
The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Exposure to Feed-Borne ZEN
2.1. Presence of ZEN in Feeds
2.2. Eryptosis—One of the First Consequences of Exposure to ZEN
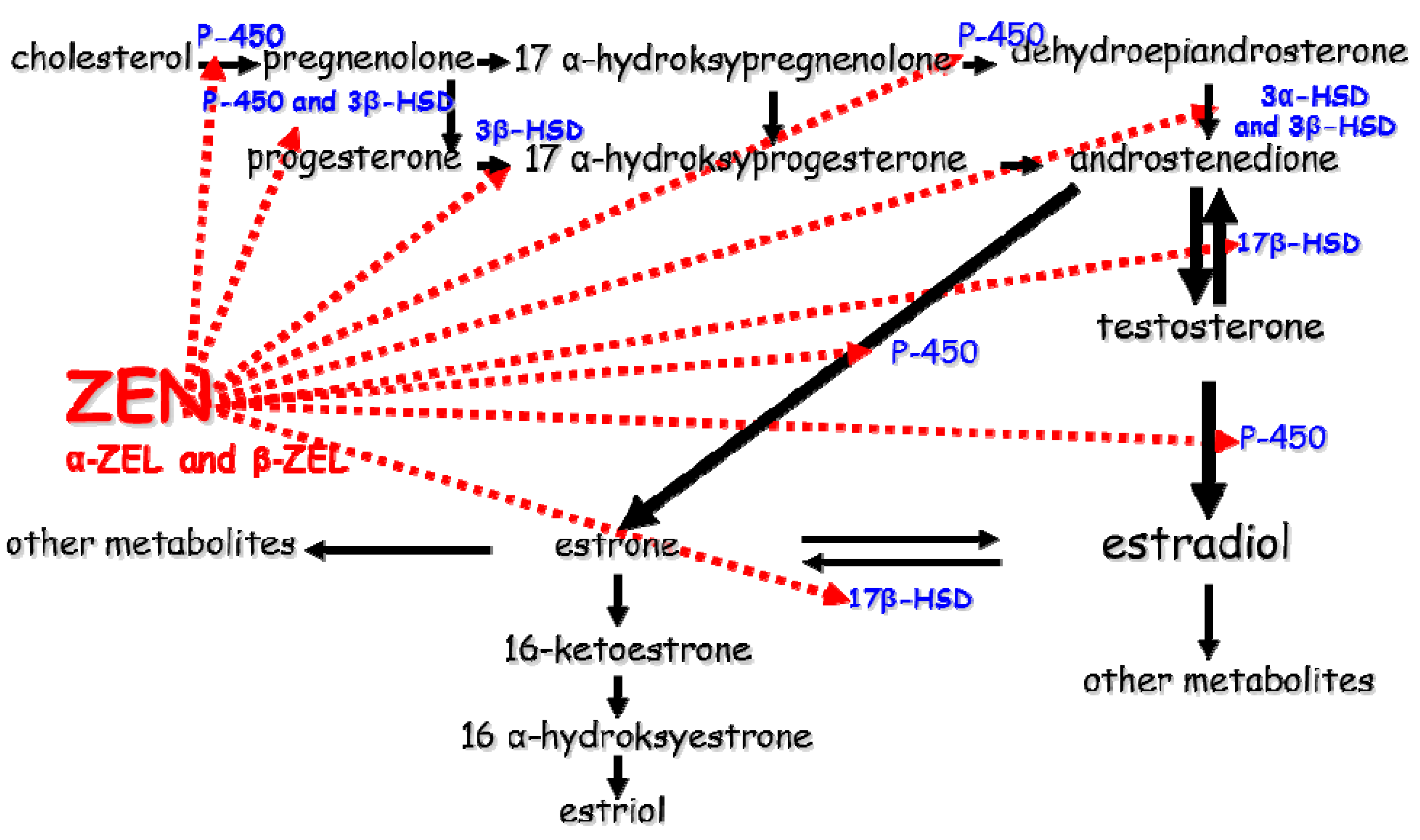
2.3. Disrupted Hormone Secretion during Exposure to ZEN
Involvement of Selected Steroid Hormones—Estradiol (E2) and Progesterone (P4)
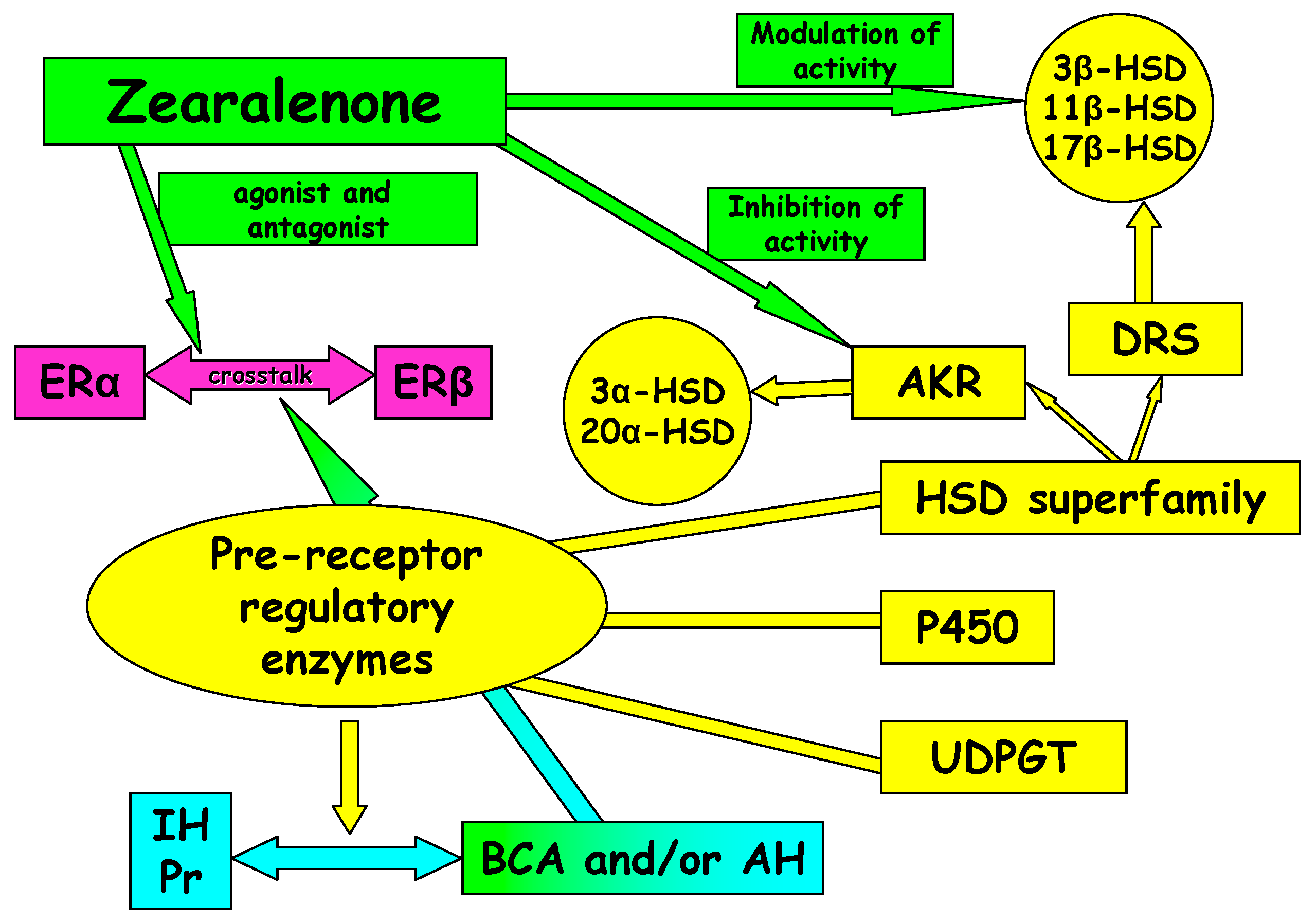
2.4. Involvement of Selected Hydroxysteroid Dehydrogenases (HSDs)


3. Oestrogen Receptors
3.1. Ovaries
3.2. Uterus
4. Metabolic Activity
4.1. Ovaries
4.2. Uterus
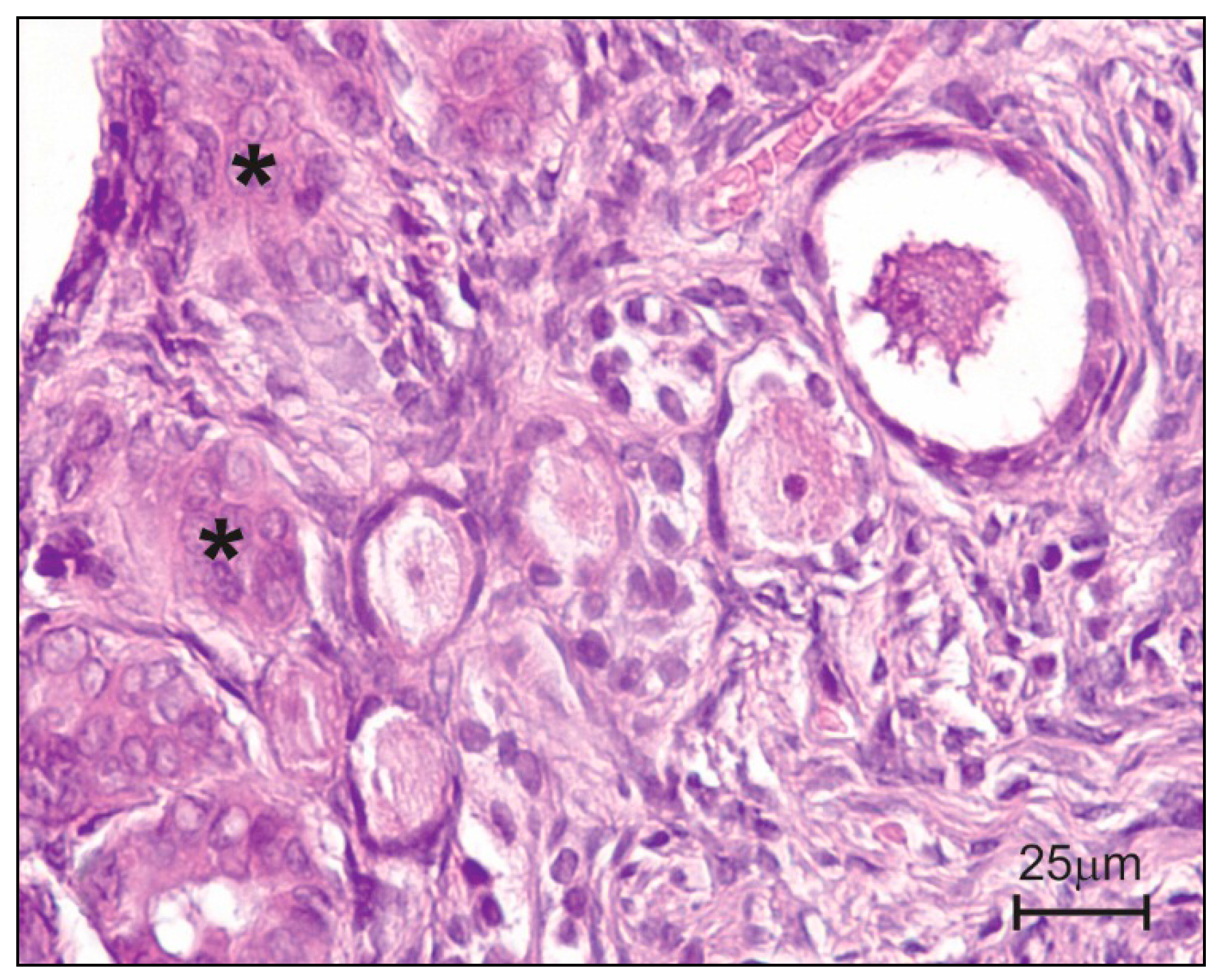
5. Histopathology of Selected Reproductive Structure
5.1. Ovaries

5.2. Uterus
6. Summary

7. Conclusions
- -
- exposure to ZEN promotes changes in the metabolic profile of the reproductive system, which leads to adverse reproductive outcomes, such as follicular atresia,
- -
- selected laboratory tests, such as the HSD activity assay, can be used to determine the presence of ZEN or predict pathological states in the affected tissues,
- -
- modulation of HSD activity could be an effective measure to reduce the risk of ZEN intoxication,
- -
- NOAEL doses of ZEN and its metabolites have a stimulatory effect on pre-pubertal bitches, whereas above NOAEL doses could inhibit vital life processes,
- -
- the reviewed publications do not provide a clear answer as to whether the observed changes in the health status of animals are indicative of mycotoxicosis or result merely from exposure to ZEN.
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses. Endocr. Rev. 2012, 33, 378–445. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, B.; Patel, M.; Fahey, J.; Wira, C. Endocrine control of mucosal immunity in the female reproductive tract: Impact of environmental disruptors. Mol. Cell. Endocrinol. 2012, 354, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Kolle, S.N.; Ramirez, T.; Kamp, H.G.; Buesen, R.; Flick, B.; Strauss, V.; van Ravenzwaay, B. A testing strategy for the identification of mammalian, systemic endocrine disruptors with particular focus on steroids. Regul. Toxicol. Pharmacol. 2012, 63, 259–278. [Google Scholar] [CrossRef] [PubMed]
- Marchais-Oberwinkler, S.; Henn, C.; Moller, G.; Klein, T.; Negri, M.; Oster, A.; Spadaro, A.; Werth, R.; Wetzel, M.; Xu, K.; et al. 17β-Hydroxysteroid dehydrogenases (17β-HSDs) as therapeutic targets: Protein structures, functions, and recent progress in inhibitor development. J. Steroid Biochem. 2011, 125, 66–82. [Google Scholar] [CrossRef] [PubMed]
- Chighizola, C.; Meroni, P.L. The role of environmental oestrogens and autoimmunity. Autoimmun. Rev. 2012, 11, A493–A501. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.J. Longevity in pet dogs: Understanding what’s missing. Vet. J. 2014, 200, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Escriva, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Obremski, K.; Jakimiuk, E.; Skorska-Wyszyńska, E.; Zielonka, Ł.; Gajęcki, M. Histopathological examination of ovaries in bitches after experimental zearalenone micotoxicosis. Pol. J. Vet. Sci. 2008, 11, 363–366. [Google Scholar] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Embry, M.R.; Bachman, A.N.; Bell, D.R.; Boobis, A.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doerrer, N.G.; Hines, R.N.; Moretto, A.; et al. Risk assessment in the 21st century: Roadmap and matrix. Crit. Rev. Toxicol. 2014, 44, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Pelinescu, D. Induction of pro-inflammatory gene expression by Escherichia coli and mycotoxin zearalenone contamination and protection by a Lactobacillus mixture in porcine IPEC-1 cells. Toxicon 2015, 97, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Zachariasova, M.; Dzumana, Z.; Veprikova, Z.; Hajkovaa, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feedingstuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- De Saeger, S.; van Egmond, H.P. Special issue: Masked mycotoxins. World Mycotoxin J. 2012, 5, 203–206. [Google Scholar] [CrossRef]
- Słomczyńska, M. The effect of phytoestrogens on the reproductive tract. Pol. J. Vet. Sci. 2004, 7, 223–226. [Google Scholar] [PubMed]
- Zielonka, Ł.; Gajęcka, M.; Rozicka, A.; Dąbrowski, M.; Żmudzki, J.; Gajęcki, M. The effect of environmental mycotoxins on selected ovarian tissue fragments of multiparous female wild boars at the beginning of astronomical winter. Toxicon 2014, 89, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panini, N.V.; Salinas, E.; Messina, G.A.; Raba, J. Modified paramagnetic beads in a microfluidic system for the determination of zearalenone in feedstuffs samples. Food Chem. 2011, 125, 791–796. [Google Scholar] [CrossRef]
- Cortinovis, C.; Caloni, F.; Schreiber, N.B.; Spicer, L.J. Effects of fumonisin B1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology 2014, 81, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Stopa, E.; Gajęcka, M.; Babińska, I.; Zielonka, Ł.; Gajęcki, M. The effect of experimental exposure to low doses of zearalenone on uterine histology and morphometry in prepubertal bitches. Theriogenology 2014, 82, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Concannon, P.W. Reproductive cycles of the domestic bitch. Anim. Reprod. Sci. 2011, 124, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Mir, F.; Fontaine, E.; Albaric, O.; Greer, M.; Vannier, F.; Schlafer, D.H.; Fontbonne, A. Findings in uterine biopsies obtained by laparotomy from bitches with unexplained infertility or pregnancy loss: An observational study. Theriogenology 2013, 79, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Boermans, H.J.; Leung, M.C. Mycotoxins and the pet food industry: Toxicological evidence and risk assessment. Int. J. Food Microbiol. 2007, 119, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Gazzotti, T.; Biagi, G.; Pagliuca, G.; Pinna, C.; Scardilli, M.; Grandi, M.; Zaghini, G. Occurrence of mycotoxins in extruded commercial dog food. Anim. Feed Sci. Technol. 2015, 202, 81–89. [Google Scholar] [CrossRef]
- Zwierzchowski, W.; Gajęcki, M.; Obremski, K.; Zielonka, Ł.; Baranowski, M. The occurrence of zearalenone and its derivatives in standard and therapeutic feeds for companion animals. Pol. J. Vet. Sci. 2004, 7, 289–293. [Google Scholar] [PubMed]
- Hatoya, S.; Sugiyama, Y.; Nishida, H.; Okuno, T.; Torii, R.; Sugiura, K.; Kawate, N.; Tamada, H.; Inaba, T. Canine oocyte maturation in culture: Significance of oestrogen and EGF receptor gene expression in cumulus cells. Theriogenology 2009, 71, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Brandon, E.F.A.; Bulder, A.S.; van Engelen, J.G.M.; Mahieu, C.M.; Mennes, W.C.; Pronk, M.E.J.; Rietveld, A.G.; van de Ven, B.M.; ten Voorde, S.E.C.G.; Wolterink, G.; et al. Does EU legislation allow the use of the Benchmark dose (BMD) approach for risk assessment? Regul. Toxicol. Pharm. 2013, 67, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 378–411. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, L.; Fornalski, K.W. Hormesis—Natural phenomenon of answer of organism on stress. In Proceeding of the VII International Scientific Conference: Veterinary Feed Hygiene—The Effects of Mycotoxins on Gastrointestinal Function, Olsztyn, Poland, 23–24 September 2011; Gajęcki, M., Ed.; PTNW: Olsztyn, Poland, 2011; pp. 6–14. [Google Scholar]
- Gajęcka, M.; Stopa, E.; Tarasiuk, M.; Zielonka, Ł.; Gajęcki, M. The expression of type-1 and type-2 nitric oxide synthase in selected tissues of the gastrointestinal tract during mixed mycotoxicosis. Toxins 2013, 5, 2281–2292. [Google Scholar] [CrossRef] [PubMed]
- EFSA. 17th Scientific Colloquium Summary Report; Low-Dose Response in Toxicology and Risk Assessment; EFSA: Parma, Italy, 14–15 June 2012. [Google Scholar]
- Hickey, G.L.; Craig, P.S.; Luttik, R.; de Zwart, D. On the quantification of intertest variability in ecotoxicity data with application to species sensitivity distributions. Environ. Toxicol. Chem. 2012, 31, 1903–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizzell, C.; Verhaegen, S.; Ropstad, E.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of ochratoxin A at the level of nuclear receptor activation and steroidogenesis. Toxicol. Lett. 2013, 217, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Brera, C.; Bertazzoni, V.; Debegnach, F.; Gregori, E.; Prantera, E.; de Santis, B. Exposure assessment for italian population groups to deoxynivalenol deriving from pasta consumption. Toxins 2013, 5, 2293–2309. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dąbrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the intestinal tissues of immature gilts exposed per os to mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, J. Toxicity data relevant for hazard characterization. Toxicol. Lett. 2004, 153, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Muri, S.D.; van der Voet, H.; Boon, P.E.; van Klaveren, J.D.; Brüschweiler, B.J. Comparison of human health risks resulting from exposure to fungicides and mycotoxins via food. Food Chem. Toxicol. 2009, 47, 2963–2974. [Google Scholar] [CrossRef] [PubMed]
- Laufersweilera, M.C.; Gadagbui, B.; Baskerville-Abraham, I.M.; Maier, A.; Willis, A.; Scialli, A.R.; Carr, G.J.; Felter, S.P.; Blackburn, K.; Daston, G. Correlation of chemical structure with reproductive and developmental toxicity as it relates to the use of the threshold of toxicological concern. Regul. Toxicol. Pharm. 2012, 62, 160–182. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Evaluation of the increase of risk for public health related to a possible temporary derogation from the maximum level of deoxynivalenol, zearalenone and fumonisins for maize and maize products. EFSA J. 2014, 12, 1–61. [Google Scholar]
- Barton, M. Position paper: The membrane oestrogen receptor GPER—Clues and questions. Steroids 2012, 77, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Jilani, K.; Lang, F. Ca(2+)-dependent suicidal erythrocyte death following zearalenone exposure. Arch. Toxicol. 2013, 87, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Y.; Zheng, Z.H.; Syn, Y.O.; Shi, X.; Jang, Y.O.; Lee, J.C. Mycotoxin zearalenone induces AIF- and ROS-mediated cell death through p53- and MAPK-dependent signaling pathways in RAW264.7 macrophages. Toxicol. In Vitro 2011, 25, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Przybylska-Gornowicz, B. The low doses effect of experimental zearalenone (ZEN) intoxication on the presence of Ca2+ in selected ovarian cells from pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Hénaff, M.; Quignard, J.F.; Biendon, N.; Morel, J.L.; Macrez, N. T-type calcium channels involved in collagen fragment-induced smooth muscle cell death. Calcium Signal. 2014, 1, 15–23. [Google Scholar]
- Lupescu, A.; Bissinger, R.; Jilani, K.; Lang, F. In vitro induction of erythrocyte phosphatidyloserine translocation by the natural Naphthoquinone Shikonin. Toxins 2014, 6, 1559–1574. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Uhlig, S.; Miles, C.O.; Verhaegen, S.; Elliott, C.T.; Eriksen, G.S.; Sørlie, M.; Ropstad, E.; Connolly, L. Biotransformation of zearalenone and zearalenols to their major glucuronide metabolites reduces estrogenic activity. Toxicol. In Vitro 2015, 29, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Pizzo, F.; Spicer, L.J.; Caloni, F. Fusarium mycotoxins: Effects on reproductive function in domestic animals—A review. Theriogenology 2013, 80, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Dąbrowski, M.; Mróz, M.; Gajęcki, M. The effect of low doses of zearalenone and its metabolites on progesterone and 17β-estradiol concentrations in blood of pre-pubertal female Beagle dogs. Toxicon 2013, 76, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Woźny, M.; Brzuzan, P.; Zielonka, Ł.; Gajęcki, M. Expression of CYPscc and 3β-HSD mRNA in bitches ovary after long-term exposure to zearalenone. Bull. Vet. Inst. Pulawy 2011, 55, 777–780. [Google Scholar]
- Van Cruchten, S.; van den Broeck, W.; D’haeseleer, M.; Simoens, P. Proliferation patterns in the canine endometrium during the estrous cycle. Theriogenology 2004, 62, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M. The effects of experimental administration of low doses of zearalenone on the histology of ovaries in pre pubertal bitches. Pol. J. Vet. Sci. 2013, 16, 313–322. [Google Scholar] [PubMed]
- Rajaram, R.D.; Brisken, C. Paracrine signaling by progesterone. Mol. Cell Endocrinol. 2012, 357, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, J.; Mevissen, M. Inhibition of in vitro metabolism of testosterone in human, dog and horse liver microsomes to investigate species differences. Toxicol. In Vitro 2015, 29, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Labrie, C.; Bélanger, A.; Labrie, F. Androgenic activity of dehydroepiandrosterone and androstenedione in the rat ventral prostate. Endocrinology 1988, 123, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Gajęcki, M. The biotransformation of chosen mycotoxins. Pol. J. Vet. Sci. 2009, 12, 293–303. [Google Scholar] [PubMed]
- Huhtinen, K.; Stahle, M.; Perheentupa, A.; Poutanen, M. Estrogen biosynthesis and signalling in endometriosis. Mol. Cell Endocrinol. 2012, 358, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Gajęcki, M.; Gajęcka, M.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K. Zearalenone–Undesirable Substance. In Mycotoxins in Food, Feed and Bioweapons; Rai, M., Varma, A., Eds.; Springer-Verlag: Heidelberg, Berlin, Germany, 2010; pp. 131–144. [Google Scholar]
- Gajęcka, M. The effect of low-dose experimental zearalenone intoxication on the immunoexpression of oestrogen receptors in the ovaries of pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 685–691. [Google Scholar] [PubMed]
- Couse, J.F.; Lindzey, J.; Gandien, K.; Gustafsson, J.A.; Korach, K.S. Tissue distribution and quantitative analysis of oestrogen receptor-α (ERα) and oestrogen receptor-β (ERβ) messenger ribonucleic acid in the wild-type and ERα-knockout mouse. Endocrinology 1997, 138, 4613–4621. [Google Scholar] [PubMed]
- Liu, Y.; Gao, H.; Marstrand, T.T.; Strom, A.; Valen, E.; Sandelin, A.; Gustafsson, J.-A.; Dahlman-Wright, K. The genome landscape of ERα- and ERβ-binding DNA regions. Proc. Natl. Acad. Sci. USA. 2008, 105, 2604–2609. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Martin-Hirsch, P.L.; Martin, F.L. Oestrogen receptor splice variants in the pathogenesis of disease. Cancer Lett. 2010, 288, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Faria, T.; de Bittencourt Brasil, F.; Sampaio, F.J.B.; da Fonte Ramos, C. Maternal malnutrition during lactation alters the folliculogenesis and gonadotropins and oestrogen isoforms ovarian receptors in the offspring at puberty. J. Endocrinol. 2008, 198, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Schlafer, D.H.; Gifford, A.T. Cystic endometrial hyperplasia, pseudo-placentational endometrial hyperplasia, and other cystic conditions of the canine and feline uterus. Theriogenology 2008, 70, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Bishop, C.V.; Stormshak, F. Non-genomic actions of progesterone and oestrogens in regulating reproductive events in domestic animals. Vet. J. 2008, 176, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on oestrogen receptor a (ERα) and ERβ in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.L.; Russo, M.; England, G.C.W. Uterine artery blood flow characteristics assessed during oestrus and the early luteal phase of pregnant and non-pregnant bitches. Vet. J. 2013, 197, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Scotti, L.; Irusta, G.; Abramovich, D.; Tesone, M.; Parborell, F. Administration of a gonadotropin-releasing hormone agonist affects corpus luteum vascular stability and development and induces luteal apoptosis in a rat model of ovarian hyperstimulation syndrome. Mol. Cell. Endocrinol. 2011, 335, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Fletcher, N.M.; Ali-Fehmi, R.; Diamond, M.P.; Abu-Soud, H.M.; Munkarah, A.R.; Saed, G.M. Modulation of redox signaling promotes apoptosis in epithelial ovarian cancer cells. Gynecol. Oncol. 2011, 122, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Dworakowska, D. Apoptotic index and selected cell cycle regulators in non-small cell lung cancer. Ann. Acad. Med. Gedanensis 2005, 35, 1–112. [Google Scholar]
- Gajęcka, M. The effect of experimental low zearalenone intoxication on ovarian follicles in pre-pubertal bitches. Pol. J. Vet. Sci. 2013, 16, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Meca, G.; Font, G.; Ruiz, M.-J. Interactive effects of zearalenone and its metabolites on cytotoxicity and metabolization in ovarian CHO-K1 cells. Toxicol. In Vitro 2014, 28, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Wang, J.Q.; Zheng, B.Q.; Li, S.L.; Zhang, Y.D.; Li, F.D.; Zheng, N. Cytotoxicity induced by ochratoxin A, zearalenone, α-zearalenol: Effects of individual and combined treatment. Food Chem. Toxicol. 2014, 71, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.I.; Custódio, J.B.A.; Nunes, E.; Oliveira, P.J.; Moreno, A.; Seica, R.; Oliveira, C.R.; Santos, M.S. Mitochondria from distinct tissues are differently affected by 17β-estradiol and tamoxifen. J. Steroid Biochem. 2011, 123, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Y.; Rajamahendran, R. Expression of Bcl-2 and Bax proteins in relation to quality of bovine oocytes and embryos produced in vitro. Anim. Reprod. Sci. 2002, 70, 159–169. [Google Scholar] [CrossRef]
- Songsasen, N.; Fickes, A.; Pukazhenthi, B.S.; Wildt, D.E. Follicular morphology, oocyte diameter and localization of fibroblast growth factors in the domestic dog ovary. Reprod. Domest. Anim. 2009, 44, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Gellerich, F.N.; Gizatullina, Z.; Trumbeckaite, S.; Nguyen, H.P.; Pallas, T.; Arandarcikaite, O.; Vielhaber, S.; Seppet, E.; Striggow, F. The regulation of OXPHOS by extramitochondrial calcium. BBA-Bioenergetics 2010, 1797, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Groppetti, D.; Pecile, A.; Arrighi, S.; di Giancamillo, A.; Cremonesi, F. Endometrial cytology and computerized morphometric analysis of epithelial nuclei: A useful tool for reproductive diagnosis in the bitch. Theriogenology 2010, 73, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Cooke, P.S.; Borsdorf, D.C.; Ekman, G.C.; Doty, K.F.; Clark, S.G.; Dziuk, P.J.; Bartol, F.F. Uterine gland development begins postnatally and is accompanied by oestrogen and progesterone receptor expression in the dog. Theriogenology 2012, 78, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Heberer, T.; Lahrssen-Wiederholt, M.; Schafft, H.; Abraham, K.; Pzyrembel, H.; Henning, K.J.; Schauzu, M.; Braeunig, J.; Goetz, M.; Niemann, L.; et al. Zero tolerances in food and animal feed-Are there any scientific alternatives? A European point of view on an international controversy. Toxicol. Lett. 2007, 175, 118–135. [Google Scholar] [CrossRef] [PubMed]
- Auersperg, N.; Wong, A.S.T.; Choi, K.-C.; Kang, S.K. Ovarian surface epithelium: Biology, endocrinology, and pathology. Endocr. Rev. 2001, 22, 255–288. [Google Scholar] [CrossRef] [PubMed]
- Songsasen, N.; Wildt, D.E. Oocyte biology and challenges in developing in vitro maturation systems in the domestic dog. Anim. Reprod. Sci. 2007, 98, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, J.H.-C. Fibroblasts and myofibroblasts in wound healing: Force generation and measurement. J. Tissue Viability 2011, 20, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review. Molecules 2015, 20, 20669-20687. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules201119726
Gajęcka M, Zielonka Ł, Gajęcki M. The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review. Molecules. 2015; 20(11):20669-20687. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules201119726
Chicago/Turabian StyleGajęcka, Magdalena, Łukasz Zielonka, and Maciej Gajęcki. 2015. "The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review" Molecules 20, no. 11: 20669-20687. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules201119726