
Susceptibility to Bismuth(III) of Aquaculture Bacterial Pathogens: Effectiveness of Bismuth–Deferiprone Therapy against Vibrio anguillarum Infection in Fish


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Routine Growth Conditions
2.2. Preparation of Bismuth Stock Solution
2.3. Test for Inhibition of Bacterial Growth
2.4. Evaluation of Bismuth–Deferiprone against V. anguillarum in Experimental Infections
3. Results
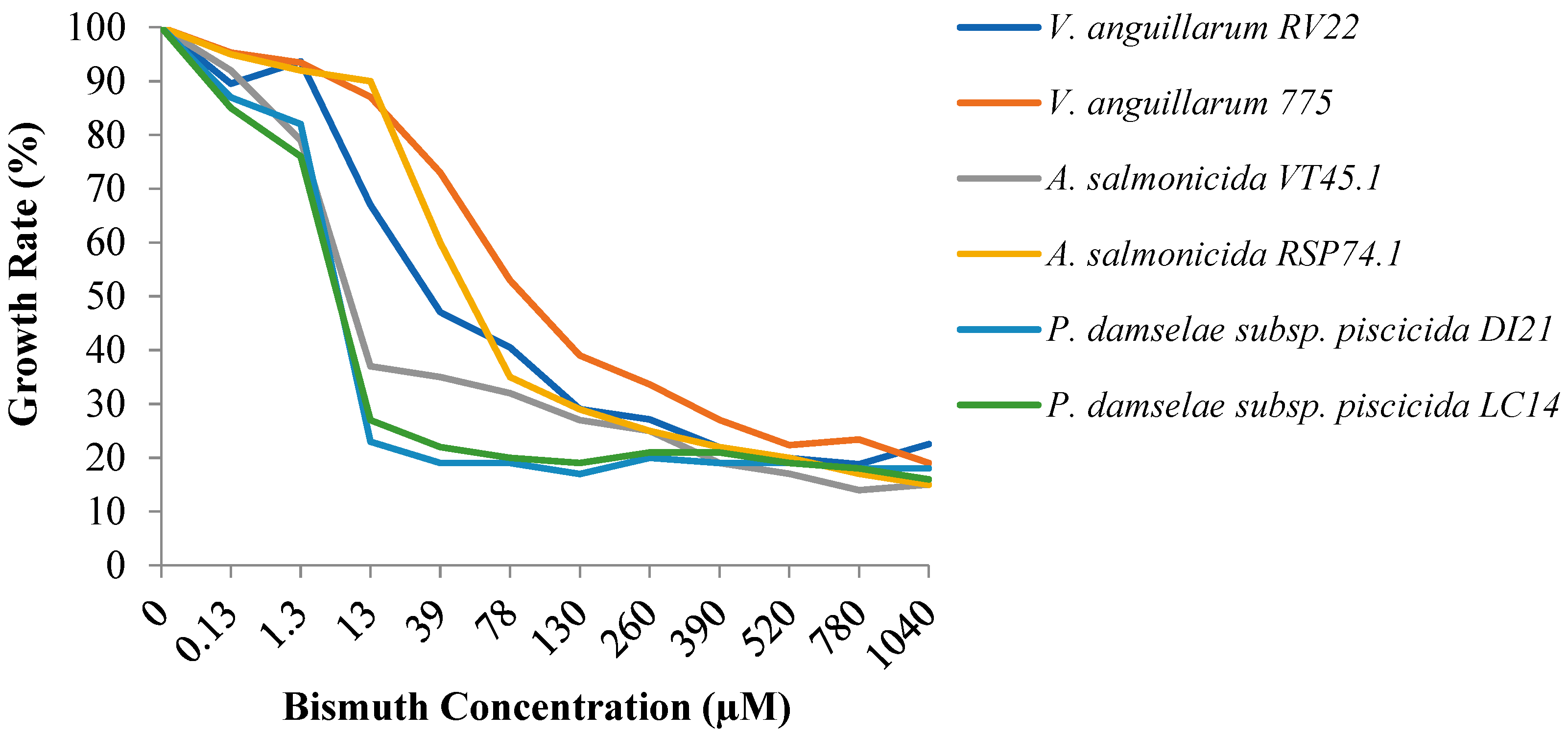
3.1. Susceptibility to Bismuth of a Collection of Aquaculture Bacterial Pathogens
3.2. Effect of Siderophore Production to Bismuth Susceptibility
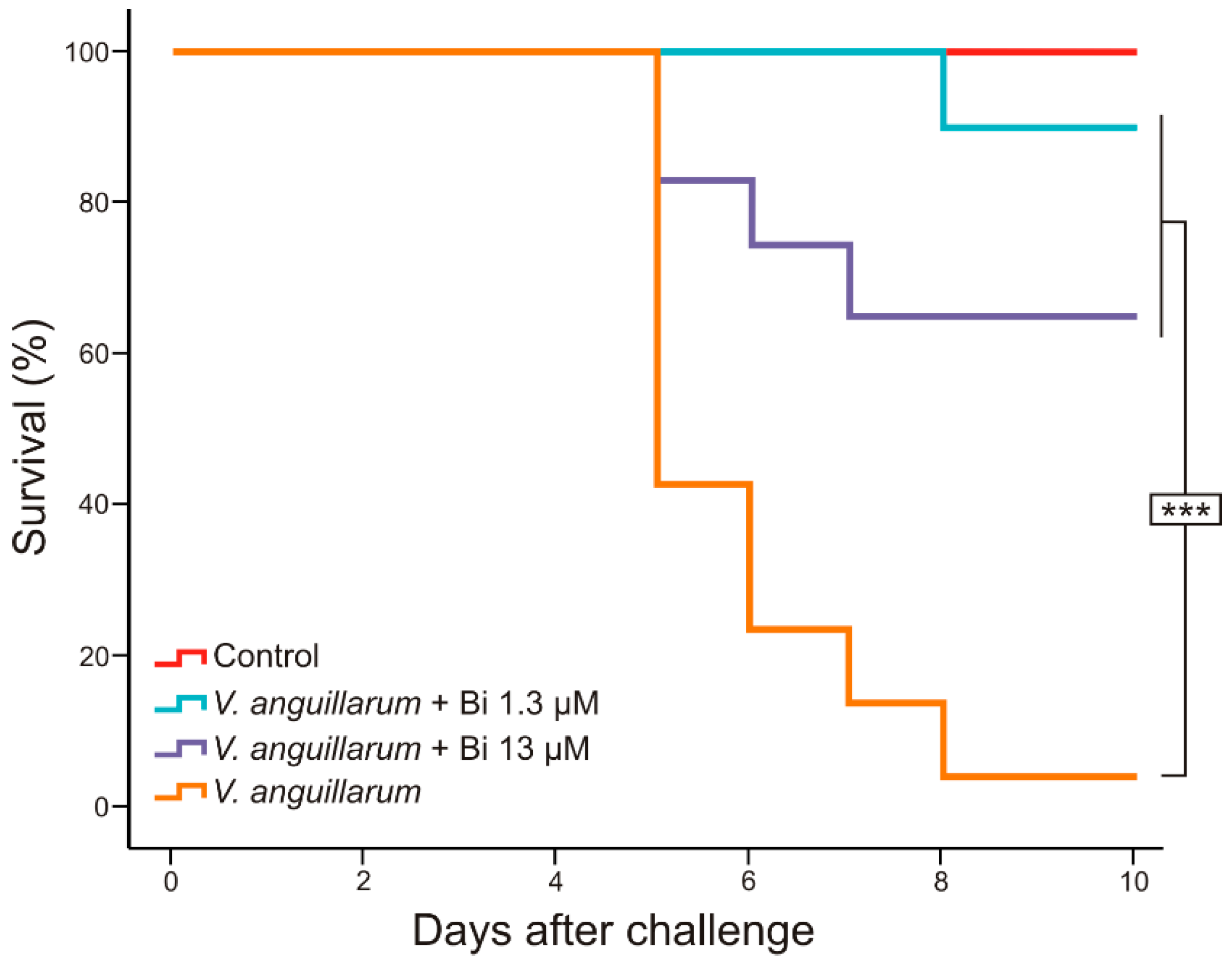
3.3. Usefulness of Deferiprone–Bismuth to Treat V. anguillarum Infections
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture (SOFIA) 2018—Meeting the Sustainable Development Goals; Food and Agricultural Organization: Rome, Italy, 2018. [Google Scholar]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Gudding, R. Vaccination as a preventive measure. In Fish Vaccination; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 12–21. [Google Scholar]
- Chen, L.; Alali, W. Editorial: Recent discoveries in human serious foodborne pathogenic bacteria: Resurgence, pathogenesis, and control strategies. Front. Microbiol. 2018, 9, 2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffin, R.N.; Werrett, M.V.; Andrews, P.C. Antimony and bismuth as antimicrobial agents. Adv. Inorg. Chem. 2020, 75, 207–255. [Google Scholar] [CrossRef]
- Thomas, F.; Diaz-Bone, R.A.; Wuerfel, O.; Huber, B.; Weidenbach, K.; Schmitz, R.A.; Hensel, R. Connection between multimetal(loid) methylation in methanoarchaea and central intermediates of methanogenesis. Appl. Environ. Microbiol. 2011, 77, 8669–8675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, S.D.; Walker, R.J.; Jones, H.; Rhodes, J.M. Rectal bismuth subsalicylate as therapy for ulcerative colitis. Aliment. Pharmacol. Ther. 2007, 4, 333–338. [Google Scholar] [CrossRef]
- Thazhath, S.S.; Haque, M.; Florin, T.H. Oral bismuth for chronic intractable diarrheal conditions? Clin. Exp. Gastroenterol. 2013, 6, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debraekeleer, A.; Remaut, H. Future perspective for potential Helicobacter pylori eradication therapies. Future Microbiol. 2018, 13, 671–687. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, C.S.; Marshall, B.J.; Blincow, E.D.; Wilson, D.H.; Blackbourn, S.; Phillips, M. Prevention of nitroimidazole resistance in Campylobacter pylori by coadministration of colloidal bismuth subcitrate: Clinical and in vitro studies. J. Clin. Pathol. 1988, 41, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Midolo, P.D.; Lambert, J.R.; Kerr, T.G.; Tee, W. In vitro synergy between ranitidine bismuth citrate and tetracycline or clarithromycin against resistant strains of Helicobacter pylori. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 832–834. [Google Scholar] [CrossRef]
- Andersen, L.P.; Colding, H.; Kristiansen, J.E. Potentiation of the action of metronidazole on Helicobacter pylori by omeprazole and bismuth subcitrate. Int. J. Antimicrob. Agents 2000, 14, 231–234. [Google Scholar] [CrossRef]
- Pindling, S.; Azulai, D.; Zheng, B.; Dahan, D.; Perron, G.G. Dysbiosis and early mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS Microbiol. Lett. 2018, 365, 188. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.L.; Lyle, D.A.; Ritz, N.L.; Granat, A.S.; Khurshid, A.N.; Kherbik, N.; Hider, R.; Lin, H.C. Bismuth(III) deferiprone effectively inhibits growth of Desulfovibrio desulfuricans ATCC 27774. BioMetals 2016, 29, 311–319. [Google Scholar] [CrossRef]
- Balado, M.; Lages, M.A.; Fuentes-Monteverde, J.C.; Martínez-Matamoros, D.; Rodríguez, J.; Jiménez, C.; Lemos, M.L. The siderophore piscibactin is a relevant virulence factor for Vibrio anguillarum favored at low temperatures. Front. Microbiol. 2018, 9, 1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thode, S.K.; Rojek, E.; Kozlowski, M.; Ahmad, R.; Haugen, P. Distribution of siderophore gene systems on a Vibrionaceae phylogeny: Database searches, phylogenetic analyses and evolutionary perspectives. PLoS ONE 2018, 13, e0191860. [Google Scholar] [CrossRef] [Green Version]
- Souto, A.; Montaos, M.A.M.A.; Rivas, A.J.A.J.; Balado, M.; Osorio, C.R.; Rodríguez, J.; Lemos, M.L.M.L.; Jiménez, C. Structure and biosynthetic assembly of piscibactin, a siderophore from Photobacterium damselae subsp. piscicida, predicted from genome analysis. Eur. J. Org. Chem. 2012, 2012, 5693–5700. [Google Scholar] [CrossRef]
- Osorio, C.R.; Juiz-Rio, S.; Lemos, M.L. A siderophore biosynthesis gene cluster from the fish pathogen Photobacterium damselae subsp. piscicida is structurally and functionally related to the Yersinia high-pathogenicity island. Microbiology 2006, 152, 3327–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balado, M.; Souto, A.; Vences, A.; Careaga, V.P.; Valderrama, K.; Segade, Y.; Rodríguez, J.; Osorio, C.R.; Jiménez, C.; Lemos, M.L. Two catechol siderophores, acinetobactin and amonabactin, are simultaneously produced by Aeromonas salmonicida subsp. salmonicida sharing part of the biosynthetic pathway. ACS Chem. Biol. 2015, 10, 2850–2860. [Google Scholar] [CrossRef] [PubMed]
- Klase, G.; Lee, S.; Liang, S.; Kim, J.; Zo, Y.-G.; Lee, J. The microbiome and antibiotic resistance in integrated fishfarm water: Implications of environmental public health. Sci. Total Environ. 2019, 649, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef] [PubMed]
- Sadler, P.J.; Li, H.; Sun, H. Coordination chemistry of metals in medicine: Target sites for bismuth. Coord. Chem. Rev. 1999, 185–186, 689–709. [Google Scholar] [CrossRef]
- Luqman, A.; Blair, V.L.; Brammananth, R.; Crellin, P.K.; Coppel, R.L.; Andrews, P.C. Powerful antibacterial activity of phenyl-thiolatobismuth(III) complexes derived from oxadiazolethiones. Eur. J. Inorg. Chem. 2015, 2015, 4935–4945. [Google Scholar] [CrossRef]
- Ge, R.; Sun, H. Bioinorganic chemistry of bismuth and antimony: Target sites of metallodrugs. Acc. Chem. Res. 2007, 40, 267–274. [Google Scholar] [CrossRef]
- Bland, M.V.; Ismail, S.; Heinemann, J.A.; Keenan, J.I. The action of bismuth against Helicobacter pylori mimics but is not caused by intracellular iron deprivation. Antimicrob. Agents Chemother. 2004, 48, 1983–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sox, T.E.; Olson, C.A. Binding and killing of bacteria by bismuth subsalicylate. Antimicrob. Agents Chemother. 1989, 33, 2075–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, L.L.; Granat, A.S.; Lee, S.; Xu, H.; Ritz, N.L.; Hider, R.; Lin, H.C. Bismuth(III) interactions with Desulfovibrio desulfuricans: Inhibition of cell energetics and nanocrystal formation of Bi2S3 and Bi0. BioMetals 2019, 32, 803–811. [Google Scholar] [CrossRef]
- Beil, W.; Birkholz, C.; Wagner, S.; Sewing, K.-F. Bismuth subcitrate and omeprazole inhibit Helicobacter pylori F1-ATPase. Pharmacology 1995, 50, 333–337. [Google Scholar] [CrossRef]
- Domenico, P.; Reich, J.; Madonia, W.; Cunha, B.A. Resistance to bismuth among gram-negative bacteria is dependent upon iron and its uptake. J. Antimicrob. Chemother. 1996, 38, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Kim, N.; Roh, H.J.; Chun, W.K.; Ho, D.T.; Lee, Y.; Kim, D.H. Administration of antibiotics can cause dysbiosis in fish gut. Aquaculture 2019, 512, 734330. [Google Scholar] [CrossRef]
- Sallum, U.W.; Chen, T.T. Inducible resistance of fish bacterial pathogens to the antimicrobial peptide cecropin B. Antimicrob. Agents Chemother. 2008, 52, 3006–3012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Antimicrobial resistance in bacteria isolated from aquaculture sources in Australia. J. Appl. Microbiol. 2006, 100, 1103–1113. [Google Scholar] [CrossRef]
- Dubert, J.; Osorio, C.R.; Prado, S.; Barja, J.L. Persistence of antibiotic resistant Vibrio spp. in shellfish hatchery environment. Microb. Ecol. 2016, 72, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Popowska, M. Insight into the mobilome of Aeromonas strains. Front. Microbiol. 2015, 6, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trudel, M.V.; Vincent, A.T.; Attéré, S.A.; Labbé, M.; Derome, N.; Culley, A.I.; Charette, S.J. Diversity of antibiotic-resistance genes in Canadian isolates of Aeromonas salmonicida subsp. salmonicida: Dominance of pSN254b and discovery of pAsa8. Sci. Rep. 2016, 6, 35617. [Google Scholar] [CrossRef] [Green Version]
- Ohge, H.; Furne, J.K.; Springfield, J.; Sueda, T.; Madoff, R.D.; Levitt, M.D. The effect of antibiotics and bismuth on fecal hydrogen sulfide and sulfate-reducing bacteria in the rat. FEMS Microbiol. Lett. 2003, 228, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Folsom, J.P.; Baker, B.; Stewart, P.S. In vitro efficacy of bismuth thiols against biofilms formed by bacteria isolated from human chronic wounds. J. Appl. Microbiol. 2011, 111, 989–996. [Google Scholar] [CrossRef]
- Domenico, P.; Baldassarri, L.; Schoch, P.E.; Kaehler, K.; Sasatsu, M.; Cunha, B.A. Activities of bismuth thiols against Staphylococci and Staphylococcal Biofilms. Antimicrob. Agents Chemother. 2001, 45, 1417–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Morain, N.R.; Dore, M.P.; O’Connor, A.J.P.; Gisbert, J.P.; O’Morain, C.A. Treatment of Helicobacter pylori infection in 2018. Helicobacter 2018, 23, e12519. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Lai, T.-P.; Gao, P.; Zhang, H.; Ho, P.-L.; Woo, P.C.-Y.; Ma, G.; Kao, R.Y.-T.; Li, H.; Sun, H. Bismuth antimicrobial drugs serve as broad-spectrum metallo-β-lactamase inhibitors. Nat. Commun. 2018, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Keogan, D.; Griffith, D. Current and potential applications of bismuth-based drugs. Molecules 2014, 19, 15258–15297. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Bacteria | Strain 1 | MIC of Bi (µM) |
|---|---|---|
| Aeromonas hydrophila | 11 | 520 |
| A. hydrophila | 3 | 260 |
| A. salmonicida | VT45.1 | 78 |
| A. salmonicida | RSP74.1 | 13 |
| A. sobria | 10 | 130 |
| Lactococcus garvieae | 2 | 260 |
| L. piscium | 1 | >780 |
| Photobacterium damselae subsp. piscicida | DI21 | 39 |
| P. damselae subsp. piscicida | LC14 | 39 |
| Pseudomonas anguilliseptica | 8 | 39 |
| Vibrio alginolyticus | ACRp31.1 | >780 |
| V. alginolyticus | LVlenguado 27-10-09 | 39 |
| V. anguillarum | RV22 | 130 |
| V. anguillarum | 775 | 130 |
| V. campbellii | ATCC 25920 2 | >780 |
| V. costicola | ACC10.2 | 13 |
| V. costicola | ARG67.1 | 13 |
| V. diazotrophicus | BLM05-9.1.1 | >780 |
| V. fischeri | IEO15.2 | 78 |
| V. fischeri | ARG 314.1 | 13 |
| V. furnissii | ACR159.1 | >780 |
| V. harveyi | AQP 15.2 | 130 |
| V. hollisae | IEO31.2 | 78 |
| V. ichtyoenteri | RPM799.1 | 39 |
| V. iliopiscarius | S-SL1.2/07 | >780 |
| V. mediterranei | ACRp62.1 | 78 |
| V. metschnikovii | ATCC 7708 2 | >780 |
| V. mimicus | CECT 4218 2 | 260 |
| V. nigrapulchritudo | Nuno-3 | 39 |
| V. pelagius | RI145.1 | 39 |
| V. scophthalmi | ACR 318.1 | 78 |
| V. splendidus biotype I | AZ233.1 | 78 |
| V. tapetis | CPV7.1 | 39 |
| Yersinia ruckeri | 4 | >780 |
| Y. ruckeri | 730 | >780 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balado, M.; Rey-Varela, D.; Albela, A.M.; Lemos, M.L. Susceptibility to Bismuth(III) of Aquaculture Bacterial Pathogens: Effectiveness of Bismuth–Deferiprone Therapy against Vibrio anguillarum Infection in Fish. Microorganisms 2021, 9, 2399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112399
Balado M, Rey-Varela D, Albela AM, Lemos ML. Susceptibility to Bismuth(III) of Aquaculture Bacterial Pathogens: Effectiveness of Bismuth–Deferiprone Therapy against Vibrio anguillarum Infection in Fish. Microorganisms. 2021; 9(11):2399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112399
Chicago/Turabian StyleBalado, Miguel, Diego Rey-Varela, Ana M. Albela, and Manuel L. Lemos. 2021. "Susceptibility to Bismuth(III) of Aquaculture Bacterial Pathogens: Effectiveness of Bismuth–Deferiprone Therapy against Vibrio anguillarum Infection in Fish" Microorganisms 9, no. 11: 2399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112399