
Relationship between the Biofilm-Forming Capacity and Antimicrobial Resistance in Clinical Acinetobacter baumannii Isolates: Results from a Laboratory-Based In Vitro Study

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Isolates
2.2. Re-Identification of Isolates
2.3. Antimicrobial Susceptibility Testing and Resistotyping
2.4. Phenotypic Detection of Carbapenemase and Metallo-β-Lactamase Production
2.5. Phenotypic Detection of Bacterial Efflux Pumps Contributing to the MDR Phenotype
2.6. Biofilm Formation Assay
2.7. Statistical Analysis
2.8. Ethical Considerations
3. Results
3.1. Antibiotic Susceptibility of Isolates Included in the Study
3.2. Detection of Carbapenemase-Production and Efflux Pump-Overexpression Using Phenotypic Methods
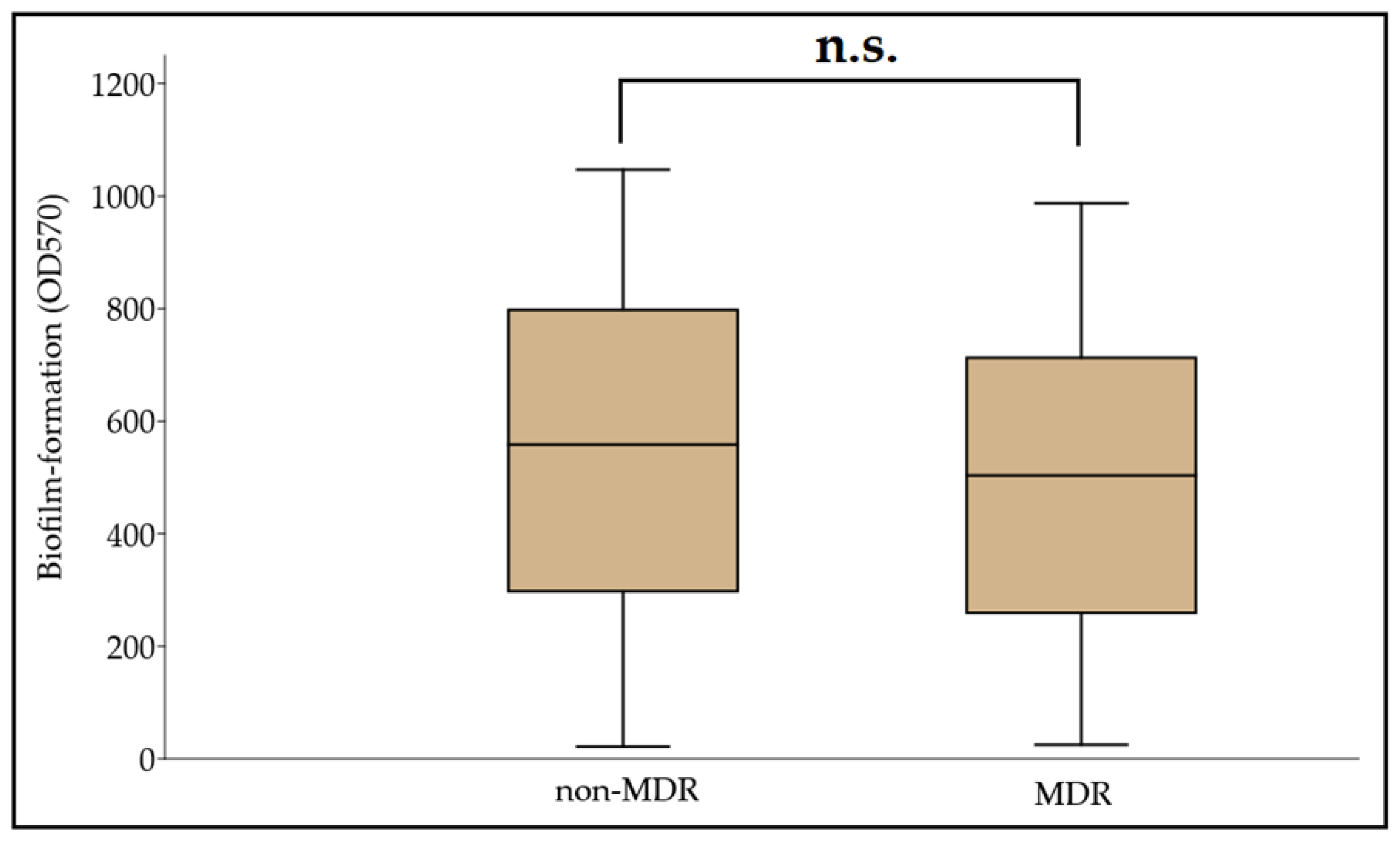
3.3. Biofilm Formation among MDR and Non-MDR A. baumannii Isolates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cannas, S.; Usai, D.; Pinna, A.; Benvenuti, S.; Tardugno, R.; Donadu, M.; Zanetti, S.; Kaliamurthy, J.; Molicotti, P. Essential oils in ocular pathology: An experimental study. J. Infect. Dev. Ctries 2015, 9, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.Z.; Zhanh, J.S.; Qiao, L. The Acinetobacter baumannii group: A systemic review. World J. Emerg. Med. 2013, 4, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Morris, F.C.; Dexter, C.; Kostoulias, X.; Uddin, M.I.; Peleg, A. The mechanisms of disease caused by Acinetobacter baumannii. Front. Microbiol. 2019, 10, e1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.H.; Lai, Y.R.; Chi, C.Y.; Ho, M.W.; Chen, C.L.; Liao, W.C.; Ho, C.H.; Chen, Y.A.; Chen, C.Y.; Lin, Y.T.; et al. Long-term surveillance of antibiotic prescriptions and the prevalence of antimicrobial resistance in non-fermenting Gram-negative bacilli. Microorganisms 2020, 8, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; Boscia, F.; Zanetti, S. In Vitro antimicrobial activity of a new ophthalmic solution containing Hexamidine Diisethionate 0.05% (Keratosept). Cornea 2020, 39, 1415–1418. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; McClean, S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines 2021, 9, 570. [Google Scholar] [CrossRef]
- Pascale, R.; Corcione, S.; Bussini, L.; Pancaldi, L.; Giacobbe, R.D.; Ambretti, S.; Lupia, T.; Costa, C.; Marchese, A.; De Rosa, F.G.; et al. Non-fermentative gram-negative bloodstream infection in northern Italy: A multicenter cohort study. BMC Infect. Dis. 2021, 21, e806. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.S.; Abidin, A.Z.; Liew, S.M.; Roberts, J.A.; Sime, F.B. The global prevalence of multidrug-resistance among Acinetobacter baumannii causing hospital-acquired and ventilator-associated pneumonia and its associated mortality: A systematic review and meta-analysis. J. Infect. 2019, 79, 593–600. [Google Scholar]
- Da Cunda, P.; Iribarnegaray, V.; Papa-Ezdra, R.; Bado, I.; González, M.J.; Zunino, P.; Vignoli, R.; Scavone, P. Characterization of the Different Stages of Biofilm Formation and Antibiotic Susceptibility in a Clinical Acinetobacter baumannii Strain. Microb. Drug Res. 2020, 26, 569–575. [Google Scholar] [CrossRef]
- Amin, M.; Navidifar, T.; Shooshtari, F.S.; Goodarzi, H. Association of the genes encoding Metallo-β-Lactamase with the presence of integrons among multidrug-resistant clinical isolates of Acinetobacter baumannii. Infect. Drug Res. 2019, 12, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- World Health Organisation (WHO). Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organisation: Geneva, Switzerland, 2017; pp. 1–7. [Google Scholar]
- Maurice, N.M.; Bedi, B.; Sadikot, R.T. Pseudomonas aeruginosa Biofilms: Host Response and Clinical Implications in Lung Infections. Am. J. Respir. Cell Mol. Biol. 2018, 58, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Cifou, O.; Rojo-Molinero, E.; Macia, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. APMIS 2017, 125, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Scribano, D.; Sarshar, M.; Di Bonaventura, G.; Palamara, A.T.; Ambrosi, C. Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter baumannii Way. Microorganisms 2021, 9, 1353. [Google Scholar] [CrossRef] [PubMed]
- Tahaei, S.A.S.; Stájer, A.; Barrak, I.; Ostorházi, E.; Szabó, D.; Gajdács, M. Correlation Between Biofilm-Formation and the Antibiotic Resistant Phenotype in Staphylococcus aureus Isolates: A Laboratory-Based Study in Hungary and a Review of the Literature. Infect. Drug Res. 2021, 14, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Cepas, V.; López, Y.; Munoz, E.; Rolo, D.; Ardanuy, C.; Martí, M.; Xercavins, M.; Horcajada, J.P.; Bosch, J.; Soto, S.M. Relationship Between Biofilm Formation and Antimicrobial Resistance in Gram-Negative Bacteria. Microb. Drug Res. 2019, 25, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila-Novoa, M.G.; Solís-Velázquez, O.A.; Rangel-López, D.E.; González-Gómez, J.P.; Guerrero-Medin, P.J.; Gutiérez-Lomelí, M. Biofilm Formation and Detection of Fluoroquinolone- and Carbapenem-Resistant Genes in Multidrug-Resistant Acinetobacter baumannii. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, e3454907. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.Y.; Baek, W.K.; Kim, H.A. Association of biofilm production with colonization among clinical isolates of Acinetobacter baumannii. Korean J. Intern. Med. 2017, 32, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Gajdács, M.; Bátori, Z.; Ábrók, M.; Lázár, A.; Burián, K. Characterization of Resistance in Gram-Negative Urinary Isolates Using Existing and Novel Indicators of Clinical Relevance: A 10-Year Data Analysis. Life 2020, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.; Kostrzewa, M. MALDI-TOF MS in the microbiology laboratory: Current trends. Curr. Issues Mol. Biol. 2017, 23, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadat, A.; El-Sherbiny, H.; Zakaria, A.; Ramadan, H.; Awad, A. Prevalence, antibiogram and virulence characterization of Vibrio isolates from fish and shellfish in Egypt: A possible zoonotic hazard to humans. J. Appl. Microbiol. 2021, 131, 485–498. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical Breakpoints and Dosing. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 5 August 2021).
- Navidifar, T.; Amin, M.; Rashno, M. Effects of sub-inhibitory concentrations of meropenem and tigecycline on the expression of genes regulating pili, efflux pumps and virulence factors involved in biofilm formation by Acinetobacter baumannii. Infect. Drug Res. 2019, 12, 1099–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Paterson, D.L. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Kichibiro, T.; Komatsu, M.; Yamasaki, K.; Nakamura, T.; Nishio, H.; Kimura, K.; Niki, M.; Kida, N.; Ohama, M.; Fukuda, N.; et al. Comparison of the performance of three carbapenem inactivation methods for the detection of carbapenemase-producing Gram-negative bacilli. J. Infect. Chemother. 2021, 27, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Abouelfetouh, A.; Torky, A.S.; Aboulmagd, E. Phenotypic and genotypic characterization of carbapenem-resistant Acinetobacter baumannii isolates from Egypt. Antimicrob. Res. Infect. Control 2019, 8, e185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Ghalavand, Z.; Yadegar, A.; Eslami, G. Characteristics and diversity of mutations in regulatory genes of resistance-nodulation-cell division efflux pumps in association with drug-resistant clinical isolates of Acinetobacter baumannii. Antimicrob. Res. Infect. Control 2021, 10, e53. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Vivas, J.; Chapartegui-González, I.; Fernández-Martínez, M.; González-Rico, C.; Fortún, J.; Escudero, R.; Marco, F.; Linares, L.; Montejo, M.; Aranzamendi, M.; et al. Biofilm formation by multidrug resistant Enterobacteriaceae strains isolated from solid organ transplant recipients. Sci. Rep. 2019, 9, e8928. [Google Scholar] [CrossRef] [Green Version]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukovic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New treatment options against carbapenem-resistant Acinetobacter baumannii infections. Antimicrob. Agents Chemother. 2019, 63, e01110–e01118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarashi, S.; Goodarzi, H.; Erfanimanesh, S.; Pormohammad, A.; Hashemi, A. Phenotypic and molecular detection of metallo-beta-lactamase genes among imipenem resistant Pseudomonas aeruginosa and Acinetobacter baumannii strains isolated from patients with burn injuries. Arch. Clin. Infect. Dis. 2016, 11, e39036. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. Epidemiology and diagnostics of carbapenem resistance in Gram-negative bacteria. Clin. Infect. Dis. 2019, 69, S521–S528. [Google Scholar] [CrossRef] [Green Version]
- Gheorghe, I.; Barbu, I.C.; Surleac, M.; Sarbu, I.; Popa, L.I.; Paraschiv, S.; Feng, Y.; Lazar, V.; Chifiriuc, M.C.; Otelea, D.; et al. Subtypes, resistance and virulence platforms in extended-drug resistant Acinetobacter baumannii Romanian isolates. Sci. Rep. 2021, 11, e13288. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Isler, B.; Stewart, A. New treatment options for multiresistant Gram-negatives. Curr. Opin. Infect. Dis. 2020, 33, 214–223. [Google Scholar] [CrossRef]
- Banafsheh, D.M.; Hesan, A.; Hossein, K.; Afsaneh, K. Antimicrobial categories in describing multidrug resistance, extensive drug resistance and pan-drug resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: A systematic review. Rev. Med. Microbiol. 2021, 32, 6–11. [Google Scholar]
- Rangel, R.; Chagas, T.P.G.; De-Simone, S.G. Acinetobacter baumannii Infections in Times of COVID-19 Pandemic. Pathogens 2021, 10, 1006. [Google Scholar] [CrossRef]
- Sarshar, M.; Behzadi, P.; Scribano, D.; Palama, A.T.; Ambrosi, C. Acinetobacter baumannii: An Ancient Commensal with Weapons of a Pathogen. Pathogens 2021, 10, 387. [Google Scholar] [CrossRef] [PubMed]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect. Dis. 2019, 19, e629. [Google Scholar] [CrossRef] [Green Version]
- Hassan, P.A.; Kihder, A.K. Correlation of biofilm formation and antibiotic resistance among clinical and soil isolates of Acinetobacter baumannii in Iraq. Acta Microbiol. Immunol. Hung. 2020, 67, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, e483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sittihisak, S. Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar] [PubMed]
- Greene, C.; Vadlamudi, G.; Newton, D.; Foxman, B.; Xi, C. The influence of biofilm formation and multidrug resistance on environmental survival of clinical and environmental isolates of Acinetobacter baumannii. Am. J. Infect. Control 2016, 44, e65–e71. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Muras, A.; Parga, A.; Romero, M.; Rumbo-Feal, S.; Poza, M.; Ramos-Vivas, J.; Otero, A. Controlling Surface Associated Motility and Biofilm Formation in Acinetobacter baumannii ATCC-17978TM. Front. Microbiol. 2020, 11, e565548. [Google Scholar] [CrossRef]
- Selasi, G.N.; Nicholas, A.; Jeon, H.; Na, S.H.; Kwon, H.; Kim, J.Y.; Heo, S.T.; Oh, H.M.; Lee, J.C. Differences in biofilm mass, expression of biofilm-associated genes, and resistance to desiccation between epidemic and sporadic clones of carbapenem-resistant Acinetobacter baumannii sequence type 191. PLoS ONE 2016, 11, e0162576. [Google Scholar] [CrossRef] [PubMed]
- Azizi, O.; Shahcheraghi, F.; Salimizand, H.; Modarresi, F.; Shakibaie, M.R.; Mansouri, S.; Ramazanzadeh, R.; Badmasti, F.; Nikbin, V. Molecular Analysis and Expression of bap Gene in Biofilm-Forming Multi-Drug-Resistant Acinetobacter baumannii. Rep. Biochem. Mol. Biol. 2016, 5, 62–72. [Google Scholar] [PubMed]
- Gallant, C.V.; Daniels, C.; Leung, J.M.; Ghosh, A.S.; Young, K.D.; Kotra, L.P.; Burrows, L.L. Common β-lactamases inhibit bacterial biofilm formation. Mol. Microbiol. 2005, 58, 1012–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Resistotypes | Resistance Pattern | MAR Index | MDR Status * | Ratio of Isolates (n, %) |
|---|---|---|---|---|
| I | CIP | 0.111 | non-MDR (57.28%; n = 177) | 9 (2.91%) |
| II | LEV | 0.111 | 10 (3.24%) | |
| III | SXT | 0.111 | 20 (6.47%) | |
| IV | GEN, SXT | 0.222 | 28 (9.06%) | |
| V | CIP, SXT | 0.222 | 3 (0.97%) | |
| VI | CIP, LEV | 0.222 | 14 (4.53%) | |
| VII | IMI, MER, SXT | 0.333 | 12 (3.88%) | |
| VIII | IMI, MER, GEN | 0.333 | 18 (5.83%) | |
| IX | CIP, LEV, SXT | 0.333 | 18 (5.83%) | |
| X | CIP, LEV, MER | 0.333 | 6 (1.94%) | |
| XI | CIP, LEV, IMI | 0.333 | 10 (3.24%) | |
| XII | CIP, LEV, GEN | 0.333 | 9 (2.91%) | |
| XIII | CIP, LEV, IMI, MER | 0.444 | 20 (6.47%) | |
| XIV | CIP, LEV, GEN, AMI, SXT | 0.555 | MDR (42.72%; n = 132) | 29 (9.40%) |
| XV | CIP, LEV, GEN, AMI, SXT, TIG | 0.666 | 20 (6.47%) | |
| XVI | CIP, LEV, IMI, MER, GEN, SXT | 0.666 | 12 (3.88%) | |
| XVII | CIP, LEV, IMI, MER, GEN, TIG | 0.666 | 10 (3.24%) | |
| XVIII | CIP, LEV, IMI, MER, GEN, AMI, SXT | 0.777 | 47 (15.21%) | |
| XIX | CIP, LEV, IMI, MER, GEN, AMI, SXT, TIG | 0.888 | 7 (2.27%) | |
| XX | CIP, LEV, IMI, MER, GEN, AMI, SXT, COL | 0.888 | 5 (1.62%) | |
| XXI | CIP, LEV, IMI, MER, GEN, AMI, SXT, TIG, COL | 1.000 | 2 (0.65%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donadu, M.G.; Mazzarello, V.; Cappuccinelli, P.; Zanetti, S.; Madléna, M.; Nagy, Á.L.; Stájer, A.; Burián, K.; Gajdács, M. Relationship between the Biofilm-Forming Capacity and Antimicrobial Resistance in Clinical Acinetobacter baumannii Isolates: Results from a Laboratory-Based In Vitro Study. Microorganisms 2021, 9, 2384. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112384
Donadu MG, Mazzarello V, Cappuccinelli P, Zanetti S, Madléna M, Nagy ÁL, Stájer A, Burián K, Gajdács M. Relationship between the Biofilm-Forming Capacity and Antimicrobial Resistance in Clinical Acinetobacter baumannii Isolates: Results from a Laboratory-Based In Vitro Study. Microorganisms. 2021; 9(11):2384. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112384
Chicago/Turabian StyleDonadu, Matthew Gavino, Vittorio Mazzarello, Piero Cappuccinelli, Stefania Zanetti, Melinda Madléna, Ádám László Nagy, Anette Stájer, Katalin Burián, and Márió Gajdács. 2021. "Relationship between the Biofilm-Forming Capacity and Antimicrobial Resistance in Clinical Acinetobacter baumannii Isolates: Results from a Laboratory-Based In Vitro Study" Microorganisms 9, no. 11: 2384. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112384