
Non-Conventional Yeasts Whole Cells as Efficient Biocatalysts for the Production of Flavors and Fragrances

Abstract
:1. Introduction
2. Biocatalytic Production of Flavors and Fragrances by NCYs
2.1. Definition of Biocatalysis and Their Impact on the Production of Flavors and Fragrances
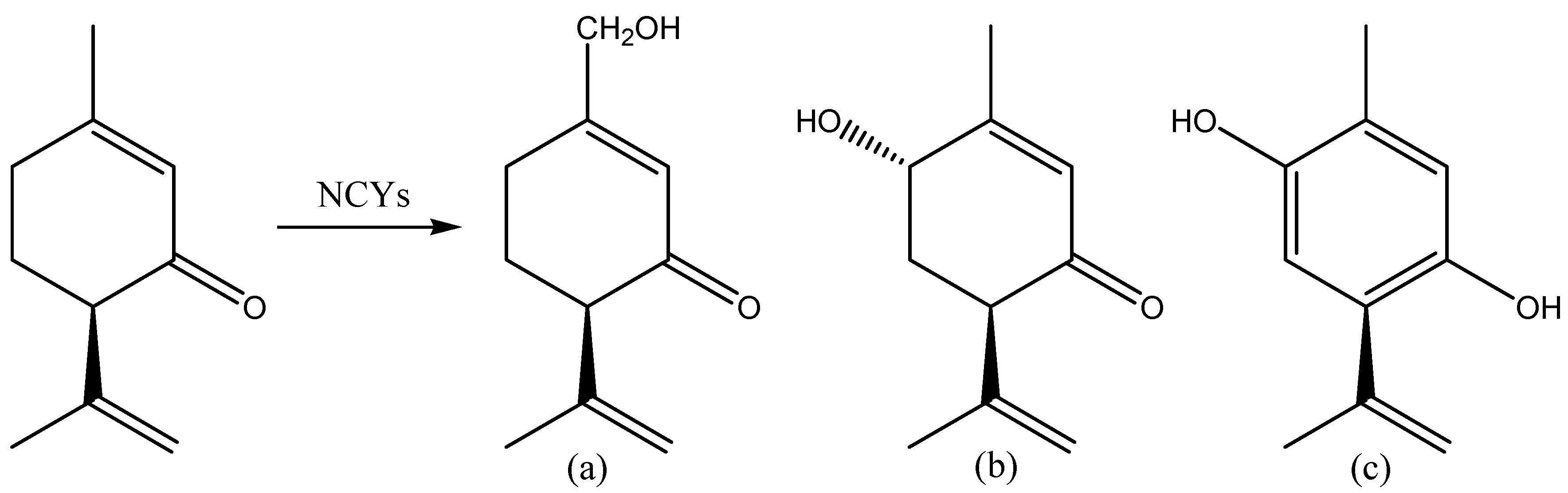
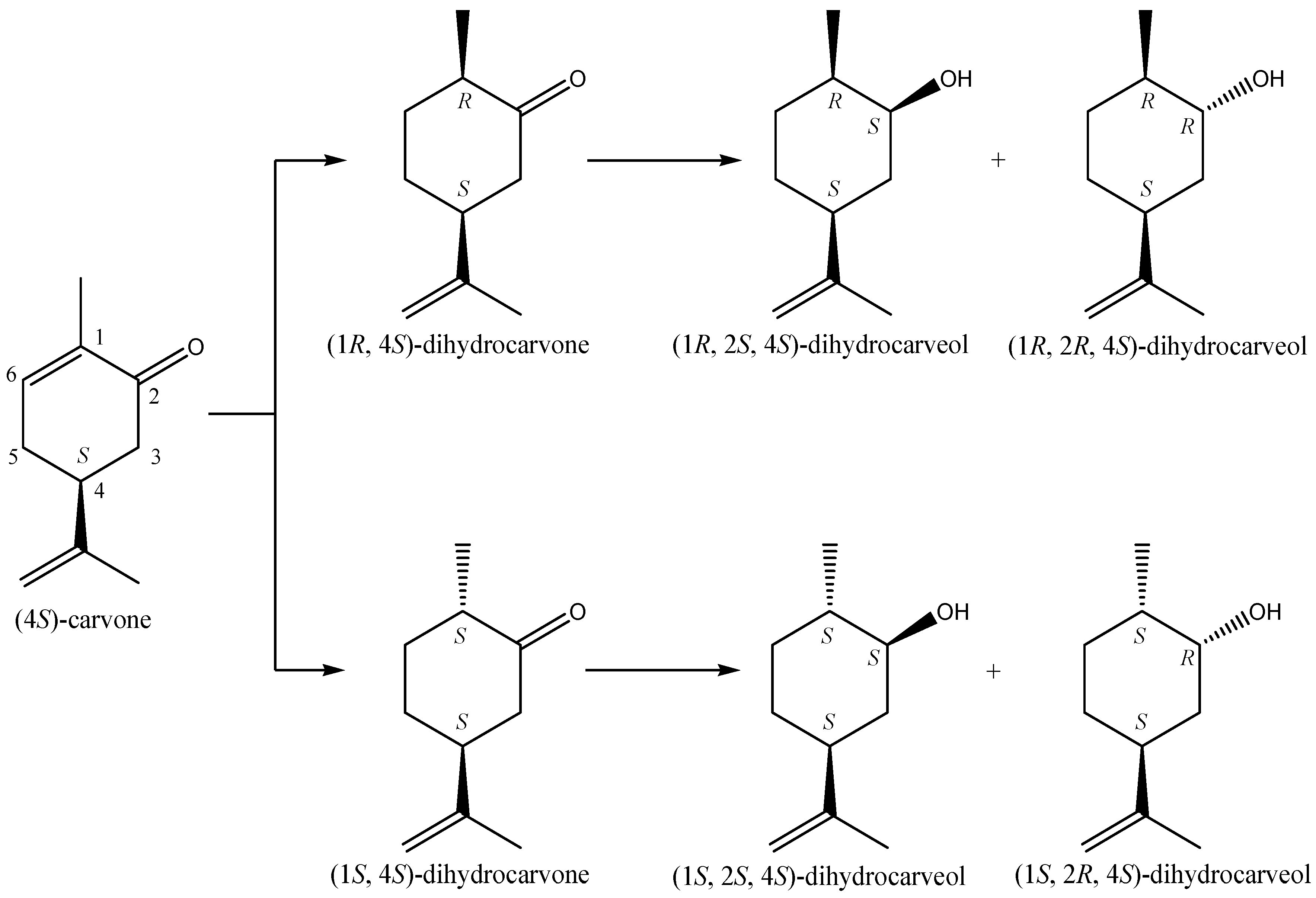
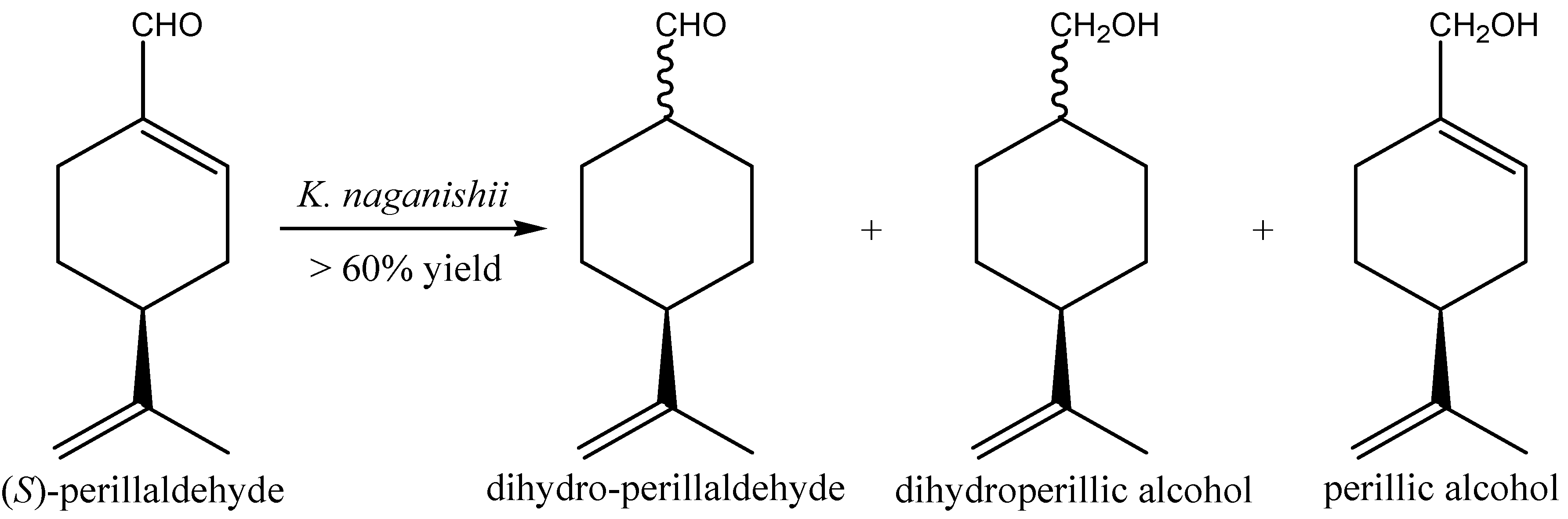
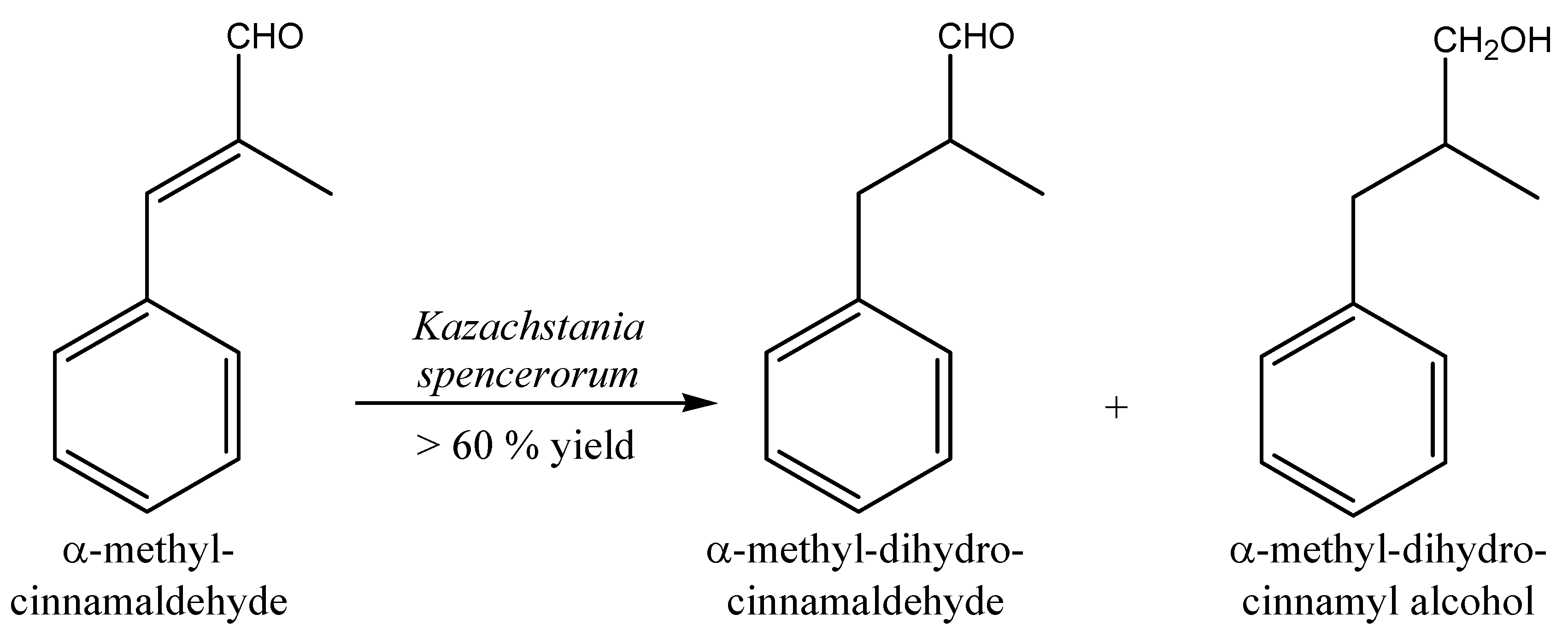
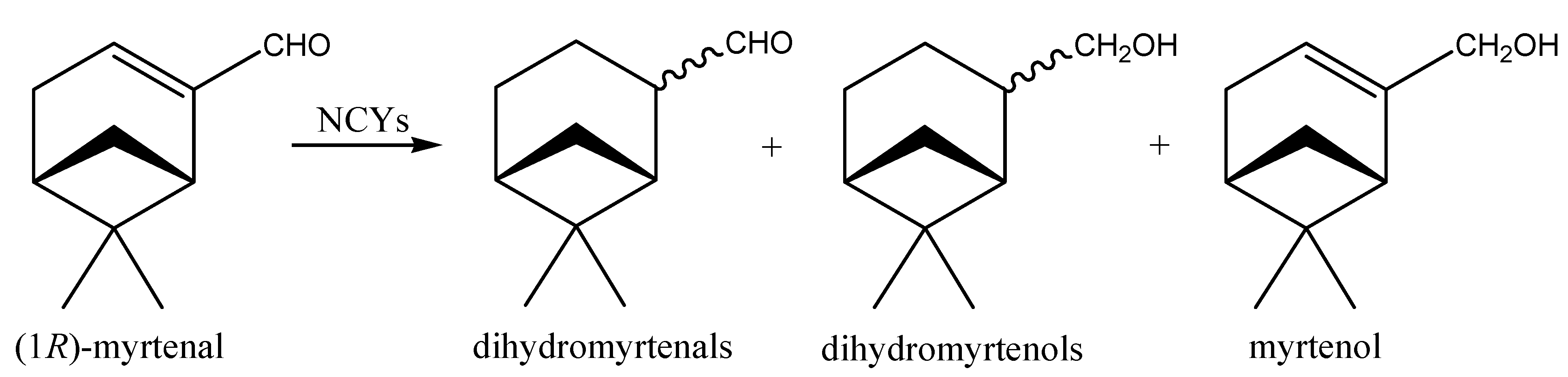
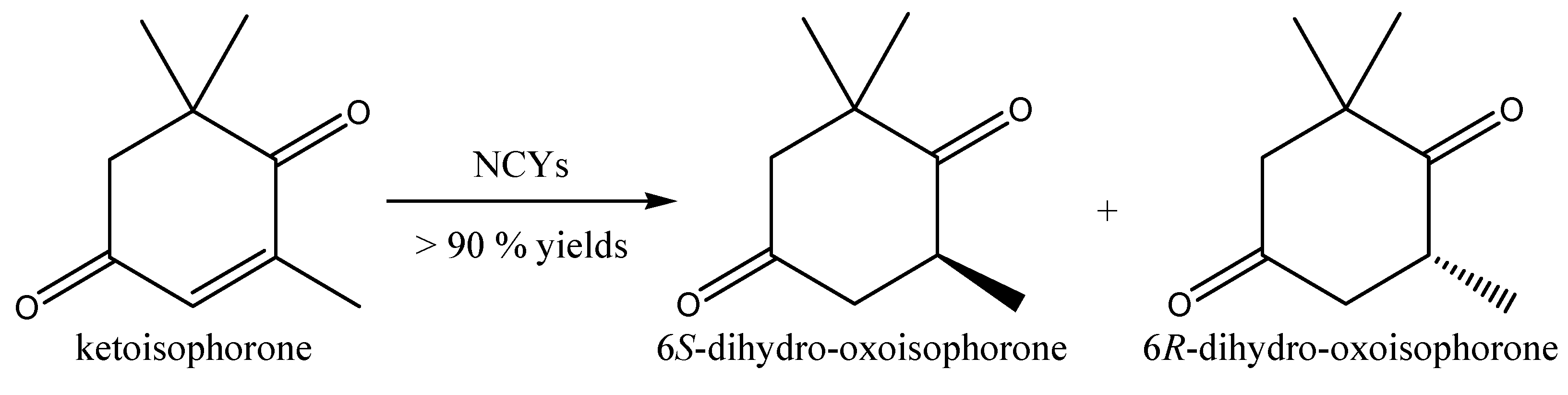
2.2. Aldehydes, Ketones and Related Compounds
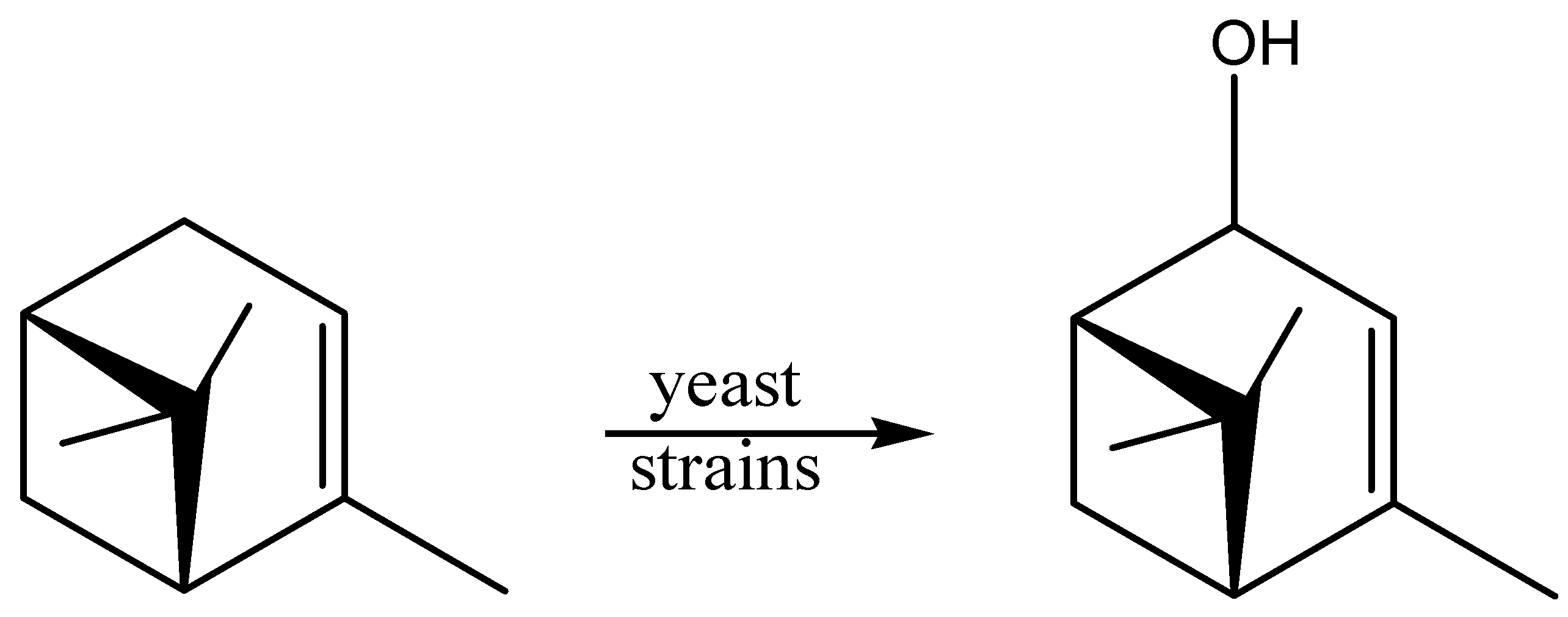
2.3. Alcohols

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2-Phenylethanol (2-PE) Production | References |
|---|---|---|
| Kluyveromyces marxianus | no more than 1.0 g/L | [29] |
| Kluyveromyces marxianus | 5.6 g/L (187% than non-optimized medium) | [30] |
| Kluyveromyces marxianus | 0.8 g/L (in bioreactor) | [31] |
| Pichia fermentans | 0.5 g/L, molar yield 71% | [32] |
| Pichia fermentans | 0.5 g/L, molar yield 74% | [33] |
2.4. Lactones
| Species | Substrate | Product | References |
|---|---|---|---|
| Candida albicans | Linoleic acid | γ-nonalactone | [55] |
| Candida tropicalis | Linoleic acid | γ-nonalactone | [55] |
| Cryptococcus laurentii | β-carotene | β-ionone, DHA | [56] |
| Lindnera saturnus (formerly Hansenula saturnus) | Ricinoleic acid | 3-hydroxylactone, decen-4-olides | [57] |
| Phaffia rhodozyma | β-carotene | β-ionone, DHA | [56] |
| Yarrowia lipolytica | Ricinoleic acid | 3-hydroxylactone, decen-4-olides | [57] |
| Yarrowia lipolytica | Ricinoleic acid | γ-decalactone | [58] |
| Yarrowia lipolytica (formerly Candida lipolytica) | Linoleic acid | γ-nonalactone | [55] |
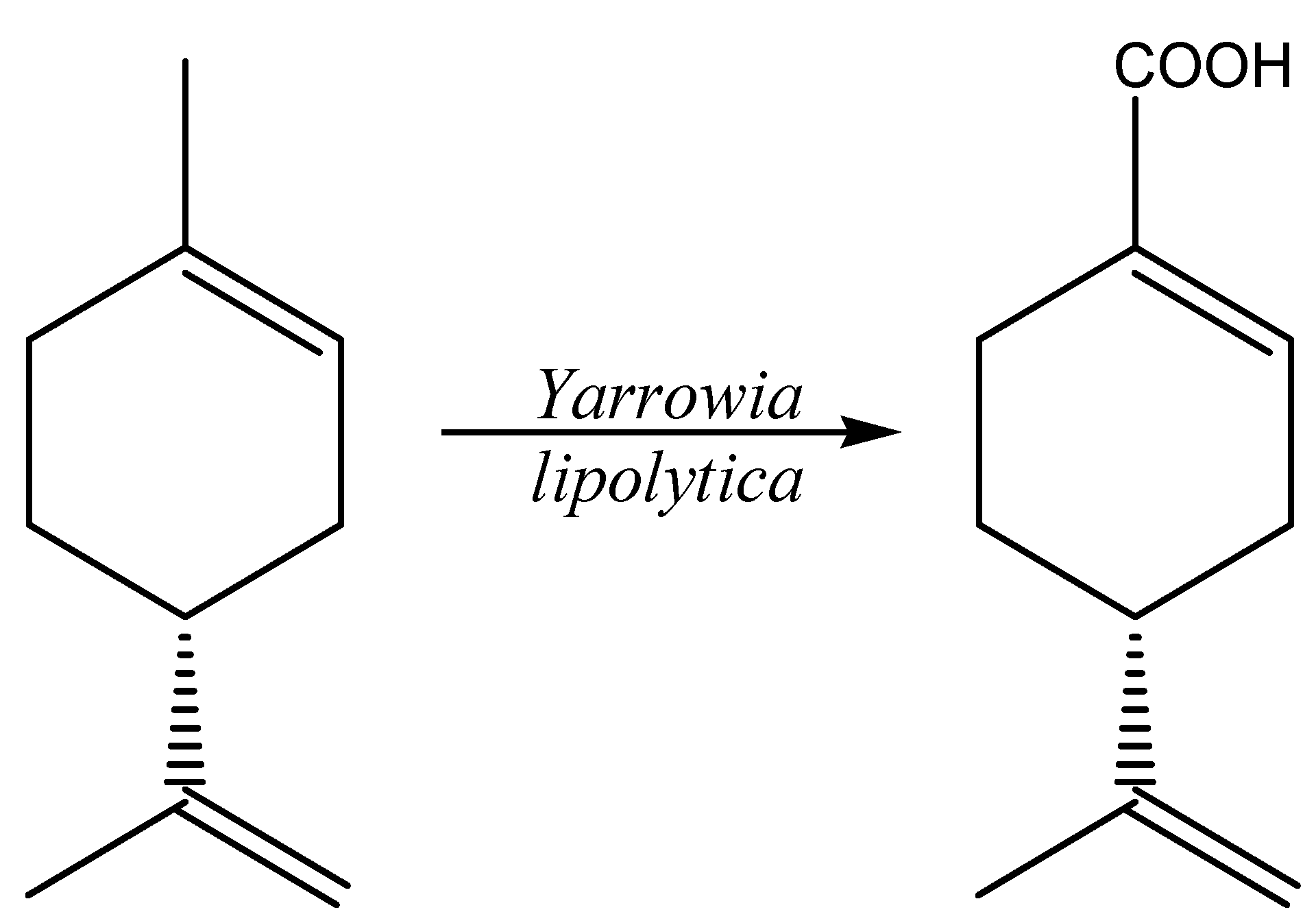
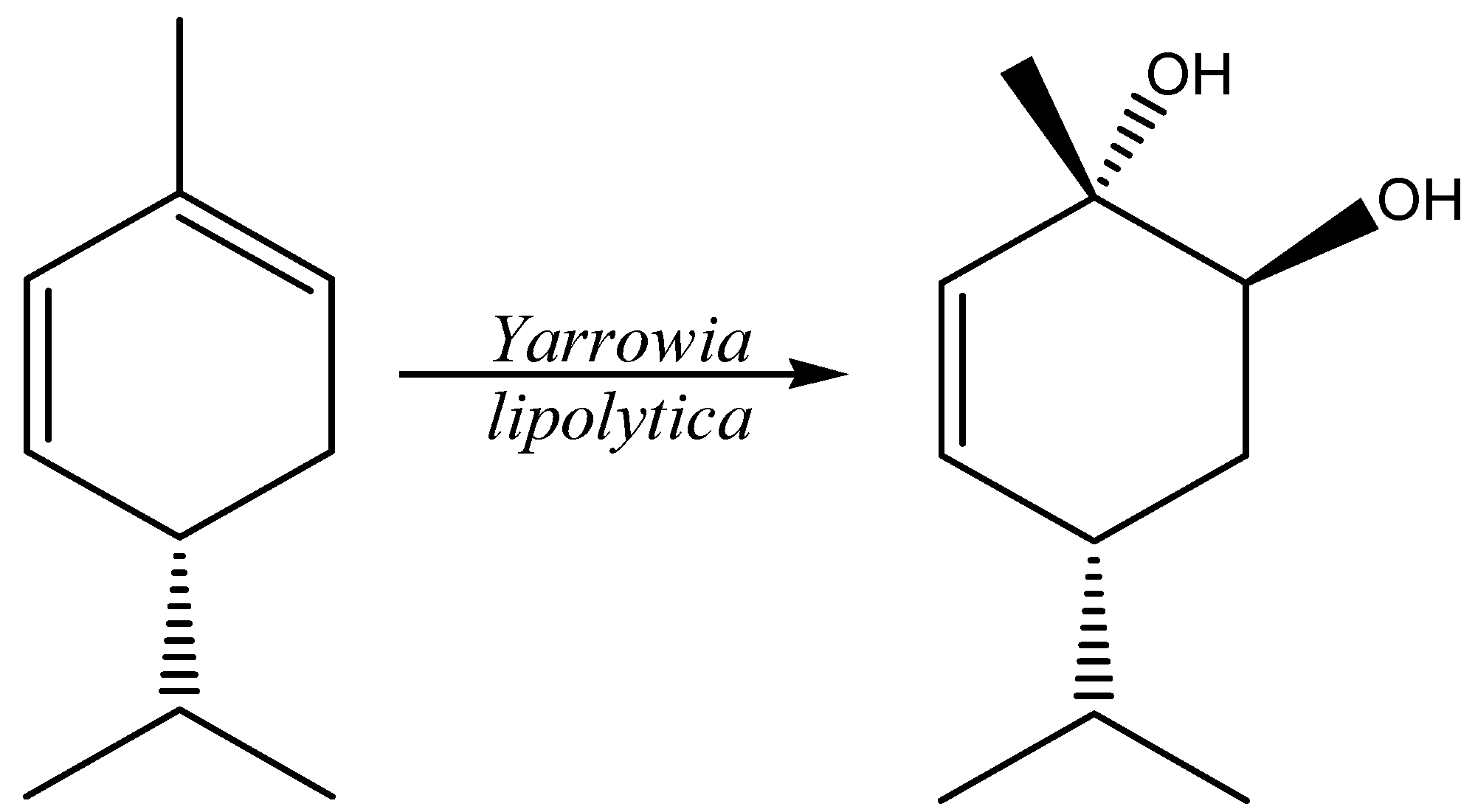
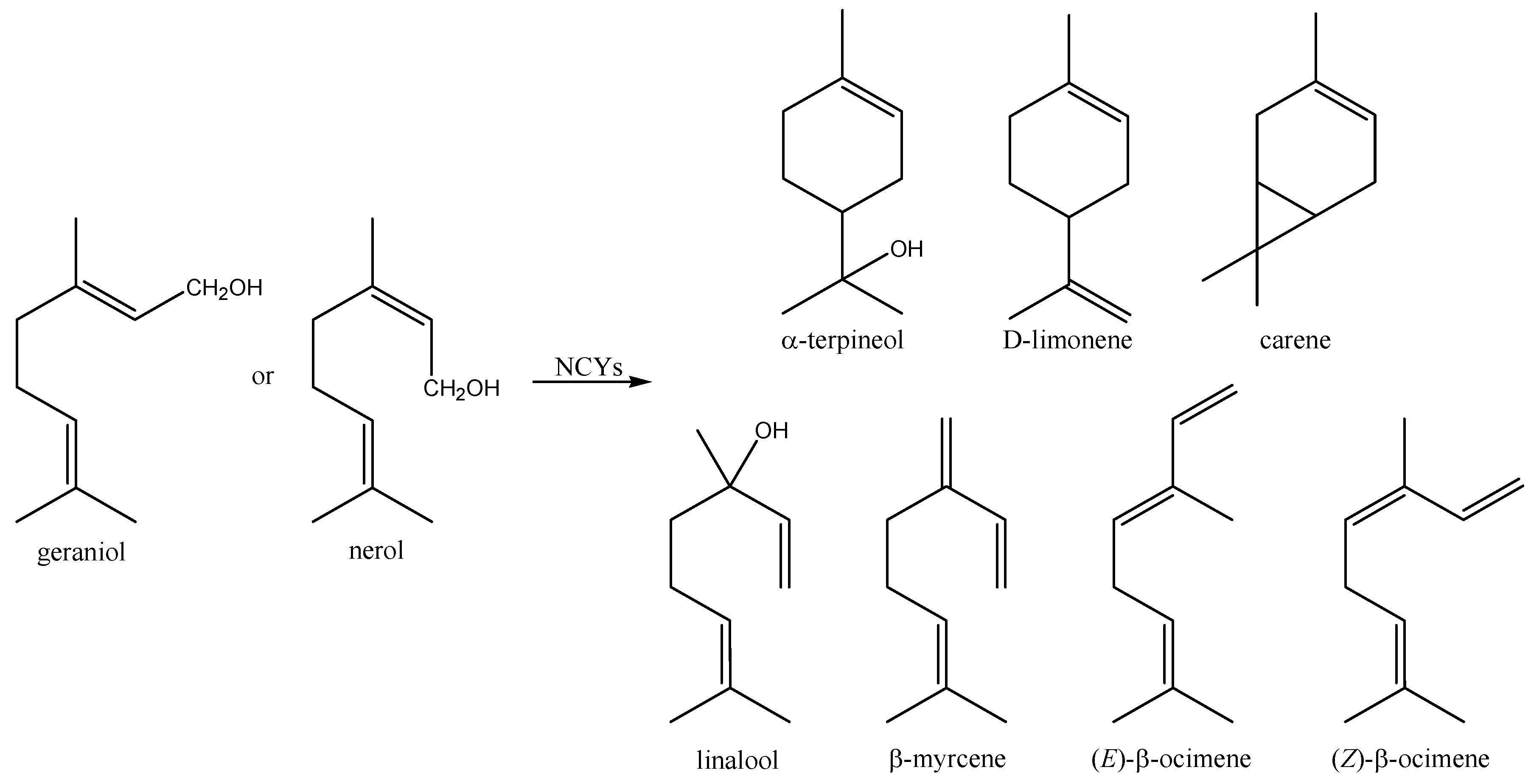
2.5. Terpenes and Terpenoids









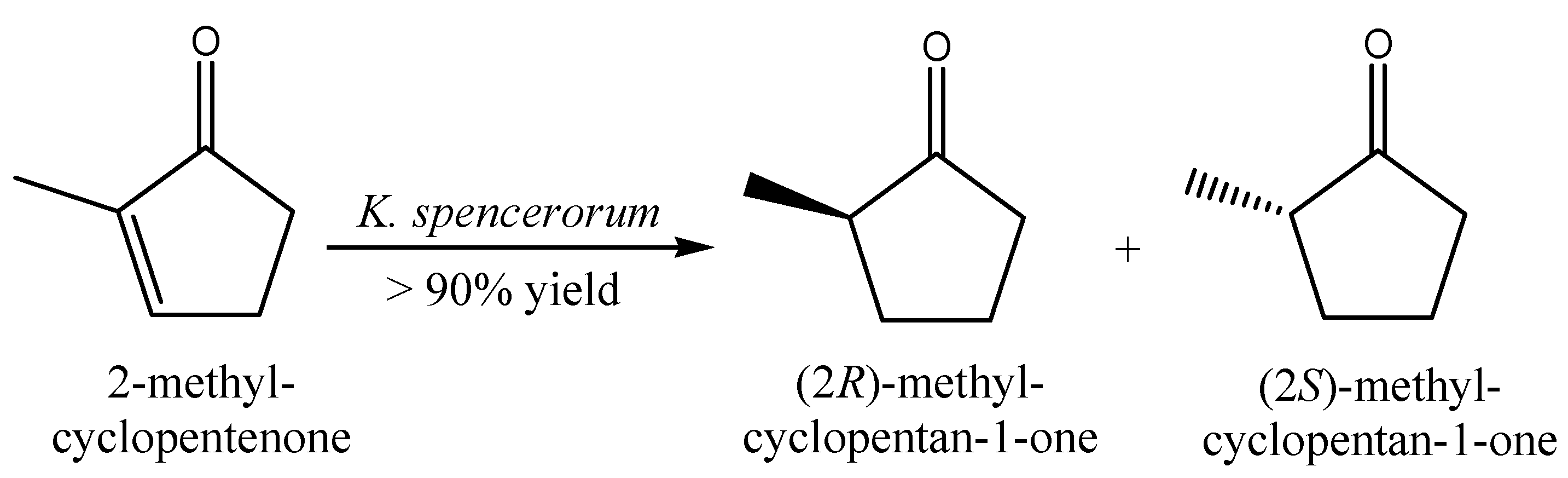
2.6. Alkenes


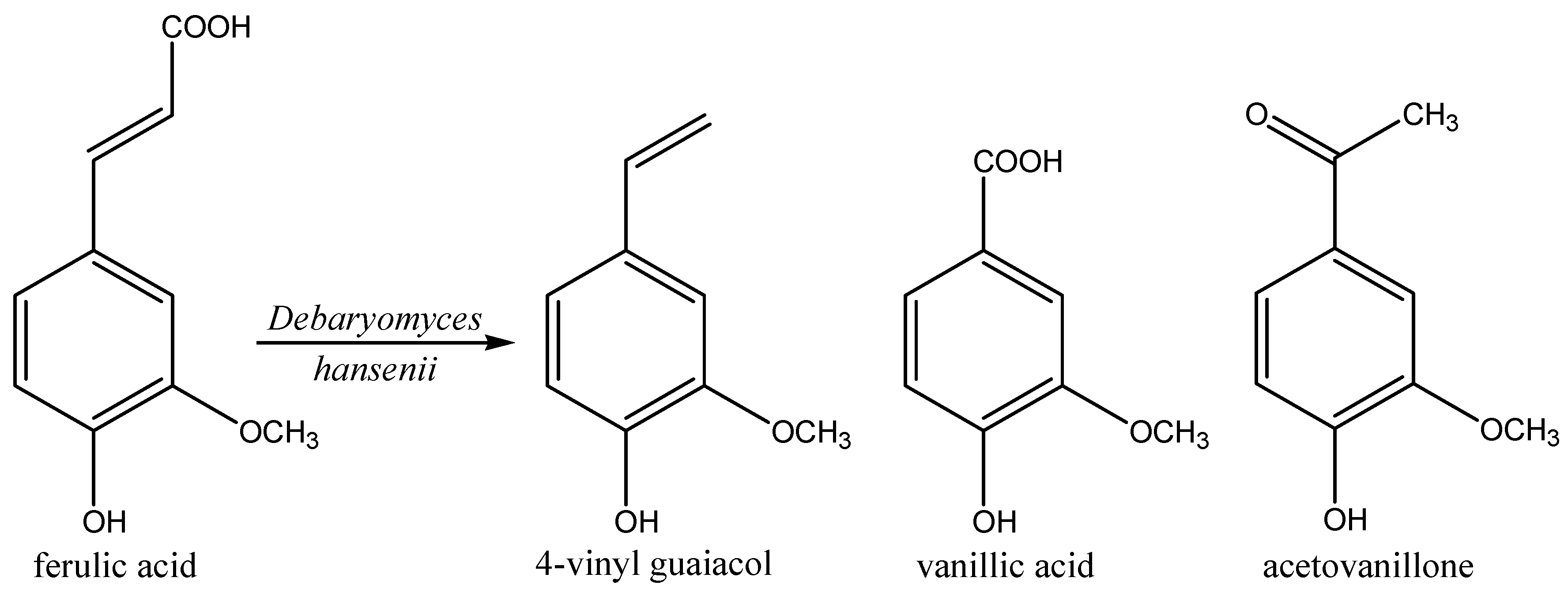
2.7. Phenols

3. Conclusions
Author Contributions
Conflicts of Interest
References
- Domínguez, Á.; Fermiñán, E.; Sánchez, M.; González, F.J.; Pérez-Campo, F.M.; García, S.; Herrero, A.B.; San Vicente, A.; Cabello, J.; Prado, M.; et al. Non-conventional yeasts as hosts for heterologous protein production. Int. Microbiol. 1998, 1, 131–142. [Google Scholar] [PubMed]
- Walker, G.M. Yeast Physiology and Biotechnology; John Wiley and Sons Ltd.: Chichester, England, UK, 1998. [Google Scholar]
- Flores, C.L.; Rodriguez, C.; Petit, T.; Gancedo, C. Carbohydrate and energy-yielding metabolism in non-conventional yeasts. FEMS Microbiol. Rev. 2000, 24, 507–529. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.A.; Echavarri-Erasun, C. Yeast biotechnology. In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 21–44. [Google Scholar]
- Hou, J.; Tyo, K.E.; Liu, Z.; Petranovic, D.; Nielsen, J. Metabolic engineering of recombinant protein secretion by Saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 491–510. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Snoek, T.; Meersman, E.; Picca Nicolino, M.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Breunig, K.; Barth, G. Non Conventional Yeasts in Genetics, Biochemistry and Biotechnology; Springer: Berlin, Germany, 2003. [Google Scholar]
- Sibirny, A.A.; Scheffers, L. Thematic section “Biochemistry, Genetics, Biotechnology and Ecology of Non-conventional Yeasts”. FEMS Yeast Res. 2002, 2, 293. [Google Scholar]
- Buzzini, P.; Vaughan-Martini, A. Yeast biodiversity and biotechnology. In Biodiversity and Ecophysiology of Yeasts; Rosa, C.A., Peter, G., Eds.; Springer: Berlin, Germany, 2006; pp. 533–559. [Google Scholar]
- The Yeasts, a Taxonomic Study; Kurtzman, C.P.; Fell, J.W.; Boekhout, T. (Eds.) Elsevier: Amsterdam, The Netherlands, 2011.
- Spencer, J.F.T.; Ragout de Spencer, A.L.; Laluce, C. Non-conventional yeasts. Appl. Microbiol. Biotechnol. 2002, 58, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Krings, U.; Berger, R.G. Biotechnological production of flavours and fragrances. Appl. Microbiol. Biotechnol. 1998, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lomascolo, A.; Stentelaire, C.; Aster, M.; Lesage-Meessen, L. Basidiomycetes as new biotechnological tools to generate natural aromatic flavours for the food industry. Trends Biotechnol. 1999, 17, 282–289. [Google Scholar] [CrossRef]
- Brenna, E.; Fuganti, C.; Gatti, F.G.; Serra, S. Biocatalytic Methods for the Synthesis of Enantioenriched Odor Active Compounds. Chem. Rev. 2011, 111, 4036–4072. [Google Scholar] [CrossRef] [PubMed]
- Straathof, A.J.J.; Panke, S.; Schmid, A. The production of fine chemicals by biotransformations. Curr. Opin. Biotechnol. 2002, 13, 548–556. [Google Scholar] [CrossRef]
- Matsuda, T.; Yamanaka, R.; Nakamura, K. Recent progress in biocatalysis for asymmetric oxidation and reduction. Tetrahedron Asymmetry 2009, 20, 513–557. [Google Scholar] [CrossRef]
- Bastos Borges, K.; de Souza Borges, W.; Durán-Patrón, R.; Tallarico Pupo, M.; Sueli Bonato, P.; González Collado, I. Stereoselective biotransformations using fungi as biocatalysts. Tetrahedron Asymmetry 2009, 20, 385–397. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis: Synthesis of key intermediates for development of pharmaceuticals. ACS Catal. 2011, 1, 1056–1074. [Google Scholar] [CrossRef]
- Faber, K. Introduction and Background Information. In Biotransformations in Organic Chemistry, 5th ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2004; pp. 3–11. [Google Scholar]
- Industrial Biotransformations, 2nd ed.; Liese, A.; Seelbach, K.; Wandrey, C. (Eds.) Wiley-VCH: Weinheim, Germany, 2006.
- Moore, J.C.; Pollard, D.J.; Kosjek, B.; Devine, P.N. Advances in the Enzymatic Reduction of Ketones. Acc. Chem. Res. 2007, 40, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Chartrain, M.; Roberge, C.; Chung, J.; McNamara, J.; Zhao, D.; Olewinski, R.; Hunt, G.; Salmon, P.; Roush, D.; Yamazaki, S.; et al. Asymmetric bioreduction of (2-(4-nitro-phenyl)-N-(2-oxo-2-pyridin-3-yl-ethyl)-acetamide) to its corresponding R alcohol [(R)-N-(2-hydroxy-2-pyridin-3-yl-ethyl)-2-(4-nitro-phenyl)-acetamide] by using Candida sorbophila MY 1833. Enzym. Microb. Technol. 1999, 25, 489–496. [Google Scholar] [CrossRef]
- Bourel, G.; Nicaud, J.M.; Nthangeni, B.; Santiago-Gomez, P.; Marc Belin, J.; Husson, F. Fatty acid hydroperoxide lyase of green bell pepper: Cloning in Yarrowia lipolytica and biogenesis of volatile aldehydes. Enzym. Microb. Technol. 2004, 35, 293–299. [Google Scholar] [CrossRef]
- Nicaud, J.M.; Madzak, C.; van den Broek, P.; Gysler, C.; Duboc, P.; Niederberger, P.; Gaillardin, C. Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res. 2002, 2, 371–379. [Google Scholar] [PubMed]
- Hua, D.; Xu, P. Recent advances in biotechnological production of 2-phenylethanol. Biotechnol. Adv. 2011, 29, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Etschmann, M.M.W.; Sell, D.; Schrader, J. Production of 2-Phenylethanol and 2-Phenylethylacetate from l-Phenylalanine by Coupling Whole-Cell Biocatalysis with Organophilic Pervaporation. Biotechnol. Bioeng. 2005, 92, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Etschmann, M.M.W.; Schrader, J. An aqueous–organic two-phase bioprocess for efficient production of the natural aroma chemicals 2-phenylethanol and 2-phenylethylacetate with yeast. Appl. Microbiol. Biotechnol. 2006, 71, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Daugulis, A.J. Bioproduction of the Aroma Compound 2-Phenylethanol in a Solid-Liquid Two-Phase Partitioning Bioreactor System by Kluyveromyces marxianus. Biotechnol. Bioeng. 2009, 104, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Fabre, C.E.; Blanc, P.J.; Goma, G. Production of 2-phenylethyl alcohol by Kluyveromyces marxianus. Biotechnol. Prog. 1998, 14, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Etschmann, M.M.W.; Sell, D.; Schrader, J. Medium optimization for the production of the aroma compound 2-phenylethanol using a genetic algorithm. J. Mol. Catal. B Enzym. 2004, 29, 187–193. [Google Scholar] [CrossRef]
- Garavaglia, J.; Flôres, S.H.; Pizzolato, T.M.; Peralba, M.C.; Ayub, M.A.Z. Bioconversion of l-phenylalanine into 2-phenylethanol by Kluyveromyces marxianus in grape must cultures. World J. Microbiol. Biotechnol. 2007, 23, 1273–1279. [Google Scholar] [CrossRef]
- Huang, C.J.; Lee, S.L.; Chou, C.C. Production and molar yield of 2-phenylethanol by Pichia fermentans L-5 as affected by some medium components. J. Biosci. Bioeng. 2000, 90, 142–147. [Google Scholar] [CrossRef]
- Huang, C.J.; Lee, S.L.; Chou, C.C. Production of 2-phenylethanol, a flavor ingredient, by Pichia fermentans L-5 under various culture conditions. Food Res. Int. 2001, 34, 277–282. [Google Scholar] [CrossRef]
- Andreu, C.; del Olmo, M. Potential of some yeast strains in the stereoselective synthesis of (R)-(−)-phenylacetylcarbinol and (S)-(+)-phenylacetylcarbinol and their reduced 1,2-dialcohol derivatives. Appl. Microbiol. Biotechnol. 2014, 98, 5901–5913. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.R.; Chari, M.A.; Narasu, M.L. Production of l-phenyl acetyl carbinol (l-PAC) by different novel strains of yeast in molasses and sugar cane juice as production medium. Res. J. Microbiol. 2006, 1, 433–437. [Google Scholar]
- Khan, M.A.; Haq, I.W.; Javed, M.M.; Quadeer, M.A.; Akhtar, N.; Bokhari, S.A.I. Studies on the production of l-phenylacetylcarbinol by Candida utilis in shake flask. Pak. J. Bot. 2012, 44, 361–364. [Google Scholar]
- Contente, M.L.; Molinari, F.; Zambelli, P.; De Vitis, V.; Gandolfi, R.; Pinto, A.; Romano, D. Biotransformation of aromatic ketones and ketoesters with the non-conventional yeast Pichia glucozyma. Tetrahedron Lett. 2014, 55, 7051–7053. [Google Scholar] [CrossRef]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Thonart, P.; Teixeira, J.A.; Destain, J.; Waché, Y. Biochemistry of lactone formation in yeast and fungi and its utilisation for the production of flavour and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef] [PubMed]
- An, J.-U.; Joo, Y.-C.; Oh, D.-K. New Biotransformation Process for Production of the Fragrant Compound γ-Dodecalactone from 10-Hydroxystearate by Permeabilized Waltomyces lipofer Cells. Appl. Environ. Microbiol. 2013, 79, 2636–2641. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Liu, Y.; Furuta, M.; Fujita, Y.; Matsumoto, T.; Fukuda, H. Preparation of high activity whole cell biocatalyst by permeabilization of recombinant flocculent yeast with alcohol. Enzym. Microb. Technol. 2000, 27, 806–811. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, C.S.; Oh, D.K. Lactulose production by betagalactosidase in permeabilized cells of Kluyveromyces lactis. Appl. Microbiol. Biotechnol. 2004, 64, 787–793. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C. Enzymatic and whole cell catalysis: Finding new strategies for old processes. Biotechnol. Adv. 2011, 29, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Farbood, M.I.; Morris, J.A.; McLean, L.B. Fermentation Process for Preparing 10-Hydroxy-C18-carboxylic Acid and Gamma-Dodecalactone Derivatives. European Patent 0578388, 12 January 1994. [Google Scholar]
- Karagül-Yüceer, Y.; Drake, M.A.; Cadwallader, K.R. Aroma-active components of non-fat dry milk. J. Agric. Food Chem. 2001, 49, 2948–2953. [Google Scholar] [CrossRef] [PubMed]
- Pino, J.A.; Marbot, R. Volatile flavor constituents of acerola (Malpighia emarginata DC.) fruit. J. Agric. Food Chem. 2001, 49, 5880–5882. [Google Scholar] [CrossRef] [PubMed]
- Herianus, J.D.L.; Zora, S.; Soon, C.T. Aroma volatiles production during fruit ripening of “Kensington Pride” mango. Postharvest Biol. Technol. 2003, 27, 323–336. [Google Scholar]
- Elss, S.; Preston, C.; Hertzig, C.; Heckel, F.; Richling, E.; Schreier, P. Aroma profiles of pineapple fruit (Ananas comosus [L.] Merr.) and pineapple products. LWT Food Sci. Technol. 2005, 38, 263–274. [Google Scholar] [CrossRef]
- Nunes, C.; Coimbra, M.A.; Saraiva, J.; Rocha, S.M. Study of the volatile components of a candied plum and estimation of their contribution to the aroma. Food Chem. 2008, 111, 897–905. [Google Scholar] [CrossRef]
- Dufossé, L.; Feron, G.; Mauvais, G.; Bonnarme, P.; Durand, A.; SPINNLER, H.E. Production of γ-Decalactone and 4-Hydroxy-Decanoic Acid in the Genus Sporidiobolus. J. Ferment. Bioeng. 1998, 86, 169–173. [Google Scholar] [CrossRef]
- Garbe, L.A.; Tressl, R. Enzymatic Baeyer-Villiger Oxidation as the Key Step in Decano-4-lactone and Decano-5-lactone Degradation by Sporobolomyces odorus. Chem. Biodiv. 2004, 1, 900–915. [Google Scholar] [CrossRef] [PubMed]
- Groguenin, A.; Waché, Y.; Escamilla-Garcia, E.; Aguedo, M.; Husson, F.; LeDall, M.T.; Nicaud, J.M.; Belin, J.M. Genetic engineering of the β-oxidation pathway in the yeast Yarrowia lipolytica to increase the production of aroma compounds. J. Mol. Catal. B Enzym. 2004, 28, 75–79. [Google Scholar] [CrossRef]
- Escamilla-Garcia, E.; Belin, J.M.; Waché, Y. Use of a Doehlert factorial design to investigate the effects of pH and aeration on the accumulation of lactones by Yarrowia lipolytica. J. Appl. Microbiol. 2007, 103, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Escamilla-Garcia, E.; Aguedo, M.; Gomes, N.; Choquet, A.; Belo, I.; Teixeira, J.A.; Belin, J.M.; Waché, Y. Production of 3-hydroxy-γ-decalactone, the precursor of two decenolides with flavouring properties, by the yeast Yarrowia lipolytica. J. Mol. Catal. B Enzym. 2009, 57, 22–26. [Google Scholar] [CrossRef]
- Braga, A.; Belo, I. Production of γ-decalactone by Yarrowia lipolytica: Insights into experimental conditions and operating mode optimization. Chem. Technol. Biotechnol. 2015, 90, 559–565. [Google Scholar] [CrossRef]
- Feron, G.; Waché, Y. Microbial biotechnology of food flavor production. In Food Biotechnology, 2nd ed.; Dominick, T., Ed.; Dekker: New York, NY, USA, 2005; pp. 407–441. [Google Scholar]
- Zorn, H.; Langhoff, S.; Scheibner, M.; Berger, R.G. Cleavage of β,β-carotene to flavor compounds by fungi. Appl. Microbiol. Biotechnol. 2003, 62, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Waché, Y.; Aguedo, M.; Choquet, A.; Gatfield, I.; Nicaud, J.M.; Belin, J.-M. Role of β-oxidation enzymes in the production of γ-decalactones from methyl ricinoleate. Appl. Environ. Microbiol. 2001, 67, 5700–5704. [Google Scholar] [PubMed]
- Waché, Y.; Aguedo, M.; Nicaud, J.M.; Belin, J.M. Catabolism of hydroxyacids and production of lactones by the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2003, 61, 393–404. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.R.; da Fonseca, M.M.R. Biotransformation of terpenes. Biotechnol. Adv. 2006, 24, 134–142, and references cited herein. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T. Biotransformation of terpenoids by mammals, microorganisms, and plant-cultured cells. Chem. Biodivers. 2005, 2, 569–590. [Google Scholar] [CrossRef] [PubMed]
- Van Rensburg, E.; Moleleki, N.; van der Walt, J.P.; Botes, P.J.; van Dyk, M.S. Biotransformation of (+)-limonene and (−)-piperitone by yeasts and yeast-like fungi. Biotechnol. Lett. 1997, 19, 779–782. [Google Scholar] [CrossRef]
- Pinder, A.R. The Chemistry of the Terpenes; Wiley: New York, NY, USA, 1960. [Google Scholar]
- Gomes, M., Jr.; Antunes, O.A.C. Upjohn Catalytic Osmium Tetroxide Oxidation Process: Diastereoselective Dihydroxylation of Monoterpenes. Catal. Commun. 2001, 2, 225–227. [Google Scholar] [CrossRef]
- Da Silva, M.J.; Robles-Deutenhefner, P.; Menini, L.; Gusevskaya, E.V. Cobalt catalyzed autoxidation of monoterpenes in acetic acid and acetonitrile solutions. J. Mol. Catal. A Chem. 2003, 201, 71–77. [Google Scholar] [CrossRef]
- Rottava, I.; Cortina, P.F.; Zanella, C.A.; Cansian, R.L.; Toniazzo, G.; Treichel, H.; Antunes, O.A.C.; Oestreicher, E.G.; de Oliveira, D. Microbial oxidation of (−)-α-pinene to verbenol production by newly isolated strains. Appl. Biochem. Biotechnol. 2010, 162, 2221–2231. [Google Scholar] [CrossRef] [PubMed]
- Javidnia, K.; Aram, F.; Solouki, M.; Reza Mehdiopour, A.; Gholami, M.; Miri, R. Microbial biotransformation of some monoterpene hydrocarbons. Ann. Microbiol. 2009, 59, 349–351. [Google Scholar] [CrossRef]
- Işcan, G.; Kırımer, N.; Demirci, F.; Demirci, B.; Noma, Y.; Başer, K.H.C. Biotransformation of (−)-(R)-α-Phellandrene: Antimicrobial Activity of Its Major Metabolite. Chem. Biodivers. 2012, 9, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Lemos, B.J.; Dionsio, A.P.; Pastore, G.M. Bio-oxidation of terpenes: An approach for the flavor industry. Chem. Rev. 2009, 109, 4518–4531. [Google Scholar]
- Van Dyk, M.S.; van Rensburg, E.; Rensburg, I.P.B.; Moleleki, N. Biotransformation of monoterpenoid ketones by yeasts and yeast-like fungi. J. Mol. Catal. B Enzym. 1998, 5, 149–154. [Google Scholar] [CrossRef]
- Goretti, M.; Ponzoni, C.; Caselli, E.; Marchigiani, E.; Cramarossa, M.R.; Turchetti, B.; Buzzini, P.; Forti, L. Biotransformation of electron-poor alkenes by yeasts: Asymmetric reduction of (4S)-(+)-carvone by yeast enoate reductases. Enzym. Microb. Technol. 2009, 45, 463–468. [Google Scholar] [CrossRef]
- Goretti, M.; Branda, E.; Turchetti, B.; Cramarossa, M.R.; Onofri, A.; Forti, L.; Buzzini, P. Response surface methodology as optimization strategy for asymmetric bioreduction of (4S)-(+)-carvone by Cryptococcus gastricus. Bioresour. Technol. 2012, 121, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Carballeira, J.D.; Valmaseda, M.; Alvarez, E.; Sinisterra Gago, J.V. Gongronella butleri, Schizosaccharomyces octosporus and Diplogelasinospora grovesii: Novel microorganisms useful for the stereoselective reduction of ketones. Enzym. Microb. Technol. 2004, 34, 611–623. [Google Scholar] [CrossRef]
- Goretti, M.; Turchetti, B.; Cramarossa, M.R.; Forti, L.; Buzzini, P. Production of flavours and fragrances via bioreduction of (4R)-(−)-carvone and (1R)-(−)-myrtenal by non-conventional yeast whole-cells. Molecules 2013, 18, 5736–5748. [Google Scholar] [CrossRef] [PubMed]
- Goretti, M.; Ponzoni, C.; Caselli, E.; Marchegiani, E.; Cramarossa, M.R.; Turchetti, B.; Forti, L.; Buzzini, P. Bioreduction of α,β-unsaturated ketones and aldehydes by non-conventional yeast (NCY) whole-cells. Bioresour. Technol. 2011, 102, 3993–3998. [Google Scholar] [CrossRef] [PubMed]
- Noma, Y.; Takahashi, H.; Asakawa, Y. Biotransformation of terpene aldehydes by Euglena gracilis Z. Phytochemistry 1991, 30, 1147–1151. [Google Scholar] [CrossRef]
- Bhatia, S.P.; McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance material review on myrtenol. Food Chem. Toxicol. 2008, 46, S237–S240. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Dickinson, J.R. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Ponzoni, C.; Gasparetti, C.; Goretti, M.; Turchetti, B.; Cramarossa, M.R.; Pagnoni, U.M.; Forti, L.; Buzzini, P. Biotransformation of acyclic monoterpenoids by Debaryomyces sp., Kluyveromyces sp. and Pichia sp. strains of environmental origin. Chem. Biodivers. 2008, 5, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.K.; Tasnádia, G.; Clay, D.; Hall, M.; Faber, K. Asymmetric bioreduction of activated alkenes to industrially relevant optically active compounds. J. Biotechnol. 2012, 162, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Yanto, Y.; Bommarius, A.S. Enzymes, Enoate Reductases “Old Yellow Enzyme”. In Encyclopedia of Industrial Biotechnology; John Wiley & Sons: Hoboken, NJ, USA, 2010; pp. 1–14. [Google Scholar]
- Toogood, H.S.; Gardiner, J.M.; Scrutton, N.S. Biocatalytic reductions and chemical versatility of the old yellow enzyme family of flavoprotein oxidoreductases. ChemCatChem 2010, 2, 892–914. [Google Scholar] [CrossRef]
- Stuermer, R.; Hauer, B.; Hall, M.; Faber, K. Asymmetric bioreduction of activated C=C bonds using enoate reductases from the old yellow enzyme family. Curr. Opin. Chem. Biol. 2007, 11, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Stueckler, C.; Kroutil, W.; Macheroux, P.; Faber, K. Asymmetric bioreduction of activated alkenes using cloned 12-oxophytodienoate reductase isoenzymes OPR-1 and OPR-3 from Lycopersicon esculentum (Tomato): A striking change of stereoselectivity. Angew. Chem. Int. Ed. 2007, 46, 3934–3937. [Google Scholar] [CrossRef] [PubMed]
- Blehert, D.S.; Fox, B.G.; Chambliss, G.H. Cloning and sequence analysis of two Pseudomonas flavoprotein xenobiotic reductases. J. Bacteriol. 1999, 181, 6254–6263. [Google Scholar] [PubMed]
- French, C.E.; Nicklin, S.; Bruce, N.C. Sequence and properties of pentaerythritol tetranitrate reductase from Enterobacter cloacae PB2. J. Bacteriol. 1996, 178, 6623–6627. [Google Scholar] [PubMed]
- French, C.E.; Bruce, N.C. Purification and characterization of morphinone reductase from Pseudomonas putida M10. Biochem. J. 1994, 301, 97–103. [Google Scholar] [PubMed]
- Miranda, M.; Ramirez, J.; Guevara, S.; Ongay-Larios, L.; Pena, A.; Coria, R. Nucleotide sequence and chromosomal localization of the gene encoding the Old Yellow Enzyme from Kluyveromyces lactis. Yeast 1995, 11, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Niino, Y.S.; Chakraborty, S.; Brown, B.J.; Massey, V. A new Old Yellow Enzyme of Saccharomyces cerevisiae. J. Biol. Chem. 1995, 270, 1983–1991. [Google Scholar] [PubMed]
- Komduur, J.A.; Leão, A.N.; Monastyrska, I.; Veenhuis, M.; Kiel, J.A. Old yellow enzyme confers resistance of Hansenula polymorpha towards allyl alcohol. Curr. Genet. 2002, 41, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Quezada, M.A.; Carballeira, J.D.; Sinisterra, J.V. Monascus kaoliang CBS 302.78 immobilized in polyurethane foam using iso-propanol as co-substrate: Optimized immobilization conditions of a fungus as biocatalyst for the reduction of ketones. Bioresour. Technol. 2009, 100, 2018–2025. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, M.; Kotaka, A.; Thiwthong, R.; Wada, M.; Nakamori, S.; Shimizu, S. Cloning and over expression of the Old Yellow Enzyme gene of Candida macedoniensis and its application to the production of a chiral compound. J. Biotechnol. 2004, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carballeira, J.D.; Quezada, M.A.; Hoyos, P.; Simeó, Y.; Hernaiz, M.J.; Alcantara, A.R.; Sinisterra, J.V. Microbial cells as catalysts for stereoselective redox reactions. Biotechnol. Adv. 2009, 27, 686–714. [Google Scholar] [CrossRef] [PubMed]
- Buque-Taboada, E.M.; Straathof, A.J.J.; Heijnen, J.J.; van der Wielen, L.A.M. Substrate inhibition and product degradation during the reduction of 4-oxoisophorone by Saccharomyces cerevisiae. Enzym. Microb. Technol. 2005, 37, 625–633. [Google Scholar] [CrossRef]
- Faber, K. Biocatalytic Applications. In Biotransformations in Organic Chemistry, 5th ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2004; pp. 205–212. [Google Scholar]
- Wanner, P.; Tressl, R. Purification and characterization of two enone reductases from Saccharomyces cerevisiae. Eur. J. Biochem. 1998, 255, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Sode, K.; Kajiwara, K.; Tamiya, E.; Karube, I. Continuous asymmetric reduction of 4-oxoisophorone by thermophilic bacteria using a hollow fiber reactor. Biocatal. Biotransform. 1987, 1, 77–86. [Google Scholar] [CrossRef]
- Nishimura, O.; Mihara, S.; Shibamoto, T. Compounds produced by the reaction of 2-hydroxy-3-methyl-2-cyclopenten-1-one with ammonia and hydrogen sulfide. J. Agric. Food Chem. 1980, 28, 39–43. [Google Scholar] [CrossRef]
- Middelhoven, W.J.; Koorevaar, M.; Schuur, G. Degradation of benzene compounds by yeasts in acidic soils. Plant Soil 1992, 145, 37–43. [Google Scholar] [CrossRef]
- Priefert, H.; Rabenhorst, J.; Steinbüchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Shanker, K.S.; Kishore, K.H.; Kanjilal, S.; Misra, S.; Murty, U.S.N.; Prasad, R.B.N. Biotransformation of ferulic acid to acetovanillone using Rhizopus oryzae. Biocatal. Biotransform. 2007, 25, 109–112. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E.; Sudheesh, S. Rapid conversion of ferulic acid to 4-vinyl guaiacol and vanillin metabolites by Debaryomyces hansenii. J. Mol. Catal. B Enzym. 2007, 44, 48–52. [Google Scholar] [CrossRef]
- Max, B.; Tugores, F.; Cortés-Diéguez, S.; Domínguez, J.M. Bioprocess design for the microbial production of natural phenolic compounds by Debaryomyces hansenii. Appl. Biochem. Biotechnol. 2012, 168, 2268–2284. [Google Scholar] [CrossRef] [PubMed]
- Donaghy, J.A.; Kelly, P.F.; McKay, A. Conversion of ferulic acid to 4-vinyl guaiacol by yeasts isolated from unpasteurised apple juice. Sci. Food Agric. 1999, 79, 453–456. [Google Scholar] [CrossRef]
- Antoniotti, S. Tuning of essential oil properties by enzymatic treatment: Toward sustainable processes for the generation of new fragrance ingredients. Molecules 2014, 19, 9203–9214. [Google Scholar] [CrossRef] [PubMed]
- Schalk, M.; Pastore, L.; Mirata, M.A.; Khim, S.; Schouwey, M.; Deguerry, F.; Pineda, V.; Rocci, L.; Daviet, L. Toward a biosynthetic route to sclareol and amber odorant. J. Am. Chem. Soc. 2012, 134, 18900–18903. [Google Scholar] [CrossRef] [PubMed]
- Patchouli and Synthetic Biology. A Case Study. Available online: http://www.etcgroup.org/sites/www.etcgroup.org/files/ETC-patchouli-synbio-casestudy2014.pdf (accessed on 25 May 2015).
- Ohloff, G.; Pickenhagen, W.; Kraft, P. Scent and Chemistry. The Molecular World of Odors; Verlag Helvetica Chimica Acta: Zurich, Switzerland, 2011; p. 257. Available online: http://www.wiley-vch.de/books/sample/3906390667_erratum.pdf (accessed on 25 May 2015).
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forti, L.; Di Mauro, S.; Cramarossa, M.R.; Filippucci, S.; Turchetti, B.; Buzzini, P. Non-Conventional Yeasts Whole Cells as Efficient Biocatalysts for the Production of Flavors and Fragrances. Molecules 2015, 20, 10377-10398. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610377
Forti L, Di Mauro S, Cramarossa MR, Filippucci S, Turchetti B, Buzzini P. Non-Conventional Yeasts Whole Cells as Efficient Biocatalysts for the Production of Flavors and Fragrances. Molecules. 2015; 20(6):10377-10398. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610377
Chicago/Turabian StyleForti, Luca, Simone Di Mauro, Maria Rita Cramarossa, Sara Filippucci, Benedetta Turchetti, and Pietro Buzzini. 2015. "Non-Conventional Yeasts Whole Cells as Efficient Biocatalysts for the Production of Flavors and Fragrances" Molecules 20, no. 6: 10377-10398. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610377