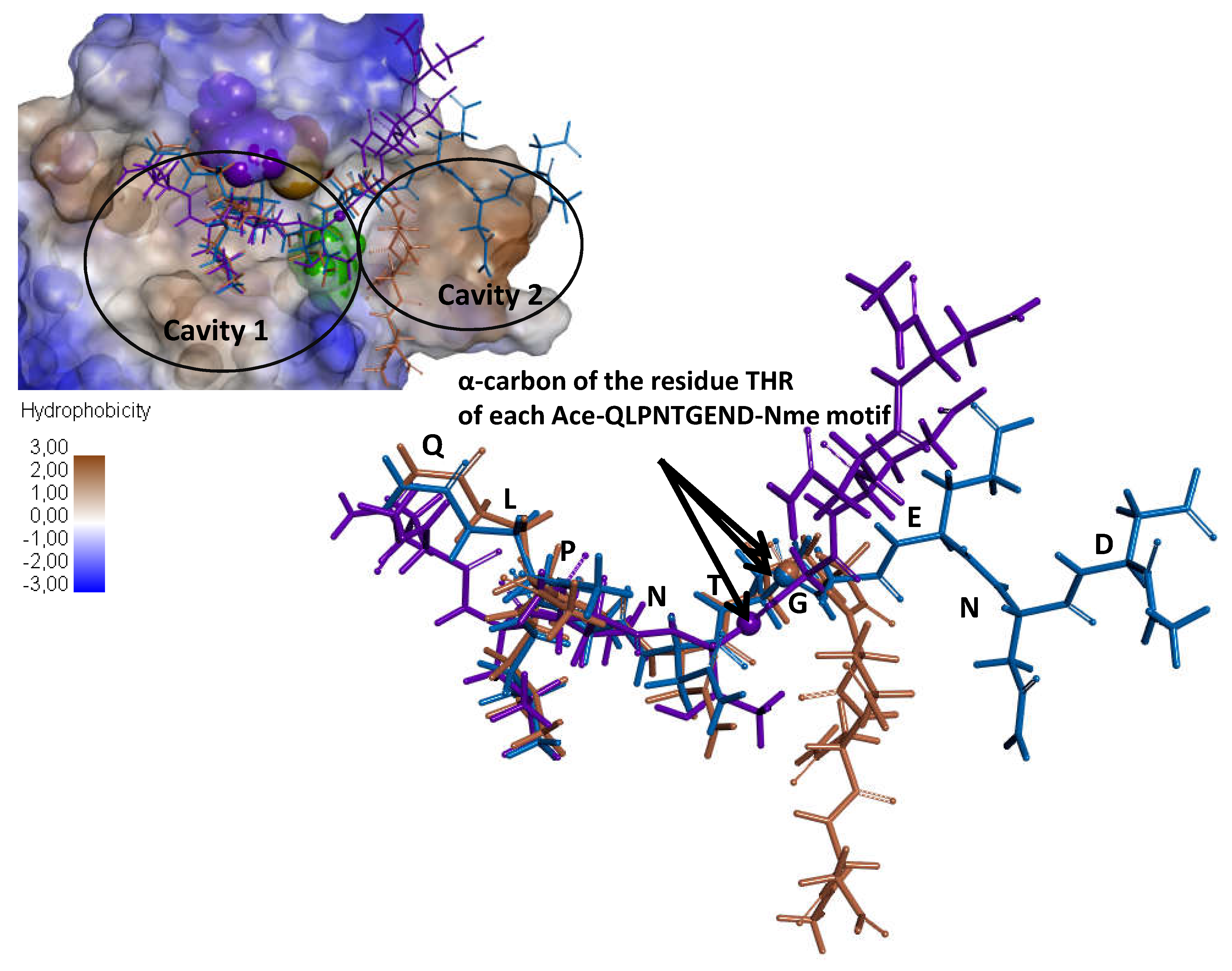
3.4. Substitution of the Ile 218 Residue Is Not Responsible for the Deficiency of SrtA4F44
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Cui, Y.; Xu, T.; Qu, X.; Hu, T.; Jiang, X.; Zhao, C. New insights into various production characteristics of Streptococcus thermophilus strains. Int. J. Mol. Sci. 2016, 17, 1701. [Google Scholar] [CrossRef]
- Jensen, A.; Valdórsson, O.; Frimodt-Møller, N.; Hollingshead, S.; Kilian, M. Commensal streptococci serve as a reservoir for β-lactam resistance genes in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2015, 59, 3529–3540. [Google Scholar] [CrossRef] [Green Version]
- Krzyściak, W.; Pluskwa, K.K.; Jurczak, A.; Kościelniak, D. The pathogenicity of the Streptococcus genus. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1361–1376. [Google Scholar] [CrossRef] [Green Version]
- Garault, P.; Letort, C.; Juillard, V.; Monnet, V. Branched-chain amino acid biosynthesis is essential for optimal growth of Streptococcus thermophilus in milk. Appl. Environ. Microbiol. 2000, 66, 5128–5133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastink, M.I.; Teusink, B.; Hols, P.; Visser, S.; de Vos, W.M.; Hugenholtz, J. Genome-scale model of Streptococcus thermophilus LMG18311 for metabolic comparison of lactic acid bacteria. Appl. Environ. Microbiol. 2009, 75, 3627–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandoy, D.; Fremaux, C.; Henry de Frahan, M.; Horvath, P.; Boyaval, P.; Hols, P.; Fontaine, L. The fast milk acidifying phenotype of Streptococcus thermophilus can be acquired by natural transformation of the genomic island encoding the cell-envelope proteinase PrtS. Microb. Cell Factories 2011, 10 (Suppl. S1), S21. [Google Scholar] [CrossRef] [Green Version]
- Galia, W.; Perrin, C.; Genay, M.; Dary, A. Variability and molecular typing of Streptococcus thermophilus strains displaying different proteolytic and acidifying properties. Int. Dairy J. 2009, 19, 89–95. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, G.M.; García-Garibay, J.M.; Cruz-Guerrero, A.E.; Gómez-Ruiz, L.; Ayala-Niño, A.; Castañeda-Ovando, A.; González-Olivares, L.G. Proteolytic system of Streptococcus thermophilus. J. Microbiol. Biotechnol. 2018, 28, 1581–1588. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Bonifait, L.; de la Cruz Dominguez-Punaro, M.; Vaillancourt, K.; Bart, C.; Slater, J.; Frenette, M.; Gottschalk, M.; Grenier, D. The cell envelope subtilisin-like proteinase is a virulence determinant for Streptococcus suis. BMC Microbiol. 2010, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Delorme, C.; Bartholini, C.; Bolotine, A.; Ehrlich, S.D.; Renault, P. Emergence of a cell wall protease in the Streptococcus thermophilus population. Appl. Environ. Microbiol. 2010, 76, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Espla, M.D.; Garault, P.; Monnet, V.; Rul, F. Streptococcus thermophilus cell wall-anchored proteinase: Release, purification, and biochemical and genetic characterization. Appl. Environ. Microbiol. 2000, 66, 4772–4778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, G.; Schmid, F.X. The mechanism of protein folding. Implications of in vitro refolding models for de novo protein folding and translocation in the cell. Biochemistry 1990, 29, 2205–2212. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Bryant, A.E.; Salmi, D.B.; Hayes-Schroer, S.M.; McIndoo, E.; Aldape, M.J.; Stevens, D.L. Identification and characterization of bicistronic speB and prsA gene expression in the Group A Streptococcus. J. Bacteriol. 2006, 188, 7626–7634. [Google Scholar] [CrossRef] [Green Version]
- Vos, P.; van Asseldonk, M.; van Jeveren, F.; Siezen, R.; Simons, G.; de Vos, W.M. A maturation protein is essential for production of active forms of Lactococcus lactis SK11 serine proteinase located in or secreted from the cell envelope. J. Bacteriol. 1989, 171, 2795–2802. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.H.; Nguyen, M.T.; Asis Das, A.; Ton-That, H. Anchoring surface proteins to the bacterial cell wall by sortase enzymes: How it started and what we know now. Curr. Opin. Microbiol. 2021, 60, 73–79. [Google Scholar] [CrossRef]
- Zrelovs, N.; Kurbatska, V.; Rudevica, V.; Leonchiks, A.; Fridmanis, D. Sorting out the superbugs: Potential of sortase A inhibitors among other antimicrobial strategies to tackle the problem of antibiotic resistance. Antibiotics 2021, 10, 164. [Google Scholar] [CrossRef]
- Freund, C.; Dirk Schwarzer, D. Engineered Sortases in Peptide and Protein Chemistry. ChemBioChem 2021, 22, 1347–1356. [Google Scholar] [CrossRef]
- Hu, P.; Bian, Z.; Fan, M.; Huang, M.; Zhang, P. Sec translocase and sortase A are colocalised in a locus in the cytoplasmic membrane of Streptococcus mutans. Arch. Oral Biol. 2008, 53, 150–154. [Google Scholar] [CrossRef]
- Kline, K.A.; Kau, A.L.; Chen, S.L.; Lim, A.; Pinkner, J.S.; Rosch, J.; Nallapareddy, S.R.; Murray, B.E.; Henriques-Normark, B.; Beatty, W.; et al. Mechanism for sortase localization and the role of sortase localization in efficient pilus assembly in Enterococcus faecalis. J. Bacteriol. 2009, 191, 3237–3247. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.N.; Aquilina, J.A.; Hains, P.G.; Henningham, A.; Sriprakash, K.S.; Caparon, M.G.; Nizet, V.; Kotb, M.; Cordwell, S.J.; Djordje-vic, S.P.; et al. Role of group A Streptococcus HtrA in the maturation of SpeB protease. Proteomics 2007, 7, 4488–4498. [Google Scholar] [CrossRef]
- Lyon, W.R.; Caparon, M.G. Trigger Factor-Mediated Prolyl Isomerization Influences Maturation of the Streptococcus pyogenes Cysteine Protease. J. Bacteriol. 2003, 185, 3661–3667. [Google Scholar] [CrossRef] [Green Version]
- Chang, O.K.; Perrin, C.; Galia, W.; Saulnier, F.; Miclo, L.; Roux, E.; Driou, A.; Humbert, G.; Dary, A. Release of the cell-envelope pro-tease PrtS in the growth medium of Streptococcus thermophilus 4F44. Int. Dairy J. 2012, 23, 91–98. [Google Scholar] [CrossRef]
- Race, P.R.; Bentley, M.L.; Melvin, J.A.; Crow, A.; Hughes, R.K.; Smith, W.D.; Sessions, R.B.; Kehoe, M.A.; McCafferty, D.G.; Ban-field, M.J. Crystal structure of Streptococcus pyogenes sortase A. implications for sortase mechanism. J. Biol. Chem. 2009, 284, 6924–6933. [Google Scholar] [CrossRef] [Green Version]
- Terzaghi, B.E.; Sandine, W.E. Improved Medium for Lactic Streptococci and Their Bacteriophages. Appl. Microbiol. 1975, 29, 807–813. [Google Scholar] [CrossRef]
- Gardan, R.; Besset, C.; Guillot, A.; Gitton, C.; Monnet, V. The oligopeptide transport system is essential for the development of natural competence in Streptococcus thermophilus strain LMD-9. J. Bacteriol. 2009, 191, 4647–4655. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G.; Decaris, B.; Leblond, P. Occurrence of deletions, associated with genetic instability in Streptomyces ambofaciens, is independent of the linearity of the chromosomal DNA. J. Bacteriol. 1997, 179, 4553–4558. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 1–3. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [Green Version]
- Lecomte, X.; Gagnaire, V.; Briard-Bion, V.; Jardin, J.; Lortal, S.; Dary, A.; Genay, M. The naturally competent strain Streptococcus thermophilus LMD-9 as a new tool to anchor heterologous proteins on the cell surface. Microb. Cell Factories 2014, 13, 82. [Google Scholar] [CrossRef] [Green Version]
- Maguin, E.; Prévost, H.; Ehrlich, S.D.; Gruss, A. Efficient insertional mutagenesis in lactococci and other gram-positive bacteria. J. Bacteriol. 1996, 178, 931–935. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Construction and characterization of Streptococcus suis–Escherichia coli shuttle cloning vectors. Plasmid 2001, 45, 101–113. [Google Scholar] [CrossRef]
- Diller, D.J.; Merz, K.M. High throughput docking for library design and library prioritization. Proteins 2001, 43, 113–124. [Google Scholar] [CrossRef]
- MacKerell, A.D.; Bashford, D.; Bellott, M.; Dunbrack, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]
- Vitikainen, M.; Lappalainen, I.; Seppala, R.; Antelmann, H.; Boer, H.; Taira, S.; Savilahti, H.; Hecker, M.; Vihinen, M.; Sarvas, M.; et al. Structure-function analysis of PrsA reveals roles for the parvulin-like and flanking N- and C-terminal domains in protein folding and se-cretion in Bacillus subtilis. J. Biol. Chem. 2004, 279, 19302–19314. [Google Scholar] [CrossRef] [Green Version]
- Germond, J.-E.; Delley, M.; Gilbert, C.; Atlan, D. Determination of the Domain of the Lactobacillus delbrueckii subsp. bulgaricus cell surface proteinase PrtB involved in attachment to the cell wall after heterologous expression of the prtB Gene in Lactococcus lactis. Appl. Environ. Microbiol. 2003, 69, 3377–3384. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Liu, G.; Ton-That, H.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef]
- Barnett, T.C.; Scott, J.R. Differential recognition of surface proteins in Streptococcus pyogenes by two sortase gene homologs. J. Bacteriol. 2002, 184, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Ton-That, H.; Schneewind, O. Sortase-catalysed anchoring of surface proteins to the cell wall of Staphylococcus aureus. Mol. Microbiol. 2001, 40, 1049–1057. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains\Plasmids | Origin | References | srtA Allele | Presence (+)/Absence (−) of the prtS Gene | PrtS Activity in mAU | |
|---|---|---|---|---|---|---|
| Cell Surface | Growth Supernatant | |||||
| Streptococcus thermophilus wild-type | ||||||
| LMD-9 | Yogurt | [30] | srtALMD-9 | + | 3110 | 0 |
| 4F44 | Cheese | [23] | srtA4F44 | + | 5620 | 3240 |
| Streptococcus thermophilus mutants | ||||||
| LMD-9srtALMD-9 | LMD-9 | Present study | srtALMD-9 | + | 3100 | 0 |
| LMD-9∆srtA | LMD-9 | Present study | Δ | + | 2600 | 1020 |
| LMD-9srtA4F44 | LMD-9 | Present study | srtA4F44 | + | 3124 | 1070 |
| LMD-9∆prtS | LMD-9 | [31] | srtALMD-9 | − | 0 | 0 |
| LMD-9prtS4F44 | LMD-9 | Present study | srtALMD-9 | + | 3093 | 0 |
| LMD-9∆prtM LMD-9srtA:Ile218→Val218 | LMD-9 LMD-9 | Present study Present study | srtALMD-9 srtA:Ile218→Val218 | + + | 3114 Present | 0 0 |
| Plasmids | Relevant markers and characteristics | |||||
| pG+host9 | - | [32] | ErmR *, pWV01 derivative, with thermoresistant replication function | |||
| pSET4s | - | [33] | SpecR *, replication function of Pg + host3 and pUC19 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awussi, A.A.; Roux, E.; Humeau, C.; Hafeez, Z.; Maigret, B.; Chang, O.K.; Lecomte, X.; Humbert, G.; Miclo, L.; Genay, M.; et al. Role of the Sortase A in the Release of Cell-Wall Proteinase PrtS in the Growth Medium of Streptococcus thermophilus 4F44. Microorganisms 2021, 9, 2380. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112380
Awussi AA, Roux E, Humeau C, Hafeez Z, Maigret B, Chang OK, Lecomte X, Humbert G, Miclo L, Genay M, et al. Role of the Sortase A in the Release of Cell-Wall Proteinase PrtS in the Growth Medium of Streptococcus thermophilus 4F44. Microorganisms. 2021; 9(11):2380. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112380
Chicago/Turabian StyleAwussi, Ahoefa Ablavi, Emeline Roux, Catherine Humeau, Zeeshan Hafeez, Bernard Maigret, Oun Ki Chang, Xavier Lecomte, Gérard Humbert, Laurent Miclo, Magali Genay, and et al. 2021. "Role of the Sortase A in the Release of Cell-Wall Proteinase PrtS in the Growth Medium of Streptococcus thermophilus 4F44" Microorganisms 9, no. 11: 2380. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112380