
Evidence of Borrelia theileri in Wild and Domestic Animals in the Kafue Ecosystem of Zambia

 ,
,  , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Real-Time and Conventional PCRs
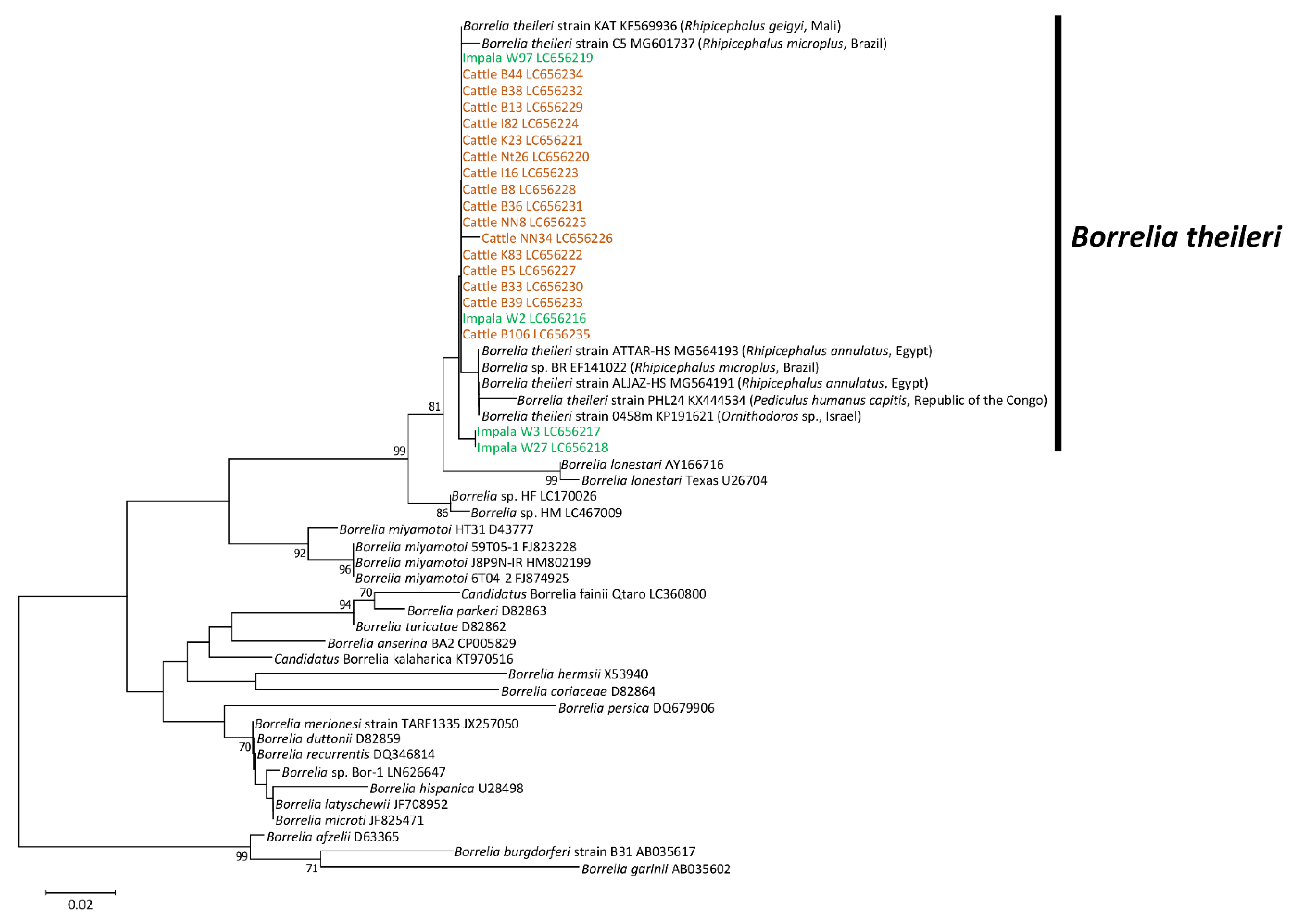
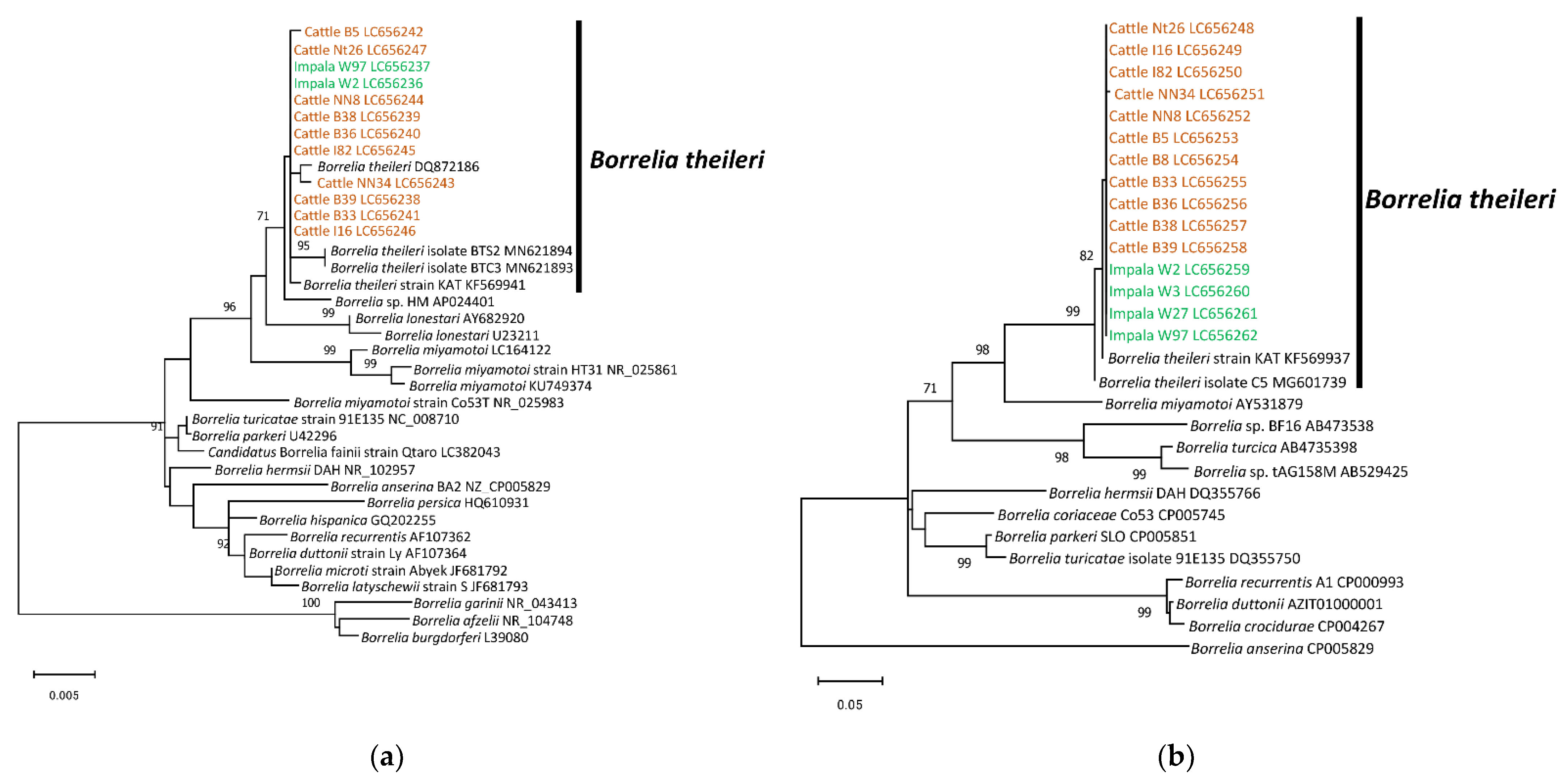
3.2. Sequence Analysis
3.3. Phylogenetic Analysis
3.4. Statistical Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takano, A.; Goka, K.; Une, Y.; Shimada, Y.; Fujita, H.; Shiino, T.; Watanabe, H.; Kawabata, H. Isolation and characterization of a novel Borrelia group of tick-borne borreliae from imported reptiles and their associated ticks. Environ. Microbiol. 2010, 12, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Franke, J.; Hildebrandt, A.; Dorn, W. Exploring gaps in our knowledge on Lyme borreliosis spirochaetes—Updates on complex heterogeneity, ecology, and pathogenicity. Ticks Tick Borne Dis. 2013, 4, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Cinco, M.; Trevisini, S.; Di Meo, N.; Ruscio, M.; Forgione, P.; Bonin, S. Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia. Biology 2021, 10, 1117. [Google Scholar] [CrossRef]
- Trape, J.-F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.H.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mané, Y.; et al. The Epidemiology and Geographic Distribution of Relapsing Fever Borreliosis in West and North Africa, with a Review of the Ornithodoros erraticus Complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef]
- Smith, R.D.; Brener, J.; Osorno, M.; Ristic, M. Pathobiology of Borrelia theileri in the tropical cattle tick, Boophilus microplus. J. Invertebr. Pathol. 1978, 32, 182–190. [Google Scholar] [CrossRef]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and Phenotypic Analysis of Borrelia miyamotoi sp. nov., Isolated from the Ixodid Tick Ixodes persulcatus, the Vector for Lyme Disease in Japan. Int. J. Syst. Evol. Microbiol. 1995, 45, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Barbour, A.G.; Maupin, G.O.; Teltow, G.J.; Carter, C.J.; Piesman, J. Identification of an uncultivable Borrelia species in the hard tick Amblyomma americanum: Possible agent of a Lyme disease-like illness. J. Infect. Dis. 1996, 173, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Scoles, G.A.; Papero, M.; Beati, L.; Fish, D. A relapsing fever group spirochete transmitted by Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001, 1, 21–34. [Google Scholar] [CrossRef]
- Brouqui, P. Arthropod-Borne Diseases Associated with Political and Social Disorder. Annu. Rev. Entomol. 2011, 56, 357–374. [Google Scholar] [CrossRef]
- McNeil, E.; Hinshaw, W.R.; Kissling, R.E. A study of Borrelia anserina infection (spirocheteosis) in Turkeys. J. Bacteriol. 1949, 57, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Fingerle, V.; Cutler, S.; Gofton, A.; Stevenson, B.; Estrada-Peña, A. Controversies in bacterial taxonomy: The example of the genus Borrelia. Ticks Tick Borne Dis. 2020, 11, 101335. [Google Scholar] [CrossRef] [PubMed]
- James, A.M.; Liveris, D.; Wormser, G.P.; Schwartz, I.; Montecalvo, M.A.; Johnson, B.J. Borrelia lonestari Infection after a Bite by an Amblyomma Americanum Tick. J. Infect. Dis. 2001, 183, 1810–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stromdahl, E.Y.; Williamson, P.C.; Kollars, T.M., Jr.; Evans, S.R.; Barry, R.K.; Vince, M.A.; Dobbs, N.A. Evidence of Borrelia lonestari DNA in Amblyomma americanum (Acari: Ixodidae) removed from humans. J. Clin. Microbiol. 2003, 41, 5557–5562. [Google Scholar] [CrossRef] [Green Version]
- Feder, H.M., Jr.; Hoss, D.M.; Zemel, L.; Telford, S.R., 3rd; Dias, F.; Wormser, G.P. Southern Tick-Associated Rash Illness (STARI) in the North: STARI following a tick bite in Long Island, New York. Clin. Infect. Dis. 2011, 53, e142–e146. [Google Scholar] [CrossRef] [Green Version]
- Burkot, T.R.; Mullen, G.R.; Anderson, R.; Schneider, B.S.; Happ, C.M.; Zeidner, N.S. Borrelia lonestari DNA in Adult Amblyomma americanum Ticks, Alabama. Emerg. Infect. Dis. 2001, 7, 471–473. [Google Scholar] [CrossRef]
- Moore, V.A.t.; Varela, A.S.; Yabsley, M.J.; Davidson, W.R.; Little, S.E. Detection of Borrelia lonestari, putative agent of southern tick-associated rash illness, in white-tailed deer (Odocoileus virginianus) from the southeastern United States. J. Clin. Microbiol. 2003, 41, 424–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyer, P.L.; Varela, A.S.; Luttrell, M.P.; Moore, V.A.t.; Stallknecht, D.E.; Little, S.E. White-tailed deer (Odocoileus virginianus) develop spirochetemia following experimental infection with Borrelia lonestari. Vet. Microbiol. 2006, 115, 229–236. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans Infected with Relapsing Fever Spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef]
- Krause, P.J.; Fish, D.; Narasimhan, S.; Barbour, A.G. Borrelia miyamotoi infection in nature and in humans. Clin. Microbiol. Infect. 2015, 21, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Hickling, G.J.; Tsao, J.I. High Prevalence of Borrelia miyamotoi among Adult Blacklegged Ticks from White-Tailed Deer. Emerg. Infect. Dis. 2016, 22, 316–318. [Google Scholar] [CrossRef]
- Laveran, A. Sur la spirillose des bovidés. CR Acad. Sci. Paris 1903, 136, 939–941. [Google Scholar]
- Theiler, A. Spirillosis of cattle. J. Comp. Path 1904, 17, 47–55. [Google Scholar] [CrossRef]
- Callow, L.L. Observations on tick-transmitted spirochaetes of cattle in Australia and South Africa. Br. Vet. J. 1967, 123, 492–497. [Google Scholar] [CrossRef]
- Uilenberg, G.; Hinaidy, H.K.; Perie, N.M.; Feenstra, T. Borrelia infections of ruminants in Europe. Vet. Q. 1988, 10, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.; Abdissa, A.; Adamu, H.; Tolosa, T.; Gashaw, A. Borrelia in Ethiopian ticks. Ticks Tick Borne Dis. 2012, 3, 14–17. [Google Scholar] [CrossRef]
- Amanzougaghene, N.; Akiana, J.; Mongo Ndombe, G.; Davoust, B.; Nsana, N.S.; Parra, H.-J.; Fenollar, F.; Raoult, D.; Mediannikov, O. Head Lice of Pygmies Reveal the Presence of Relapsing Fever Borreliae in the Republic of Congo. PLoS Negl. Trop. Dis. 2016, 10, e0005142. [Google Scholar] [CrossRef]
- Kumagai, Y.; Sato, K.; Taylor, K.R.; Zamoto-Niikura, A.; Imaoka, K.; Morikawa, S.; Ohnishi, M.; Kawabata, H. A relapsing fever group Borrelia sp. is widely distributed among wild deer in Japan. Ticks Tick Borne Dis. 2018, 9, 465–470. [Google Scholar] [CrossRef]
- Kirchgessner, M.S.; Freer, H.; Whipps, C.M.; Wagner, B. Detection of Borrelia burgdorferi outer surface protein antibodies in wild white-tailed deer (Odocoileus virginianus) in New York and Pennsylvania, USA. Vet. Immunol. Immunopathol. 2013, 153, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Stöbel, K.; Schönberg, A.; Staak, C. A new non-species dependent ELISA for detection of antibodies to Borrelia burgdorferi s. l. in zoo animals. Int. J. Med. Microbiol. 2002, 291, 88–99. [Google Scholar] [CrossRef]
- Stoebel, K.; Schoenberg, A.; Streich, W.J. The seroepidemiology of Lyme borreliosis in zoo animals in Germany. Epidemiol. Infect. 2003, 131, 975–983. [Google Scholar] [CrossRef]
- Širmarová, J.; Tichá, L.; Golovchenko, M.; Salát, J.; Grubhoffer, L.; Rudenko, N.; Nowotny, N.; Růžek, D. Seroprevalence of Borrelia burgdorferi sensu lato and tick-borne encephalitis virus in zoo animal species in the Czech Republic. Ticks Tick Borne Dis. 2014, 5, 523–527. [Google Scholar] [CrossRef]
- Diatta, G.; Duplantier, J.M.; Granjon, L.; Ba, K.; Chauvancy, G.; Ndiaye, M.; Trape, J.F. Borrelia infection in small mammals in West Africa and its relationship with tick occurrence inside burrows. Acta Trop. 2015, 152, 131–140. [Google Scholar] [CrossRef]
- Qiu, Y.; Nakao, R.; Hang’ombe, B.M.; Sato, K.; Kajihara, M.; Kanchela, S.; Changula, K.; Eto, Y.; Ndebe, J.; Sasaki, M.; et al. Human Borreliosis Caused by a New World Relapsing Fever Borrelia-like Organism in the Old World. Clin. Infect. Dis. 2019, 69, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Squarre, D.; Nakamura, Y.; Hayashida, K.; Kawai, N.; Chambaro, H.; Namangala, B.; Sugimoto, C.; Yamagishi, J. Investigation of the piroplasm diversity circulating in wildlife and cattle of the greater Kafue ecosystem, Zambia. Parasit. Vectors 2020, 13, 599. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Hayashida, K.; Delesalle, V.; Qiu, Y.; Omori, R.; Simuunza, M.; Sugimoto, C.; Namangala, B.; Yamagishi, J. Genetic Diversity of African Trypanosomes in Tsetse Flies and Cattle from the Kafue Ecosystem. Front. Vet. Sci. 2021, 8, 599815. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Ryelandt, J.; Mangold, A.J.; Mediannikov, O.; Guglielmone, A.A.; Raoult, D. Relapsing fever Borrelia in Ornithodoros ticks from Bolivia. Ann. Trop. Med. Parasitol. 2011, 105, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, A.; Fujita, H.; Kadosaka, T.; Konnai, S.; Tajima, T.; Watanabe, H.; Ohnishi, M.; Kawabata, H. Characterization of reptile-associated Borrelia sp. in the vector tick, Amblyomma geoemydae, and its association with Lyme disease and relapsing fever Borrelia spp. Environ. Microbiol. Rep. 2011, 3, 632–637. [Google Scholar] [CrossRef] [PubMed]
- McCoy, B.N.; Maiga, O.; Schwan, T.G. Detection of Borrelia theileri in Rhipicephalus geigyi from Mali. Ticks Tick Borne Dis. 2014, 5, 401–403. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Body Lice as Tools for Diagnosis and Surveillance of Reemerging Diseases. J. Clin. Microbiol. 1999, 37, 596–599. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Sharma, S.P.; Amanfu, W.; Losho, T.C. Bovine borreliosis in Botswana. Onderstepoort. J. Vet. Res. 2000, 67, 221–223. [Google Scholar]
- Abanda, B.; Paguem, A.; Abdoulmoumini, M.; Kingsley, M.T.; Renz, A.; Eisenbarth, A. Molecular identification and prevalence of tick-borne pathogens in zebu and taurine cattle in North Cameroon. Parasit. Vectors 2019, 12, 448. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Takano, A.; Taylor, K.; Sashika, M.; Shimozuru, M.; Konnai, S.; Kawabata, H.; Tsubota, T. A relapsing fever group Borrelia sp. similar to Borrelia lonestari found among wild sika deer (Cervus nippon yesoensis) and Haemaphysalis spp. ticks in Hokkaido, Japan. Ticks Tick Borne Dis. 2014, 5, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Nambota, A.; Samui, K.; Sugimoto, C.; Kakuta, T.; Onuma, M. Theileriosis in Zambia: Etiology, epidemiology and control measures. Jpn. J. Vet. Res. 1994, 42, 1–18. [Google Scholar]
- Simuunza, M.; Weir, W.; Courcier, E.; Tait, A.; Shiels, B. Epidemiological analysis of tick-borne diseases in Zambia. Vet. Parasitol. 2011, 175, 331–342. [Google Scholar] [CrossRef]
- Laohasinnarong, D.; Goto, Y.; Asada, M.; Nakao, R.; Hayashida, K.; Kajino, K.; Kawazu, S.-I.; Sugimoto, C.; Inoue, N.; Namangala, B. Studies of trypanosomiasis in the Luangwa valley, north-eastern Zambia. Parasit. Vectors 2015, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De Leeuw, P.N.; McDermott, J.J.; Lebbie, S.H.B. Monitoring of livestock health and production in sub-Saharan Africa. Prev. Vet. Med. 1995, 25, 195–212. [Google Scholar] [CrossRef]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Espinaze, M.P.A.; Hellard, E.; Horak, I.G.; Cumming, G.S. Domestic mammals facilitate tick-borne pathogen transmission networks in South African wildlife. Biol. Conserv. 2018, 221, 228–236. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Target Gene | Primer Name | Sequence (5′–3′) | Annealing Temperature | Expected Size | Reference |
|---|---|---|---|---|---|
| Flagellin (P41) | BflaPAD 1 | GATCARGCWCAAYATAACCAWATGCA | 50 °C | 453 bp | [37] |
| BflaPDU 1 | AGATTCAAGTCTGTTTTGGAAAGC | ||||
| BflaPBU 2 | GCTGAAGAGCTTGGAATGCAACC | 50 °C | 347 bp | [37] | |
| BflaPCR 2 | TGATCAGTTATCATTCTAATAGCA | ||||
| hypoxanthine-guanine phosphoribosyltransferase | hptdegF | GCAGAYATTACAAGAGARATGG | 55 °C | 433 bp | [38] |
| hptdegR | CYTCRTCACCCCATTGAGTTCC | ||||
| 16S ribosomal DNA | BF1 | GCTGGCAGTGCGTCTTAAGC | 55 °C | 1371 bp | [39] |
| BR1 | GCTTCGGGTATCCTCAACTC |
| Species | Real-Time PCR | flaB | 16S rDNA | hpt |
|---|---|---|---|---|
| Impala | 5/97 (5.1%) | 4/5 | 2/5 | 4/5 |
| Hartebeest | 0/37 (0%) | NA | NA | NA |
| Lion | 0/4 (0%) | NA | NA | NA |
| Wild dog | 0/2 (0%) | NA | NA | NA |
| Cattle | 20/488 (4.1%) | 16/20 | 10/20 | 11/20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Squarre, D.; Nakamura, Y.; Lau, A.C.C.; Moonga, L.C.; Kawai, N.; Ohnuma, A.; Hayashida, K.; Nakao, R.; Yamagishi, J.; et al. Evidence of Borrelia theileri in Wild and Domestic Animals in the Kafue Ecosystem of Zambia. Microorganisms 2021, 9, 2405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112405
Qiu Y, Squarre D, Nakamura Y, Lau ACC, Moonga LC, Kawai N, Ohnuma A, Hayashida K, Nakao R, Yamagishi J, et al. Evidence of Borrelia theileri in Wild and Domestic Animals in the Kafue Ecosystem of Zambia. Microorganisms. 2021; 9(11):2405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112405
Chicago/Turabian StyleQiu, Yongjin, David Squarre, Yukiko Nakamura, Alice C. C. Lau, Lavel Chinyama Moonga, Naoko Kawai, Aiko Ohnuma, Kyoko Hayashida, Ryo Nakao, Junya Yamagishi, and et al. 2021. "Evidence of Borrelia theileri in Wild and Domestic Animals in the Kafue Ecosystem of Zambia" Microorganisms 9, no. 11: 2405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112405