
New Insights into the Host–Pathogen Interaction of Mycoplasma gallisepticum and Avian Metapneumovirus in Tracheal Organ Cultures of Chicken

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tracheal Organ Culture
2.2. Virus and Bacterium
2.3. Experimental Design
2.3.1. RNA and DNA Isolation
2.3.2. Quantification of Pathogens by (RT)qPCR
2.3.3. Immunohistochemical (IHC) Staining
2.3.4. Transmission Electron Microscopy
2.3.5. Ciliostasis Assay
2.3.6. Histopathological Examination
2.3.7. Determination of Cytokine mRNA Expression by RTqPCR
2.4. Statistical Analysis
3. Results
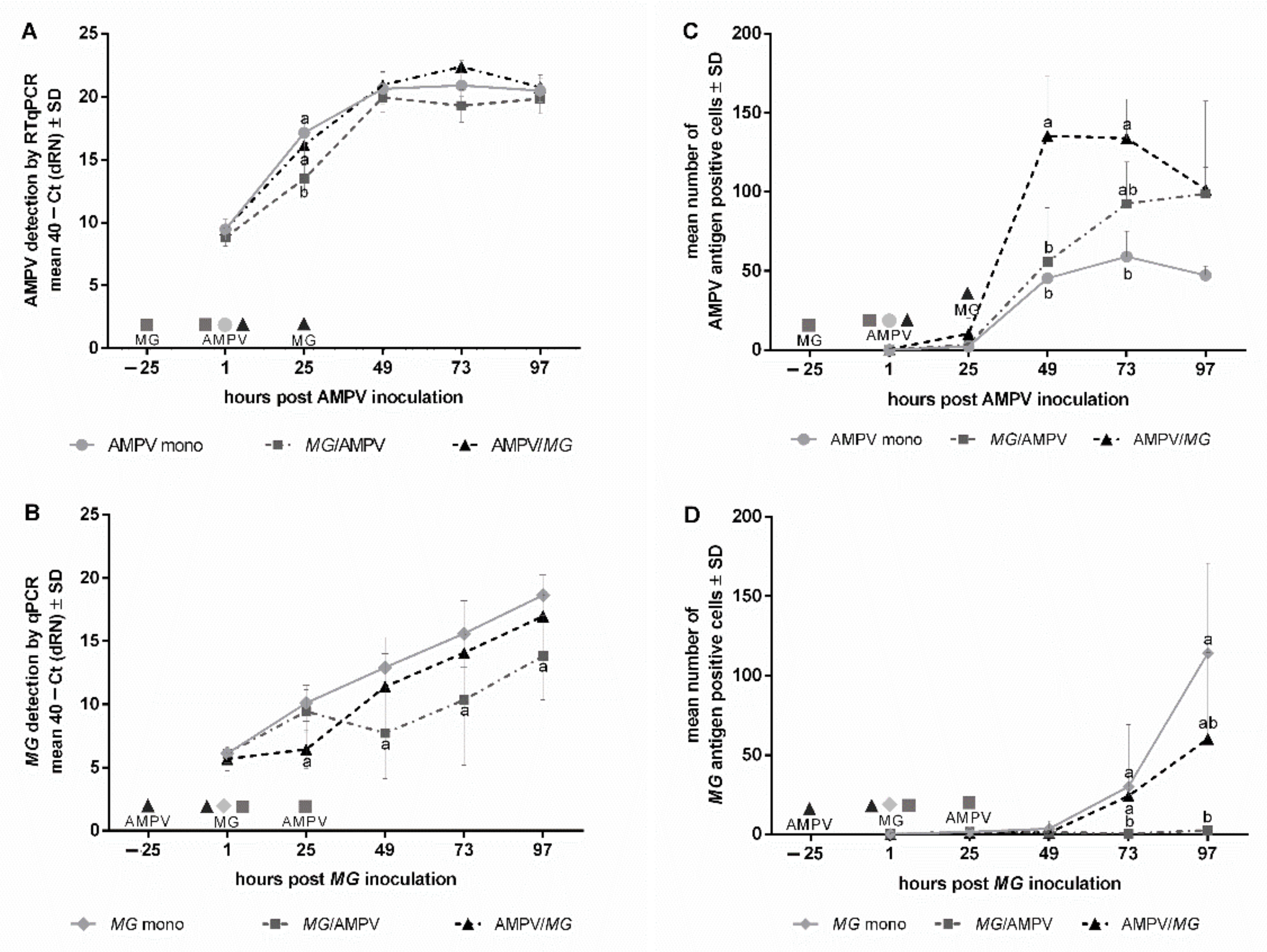
3.1. The Effect of the Inoculation-Scheme on M. gallispeticum and AMPV Replication
Quantification and Detection of Viral Colonization and Replication
3.2. Quantification and Detection of Bacterial Colonization and Replication
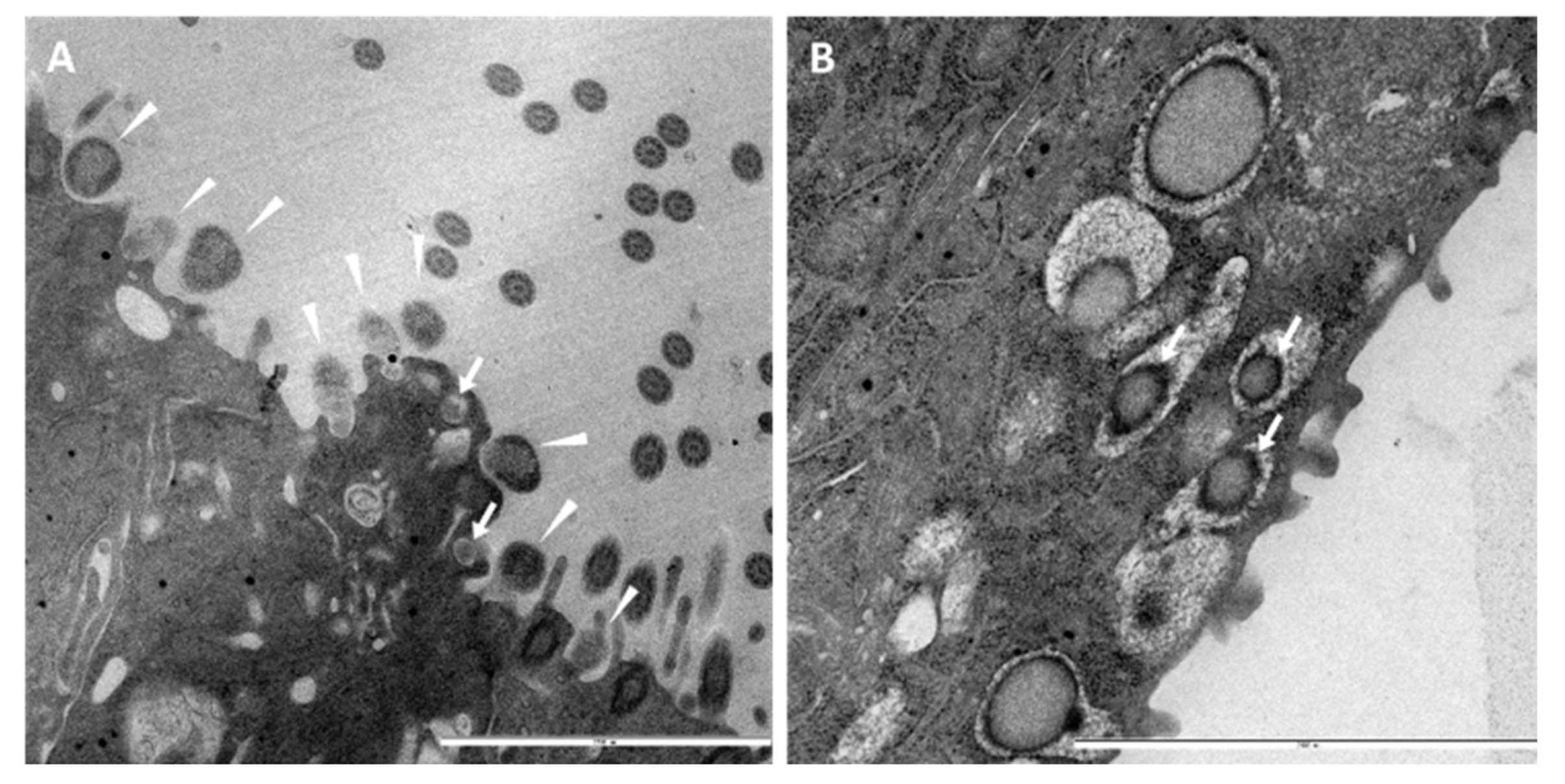
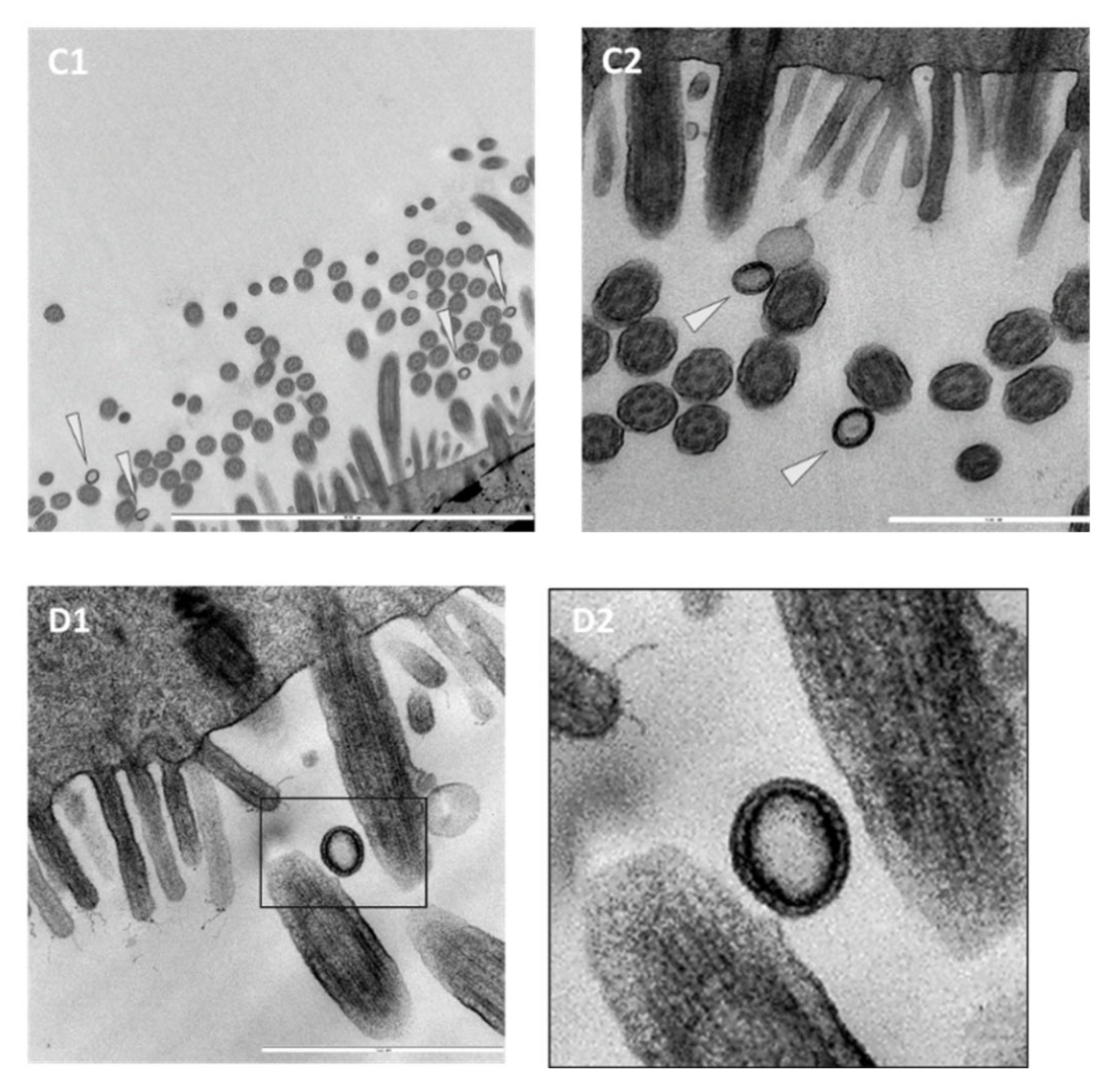
3.3. Ultrastructural Insights into Host–Pathogen Interactions after AMPV and M. gallispeticum Mono- and Dual-Infection
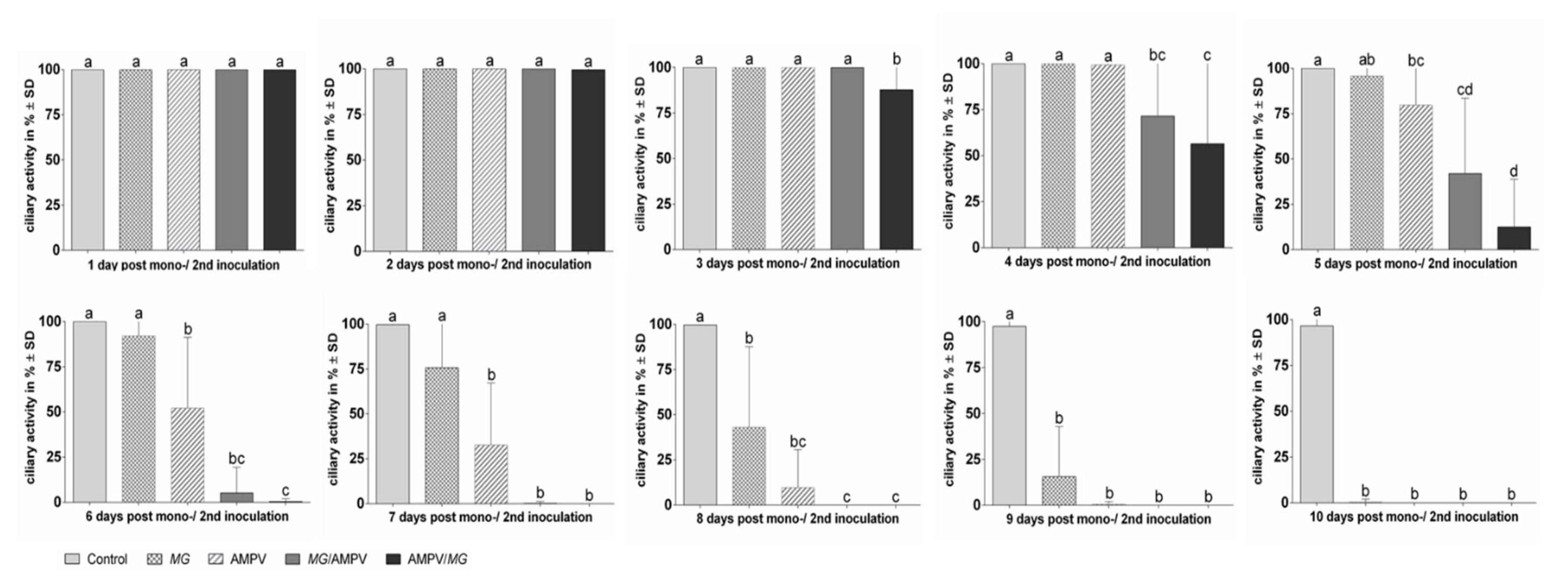
3.4. The Effect of AMPV and M. gallispeticum Mono- and Dual-Infection on the Lesion Development
Ciliostasis Assay
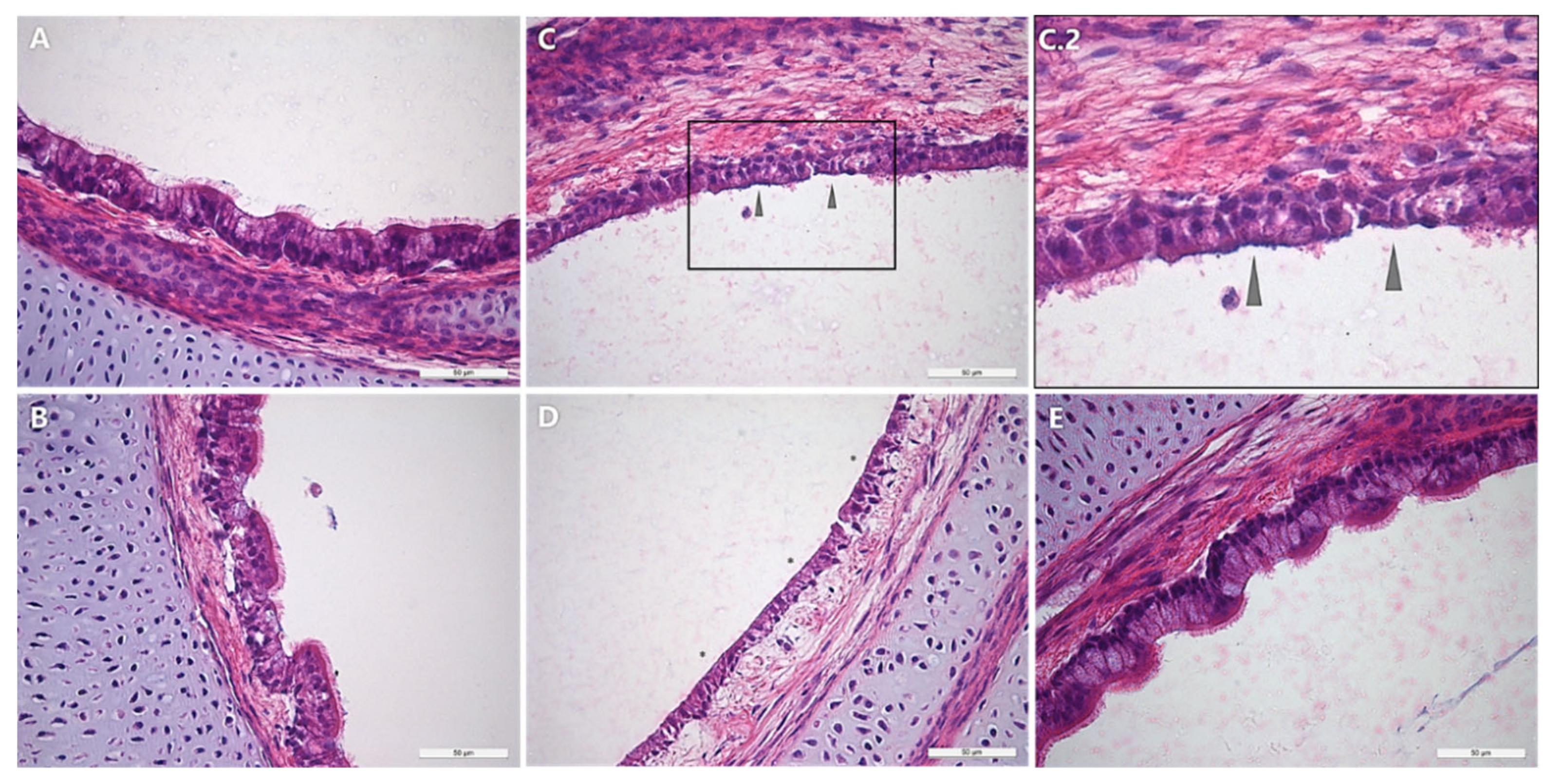
3.5. Histopathological Examination
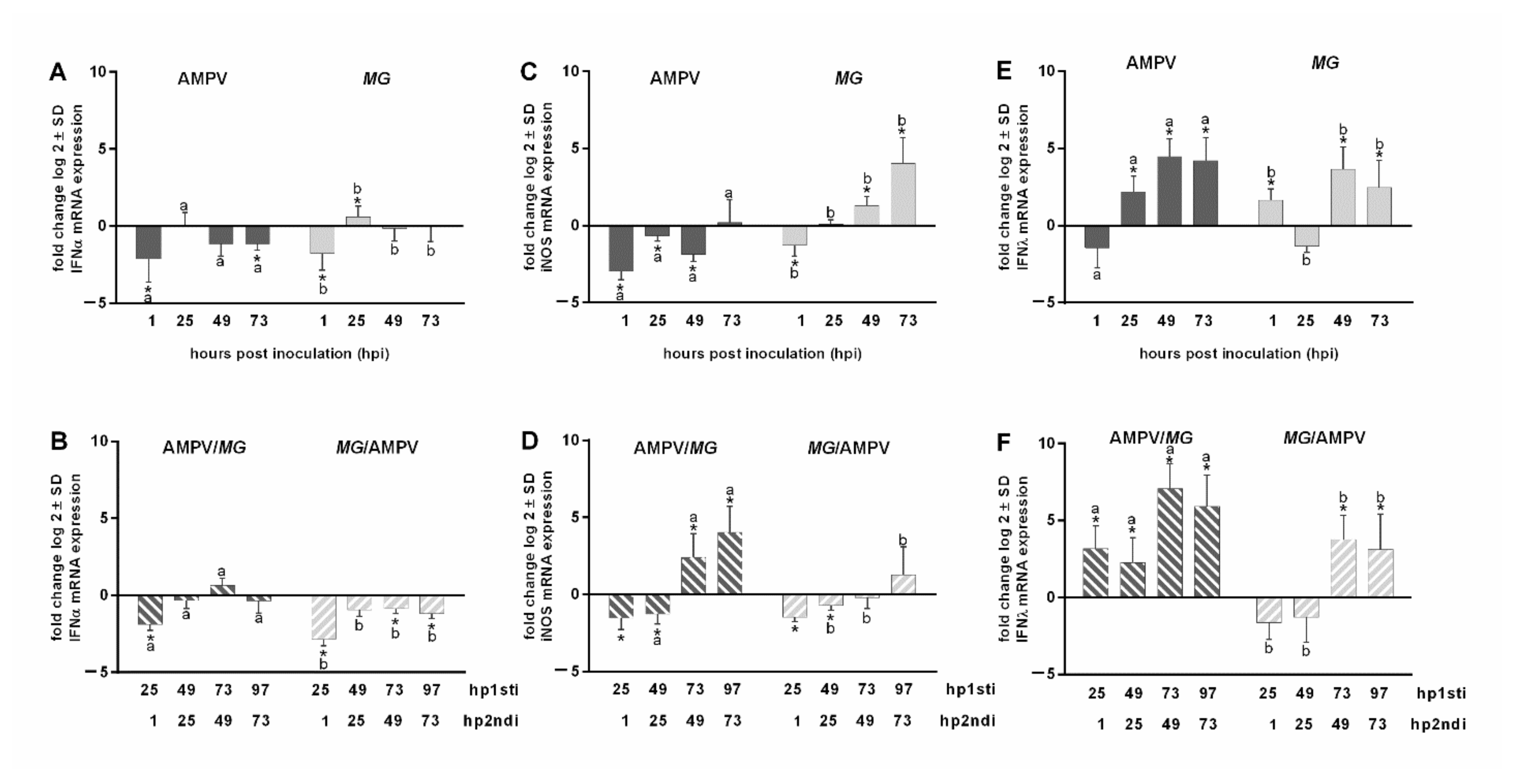
3.6. The Effect of M. gallispeticum and AMPV Mono- and Co-Infection on Innate Immune Responses
Quantification of IFNα, iNOS, and IFNλ mRNA Expression
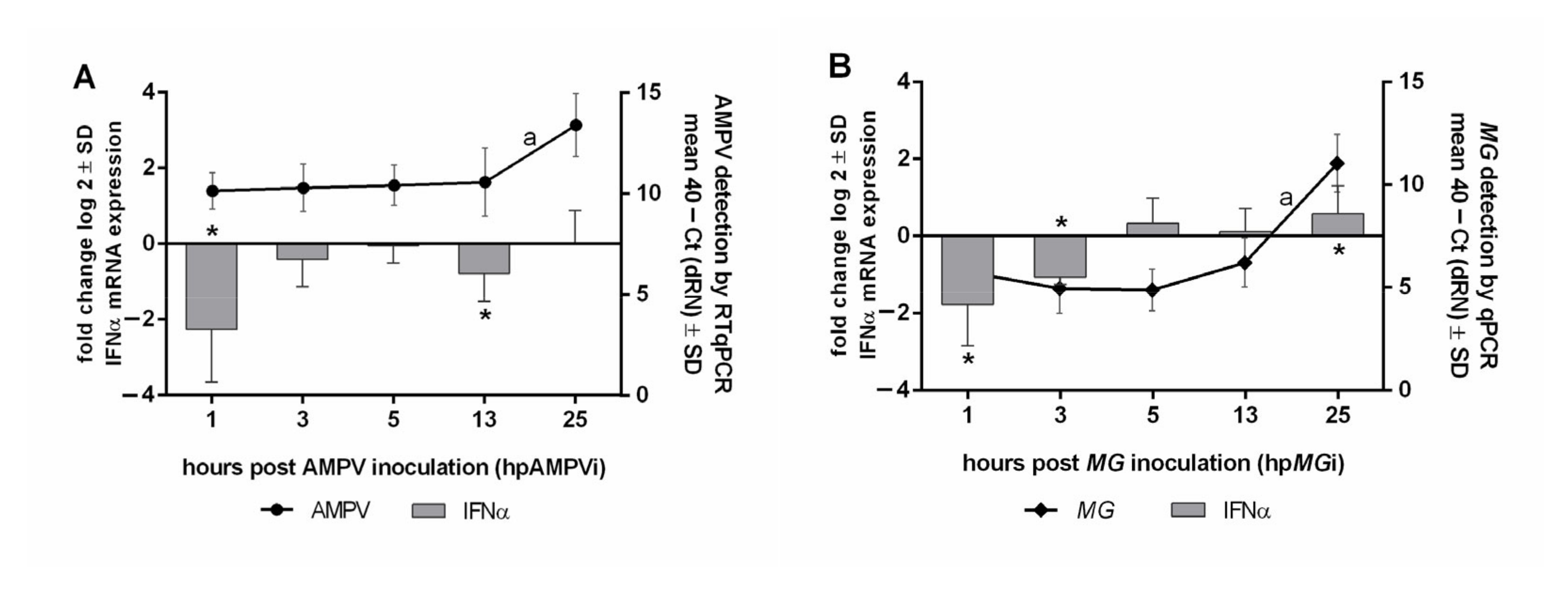
3.7. Correlation of Pathogen Replication and IFNα mRNA Expression 1 to 25 hpi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohammed, H.O.; Carpenter, T.E.; Yamamoto, R. Economic Impact of Mycoplasma gallisepticum and M. synoviae in Commercial Layer Flocks. Avian Dis. 1987, 31, 477. [Google Scholar] [CrossRef]
- Felice, V.; Lupini, C.; Mescolini, G.; Silveira, F.; Guerrini, A.; Catelli, E.; Di Francesco, A. Molecular detection and characterization of Mycoplasma gallisepticum and Mycoplasma synoviae strains in backyard poultry in Italy. Poult. Sci. 2020, 99, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Sid, H.; Benachour, K.; Rautenschlein, S. Co-infection with Multiple Respiratory Pathogens Contributes to Increased Mortality Rates in Algerian Poultry Flocks. Avian Dis. 2015, 59, 440–446. [Google Scholar] [CrossRef]
- Sid, H.; Hartmann, S.; Petersen, H.; Ryll, M.; Rautenschlein, S. Mycoplasma gallisepticum modifies the pathogenesis of influenza A virus in the avian tracheal epithelium. Int. J. Med Microbiol. 2016, 306, 174–186. [Google Scholar] [CrossRef]
- Hutton, S.; Bettridge, J.; Christley, R.; Habte, T.; Ganapathy, K. Detection of infectious bronchitis virus 793B, avian metapneumovirus, Mycoplasma gallisepticum and Mycoplasma synoviae in poultry in Ethiopia. Trop. Anim. Heal. Prod. 2016, 49, 317–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samy, A.; Naguib, M.M. Avian Respiratory Coinfection and Impact on Avian Influenza Pathogenicity in Domestic Poultry: Field and Experimental Findings. Veter. Sci. 2018, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Naylor, C.J.; Al-Ankari, A.R.; Al-Afaleq, A.I.; Bradbury, J.M.; Jones, R.C. Exacerbation of Mycoplasma gallisepticum infection in turkeys by rhinotracheitis virus. Avian Pathol. 1992, 21, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stipkovits, L.; Egyed, L.; Palfi, V.; Beres, A.; Pitlik, E.; Somogyi, M.; Szathmary, S.; Denes, B. Effect of low-pathogenicity influenza virus H3N8 infection on Mycoplasma gallisepticum infection of chickens. Avian Pathol. J. W.V.P.A. 2012, 41, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Stipkovits, L.; Glavits, R.; Palfi, V.; Beres, A.; Egyed, L.; Dénes, B.; Somogyi, M.; Szathmary, S. Pathologic Lesions Caused by Coinfection of Mycoplasma gallisepticum and H3N8 Low Pathogenic Avian Influenza Virus in Chickens. Veter. Pathol. 2011, 49, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Kishida, N.; Sakoda, Y.; Eto, M.; Sunaga, Y.; Kida, H. Co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2 influenza A virus infection in chickens. Arch. Virol. 2004, 149, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, A.; Zhang, F.; Ling, Y.; Ou, C.; Hou, N.; He, C. Co-infection of broilers with Ornithobacterium rhinotracheale and H9N2 avian influenza virus. BMC Veter. Res. 2012, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghighat-Jahromi, M.; Asasi, K.; Nili, H.; Dadras, H.; Shooshtari, A.H. Coinfection of avian influenza virus (H9N2 subtype) with infectious bronchitis live vaccine. Arch. Virol. 2008, 153, 651–655. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Lupini, C.; Alejo, C.T.; Listorti, V.; Mescolini, G.; Brandão, P.E.; Martini, M.; Catelli, E.; Cecchinato, M. Avian Metapneumovirus circulation in Italian broiler farms. Poult. Sci. 2018, 97, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Ball, C.; Forrester, A.; Ganapathy, K. Co-circulation of genetically diverse population of vaccine related and unrelated respiratory mycoplasmas and viruses in UK poultry flocks with health or production problems. Veter. Microbiol. 2018, 225, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Roussan, D.; Haddad, R.; Khawaldeh, G. Molecular Survey of Avian Respiratory Pathogens in Commercial Broiler Chicken Flocks with Respiratory Diseases in Jordan. Poult. Sci. 2008, 87, 444–448. [Google Scholar] [CrossRef]
- Richard, C.J.; Rautenschlein, S. Avian Metapneumovirus, in Diseases of Poultry; Swayne, D.E., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 112–119. [Google Scholar]
- Falsey, A.R.; Becker, K.L.; Swinburne, A.J.; Nylen, E.S.; Formica, M.A.; Hennessey, P.A.; Criddle, M.M.; Peterson, D.R.; Baran, A.; Walsh, E.E. Bacterial Complications of Respiratory Tract Viral Illness: A Comprehensive Evaluation. J. Infect. Dis. 2013, 208, 432–441. [Google Scholar] [CrossRef] [Green Version]
- Almand, E.A.; Moore, M.D.; Jaykus, L.-A. Virus-Bacteria Interactions: An Emerging Topic in Human Infection. Viruses 2017, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.A.T.M.; Biesbroek, G.; Trzcinski, K.; Sanders, E.A.M.; Bogaert, D. Viral and Bacterial Interactions in the Upper Respiratory Tract. PLOS Pathog. 2013, 9, e1003057. [Google Scholar] [CrossRef] [Green Version]
- Tajima, M.; Nunoya, T.; Yagihashi, T. An ultrastructural study on the interaction of Mycoplasma gallisepticum with the chicken tracheal epithelium. Am. J. Veter. Res. 1979, 40, 1009–1014. [Google Scholar]
- Hartmann, S.; Sid, H.; Rautenschlein, S. Avian metapneumovirus infection of chicken and turkey tracheal organ cultures: Comparison of virus–host interactions. Avian Pathol. 2015, 44, 480–489. [Google Scholar] [CrossRef]
- Winner, F.; Rosengarten, R.; Citti, C. In Vitro Cell Invasion of Mycoplasma gallisepticum. Infect. Immun. 2000, 68, 4238–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyushkina, D.; Pobeguts, O.; Butenko, I.; Vanyushkina, A.; Anikanov, N.; Bukato, O.; Evsyutina, D.; Bogomazova, A.; Lagarkova, M.; Semashko, T.; et al. Phase Transition of the Bacterium upon Invasion of a Host Cell as a Mechanism of Adaptation: A Mycoplasma gallisepticum Model. Sci. Rep. 2016, 6, 35959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rima, B.; Collins, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.A.; Lee, B.; Maisner, A.; Rota, P.; Wang, L.; et al. ICTV Virus Taxonomy Profile: Pneumoviridae. J. Gen. Virol. 2017, 98, 2912–2913. [Google Scholar] [CrossRef]
- Rosengarten, R.; Citti, C.; Glew, M.; Lischewski, A.; Droeße, M.; Much, P.; Winner, F.; Brank, M.; Spergser, J. Host-pathogen interactions in mycoplasma pathogenesis: Virulence and survival strategies of minimalist prokaryotes. Int. J. Med Microbiol. 2000, 290, 15–25. [Google Scholar] [CrossRef]
- Vogl, G.; Plaickner, A.; Szathmary, S.; Stipkovits, L.; Rosengarten, R.; Szostak, M.P. Mycoplasma gallisepticum Invades Chicken Erythrocytes during Infection. Infect. Immun. 2008, 76, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Hardy, R.D.; Atkinson, T.P.; Cassell, G.H. Immune Responses to Mycoplasma. Mucosal Immunology 2005, 1451–1464. [Google Scholar]
- Rivera-Benitez, J.F.; Martínez-Bautista, R.; Ríos-Cambre, F.; Ramírez-Mendoza, H. Molecular detection and isolation of avian metapneumovirus in Mexico. Avian Pathol. 2014, 43, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.K.A.; Kinloch, S.; Ellis, M.M. In vitro and in vivo studies in chickens and turkeys on strains of turkey rhinotracheitis virus isolated from the two species. Avian Pathol. 1993, 22, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Borchsenius, S.N.; Daks, A.; Fedorova, O.; Chernova, O.; Barlev, N.A. Effects of mycoplasma infection on the host organism response via p53/NF-κB signaling. J. Cell. Physiol. 2018, 234, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Goraya, M.U.; Ali, L.; Younis, I. Innate Immune Responses Against Avian Respiratory Viruses. Hosts Viruses 2017, 4, 4. [Google Scholar] [CrossRef]
- Reuter, A.; Soubies, S.M.; Härtle, S.; Schusser, B.; Kaspers, B.; Staeheli, P.; Rubbenstroth, D.; Garcia-Sastre, A. Antiviral Activity of Lambda Interferon in Chickens. J. Virol. 2013, 88, 2835–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolli, D.; Bao, X.; Casola, A. Human Metapneumovirus Antagonism of Innate Immune Responses. Viruses 2012, 4, 3551–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oven, I.; Rus, K.R.; Dušanić, D.; Benčina, D.; Keeler, C.L.; Narat, M. Diacylated lipopeptide from Mycoplasma synoviae mediates TLR15 induced innate immune responses. Veter. Res. 2013, 44, 99. [Google Scholar] [CrossRef]
- Mohammed, J.; Frasca, S.; Cecchini, K.; Rood, D.; Nyaoke, A.C.; Geary, S.J.; Silbart, L.K. Chemokine and cytokine gene expression profiles in chickens inoculated with Mycoplasma gallisepticum strains Rlow or GT5. Vaccine 2007, 25, 8611–8621. [Google Scholar] [CrossRef]
- Nakayama, T.; Sonoda, S.; Urano, T.; Osano, M.; Maehara, N.; Sasaki, K.; Hayatsu, E.; Makino, S. Interferon production during the course of Mycoplasma pneumoniae infection. Pediatr. Infect. Dis. J. 1992, 11, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ding, L.; Bao, J.; Liu, Y.; Zhang, Q.; Wang, J.; Li, R.; Ishfaq, M.; Li, J. Co-infection of Mycoplasma gallisepticum and Escherichia coli Triggers Inflammatory Injury Involving the IL-17 Signaling Pathway. Front. Microbiol. 2019, 10, 2615. [Google Scholar] [CrossRef]
- Nicolas de Lamballerie, C.; Pizzorno, A.; Dubois, J.; Julien, T. ’ Padey, B.’ Bouveret, M.; Traversier, A.; Legras-Lachuer, C.; Lina, B.; Terrier, O.; et al. Characterization of cellular transcriptomic signatures induced by different respiratory viruses in human reconstituted airway epithelia. Sci. Rep. 2019, 9, 11493. [Google Scholar] [CrossRef] [Green Version]
- Liman, M.; Rautenschlein, S. Induction of local and systemic immune reactions following infection of turkeys with avian Metapneumovirus (aMPV) subtypes A and B. Veter. Immunol. Immunopathol. 2007, 115, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Aung, Y.H.; Liman, M.; Neumann, U.; Rautenschlein, S. Reproducibility of swollen sinuses in broilers by experimental infection with avian metapneumovirus subtypes A and B of turkey origin and their comparative pathogenesis. Avian Pathol. 2008, 37, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Froehlich, E.; Salar-Behzadi, S. Toxicological Assessment of Inhaled Nanoparticles: Role of in Vivo, ex Vivo, in Vitro, and in Silico Studies. Int. J. Mol. Sci. 2014, 15, 4795–4822. [Google Scholar] [CrossRef] [PubMed]
- Dobrescu, I.; Levast, B.; Lai, K.; Delgado-Ortega, M.; Walker, S.; Banman, S.; Townsend, H.; Simon, G.; Zhou, Y.; Gerdts, V.; et al. In vitro and ex vivo analyses of co-infections with swine influenza and porcine reproductive and respiratory syndrome viruses. Veter. Microbiol. 2014, 169, 18–32. [Google Scholar] [CrossRef]
- Cherry, J.; Taylor-Robinson, D.J.A.M. Large-quantity production of chicken embryo tracheal organ cultures and use in virus and mycoplasma studies. Appl. Microbiol. 1970, 19, 658–662. [Google Scholar] [CrossRef] [PubMed]
- A Biron, C. Role of early cytokines, including alpha and beta interferons (IFN-α\β), in innate and adaptive immune responses to viral infections. Semin. Immunol. 1998, 10, 383–390. [Google Scholar] [CrossRef]
- Zhou, J.-H.; Wang, Y.-N.; Chang, Q.-Y.; Ma, P.; Hu, Y.; Cao, X. Type III Interferons in Viral Infection and Antiviral Immunity. Cell. Physiol. Biochem. 2018, 51, 173–185. [Google Scholar] [CrossRef]
- Chakravortty, D.; Hensel, M. Inducible nitric oxide synthase and control of intracellular bacterial pathogens. Microbes Infect. 2003, 5, 621–627. [Google Scholar] [CrossRef]
- Reiss, C.; Komatsu, T. Does Nitric Oxide Play a Critical Role in Viral Infections? J. Virol. 1998, 72, 4547–4551. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.K.A.; Darbyshire, J.H.; Peters, R.W. The use of chicken tracheal organ cultures for the isolation and assay of avian infectious bronchitis virus. Arch. Virol. 1976, 50, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Albers, A.C.; Fletcher, R.D. Simple method for quantitation of viable mycoplasmas. Appl. Environ. Microbiol. 1982, 43, 958–960. [Google Scholar] [CrossRef] [Green Version]
- Brealey, J.C.; Sly, P.D.; Young, P.R.; Chappell, K.J. Viral bacterial co-infection of the respiratory tract during early childhood. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [Green Version]
- De Graaf, M.; Osterhaus, A.; Fouchier, R.; Holmes, E.C. Evolutionary dynamics of human and avian metapneumoviruses. J. Gen. Virol. 2018, 89 Pt 12, 2933–2942. [Google Scholar] [CrossRef]
- Verkaik, N.; Nguyen, D.; De Vogel, C.; Moll, H.A.; Verbrugh, H.; Jaddoe, V.; Hofman, A.; Van Wamel, W.; Hoogen, B.V.D.; Buijs-Offerman, R.; et al. Streptococcus pneumoniae exposure is associated with human metapneumovirus seroconversion and increased susceptibility to in vitro HMPV infection. Clin. Microbiol. Infect. 2011, 17, 1840–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Li, S.; Zhu, C.; Zhou, R.; Leung, P. Mycoplasma pneumoniae Infections: Pathogenesis and Vaccine Development. Pathogens 2021, 10, 119. [Google Scholar] [CrossRef]
- Christodoulides, A.; Gupta, N.; Yacoubian, V.; Maithel, N.; Parker, J.; Kelesidis, T. The Role of Lipoproteins in Mycoplasma-Mediated Immunomodulation. Front. Microbiol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P. Nitric oxide and cell death. Biochimica et Biophysica Acta (BBA). Bioenergetics 1999, 1411, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Arachchige, S.N.K.; Young, N.D.; Shil, P.K.; Legione, A.R.; Condello, A.K.; Browning, G.F.; Wawegama, N.K. Differential Response of the Chicken Trachea to Chronic Infection with Virulent Mycoplasma gallisepticum Strain Ap3AS and Vaxsafe MG (Strain ts-304): A Transcriptional Profile. Infect. Immun. 2020, 88, e00053-20. [Google Scholar] [CrossRef]
- Hatchel, J.M.; Balish, M.F. Attachment organelle ultrastructure correlates with phylogeny, not gliding motility properties, in Mycoplasma pneumoniae relatives. Microbiol. 2008, 154, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.R.; Chen, L.; Driver, A.D.; Arnold, E.A.; Sheppard, E.S.; Locklin, J.; Krause, D.C. Sialylated Receptor Setting InfluencesMycoplasma pneumoniaeAttachment and Gliding Motility. Mol. Microbiol. 2018, 109, 735–744. [Google Scholar] [CrossRef]
- Miyata, M.; Hamaguchi, T. Integrated Information and Prospects for Gliding Mechanism of the Pathogenic Bacterium Mycoplasma pneumoniae. Front. Microbiol. 2016, 7, 960. [Google Scholar] [CrossRef] [Green Version]
- Glasgow, L.R.; Hill, R.L. Interaction of Mycoplasma gallisepticum with sialyl glycoproteins. Infect. Immun. 1980, 30, 353–361. [Google Scholar] [CrossRef]
- Cagno, V.; Tseligka, E.D.; Jones, S.T.; Tapparel, C. Heparan Sulfate Proteoglycans and Viral Attachment: True Receptors or Adaptation Bias? Viruses 2019, 11, 596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pasquale, V.; Quiccione, M.S.; Tafuri, S.; Avallone, L.; Pavone, L.M. Heparan Sulfate Proteoglycans in Viral Infection and Treatment: A Special Focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 6574. [Google Scholar] [CrossRef]
- Klimyte, E.M.; Smith, S.E.; Oreste, P.; Lembo, D.; Dutch, R.E. Inhibition of Human Metapneumovirus Binding to Heparan Sulfate Blocks Infection in Human Lung Cells and Airway Tissues. J. Virol. 2016, 90, 9237–9250. [Google Scholar] [CrossRef] [Green Version]
- Thammawat, S.; A Sadlon, T.; Adamson, P.; Gordon, D.L. Effect of sialidase fusion protein (DAS 181) on human metapneumovirus infection of Hep-2 cells. Antivir. Chem. Chemother. 2015, 24, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanardi, D.; Lupini, C.; Pesente, P.; Rossi, G.; Ortali, G.; Catelli, E. Longitudinal field studies of Avian Metapneumovirus and Turkey Hemorrhagic Enteritis Virus in turkeys suffering from colibacillosis associated mortality. Veter. Res. Commun. 2014, 38, 129–137. [Google Scholar] [CrossRef]
- Van de Zande, S.; Nauwynck, H.; Pensaert, M. The clinical, pathological and microbiological outcome of an Escherichia coli O2:K1 infection in avian pneumovirus infected turkeys. Veter. Microbiol. 2001, 81, 353–365. [Google Scholar] [CrossRef]
- Barjesteh, N.; O’Dowd, K.; Vahedi, S.M. Antiviral responses against chicken respiratory infections: Focus on avian influenza virus and infectious bronchitis virus. Cytokine 2020, 127, 154961. [Google Scholar] [CrossRef]
- Pott, J.; Mahlakõiv, T.; Mordstein, M.; Duerr, C.U.; Michiels, T.; Stockinger, S.; Staeheli, P.; Hornef, M.W. IFN-lambda determines the intestinal epithelial antiviral host defense. Proc. Nat. Acad. Sci. USA 2011, 108, 7944–7949. [Google Scholar] [CrossRef] [Green Version]
- Cox, R.G.; Mainou, B.A.; Johnson, M.; Hastings, A.K.; Schuster, J.E.; Dermody, T.S.; Williams, J.V. Human Metapneumovirus Is Capable of Entering Cells by Fusion with Endosomal Membranes. PLOS Pathog. 2015, 11, e1005303. [Google Scholar] [CrossRef] [Green Version]
- Easton, A.J.; Domachowske, J.B.; Rosenberg, H.F. Animal Pneumoviruses: Molecular Genetics and Pathogenesis. Clin. Microbiol. Rev. 2004, 17, 390–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | AMPV | M. gallispeticum | Experiment 1a, 1b, 1c | Experiment 2a, 2b | Experiment 3a, 3b |
|---|---|---|---|---|---|
| Control | − | − | Pathogen colonization and replication | Development of pathological lesions | Innate immune response |
| AMPV | + | − | |||
| M. gallispeticum | − | + | |||
| AMPV/M. gallispeticum | +1 | +2 | |||
| M. gallispeticum/AMPV | +2 | +1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rüger, N.; Sid, H.; Meens, J.; Szostak, M.P.; Baumgärtner, W.; Bexter, F.; Rautenschlein, S. New Insights into the Host–Pathogen Interaction of Mycoplasma gallisepticum and Avian Metapneumovirus in Tracheal Organ Cultures of Chicken. Microorganisms 2021, 9, 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112407
Rüger N, Sid H, Meens J, Szostak MP, Baumgärtner W, Bexter F, Rautenschlein S. New Insights into the Host–Pathogen Interaction of Mycoplasma gallisepticum and Avian Metapneumovirus in Tracheal Organ Cultures of Chicken. Microorganisms. 2021; 9(11):2407. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112407
Chicago/Turabian StyleRüger, Nancy, Hicham Sid, Jochen Meens, Michael P. Szostak, Wolfgang Baumgärtner, Frederik Bexter, and Silke Rautenschlein. 2021. "New Insights into the Host–Pathogen Interaction of Mycoplasma gallisepticum and Avian Metapneumovirus in Tracheal Organ Cultures of Chicken" Microorganisms 9, no. 11: 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112407