
Genetic Diversity and Association of EST-SSR and SCoT Markers with Rust Traits in Orchardgrass (Dactylis glomerata L.)


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. EST-SSR and SCoT Analysis
2.2. Markers Efficiency Analysis

{kind=link}
{kind=link}
| Items | EST-SSRs | SCoTs |
|---|---|---|
| Number of primers | 18 | 21 |
| Number of total loci | 164 | 289 |
| Number of average loci per primers | 9.11 | 13.76 |
| Percentage of polymorphic bands | 0.90 | 0.94 |
| Average band informativeness (Ibav) | 0.59 | 0.62 |
| Effective multiplex ratio (EMR) | 8.22 | 12.95 |
| Mark index (MI) | 4.82 | 8.07 |
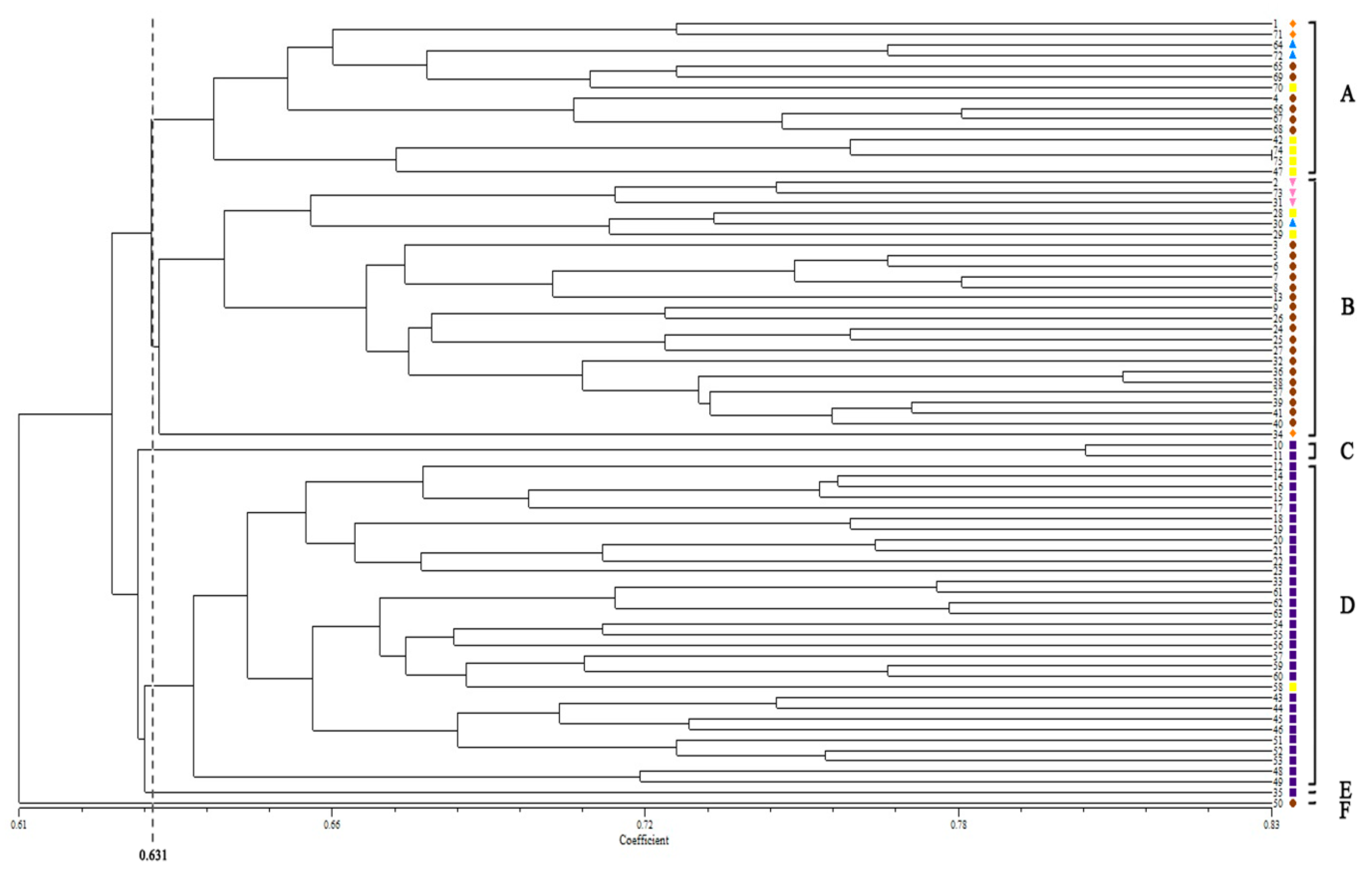
2.3. Cluster Analysis
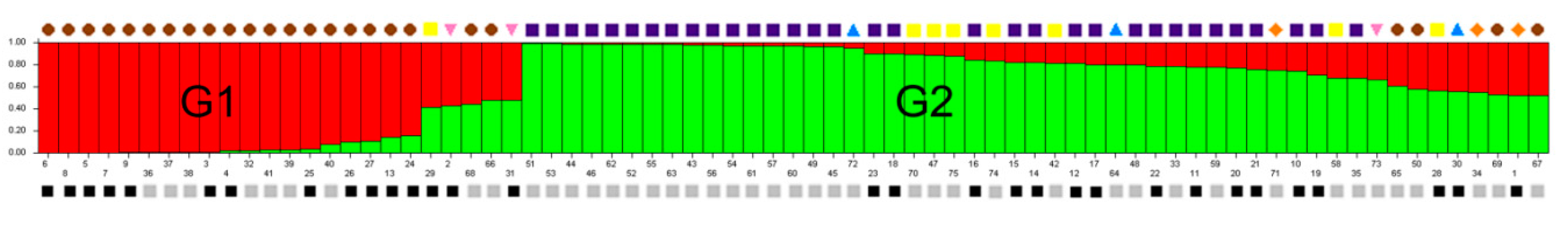
2.4. Population Structure Analysis


2.5. LD Analysis
2.6. Phenotype Analysis
| Continent | Trait | The Number of Accessions |
|---|---|---|
| Asia | resistance | 11 |
| susceptibility | 14 | |
| Europe | resistance | 13 |
| susceptibility | 20 | |
| North America | resistance | 2 |
| susceptibility | 1 | |
| South America | resistance | 1 |
| susceptibility | 2 | |
| Oceania | resistance | 2 |
| susceptibility | 1 | |
| Africa | resistance | 2 |
| susceptibility | 6 |
2.7. Association Analysis
| Location | Year | Alleles | p Value (<0.01) | r2 Value |
|---|---|---|---|---|
| SCo2012-680 bp | 2012 | 156 | 0.0010 | 0.1253 |
| SCoT37-280 bp | 2012 | 321 | 0.0011 | 0.1241 |
| SCo2012-600 bp | 2012 | 157 | 0.0011 | 0.1237 |
| SCo20126-720 bp | 2011 | 268 | 0.0012 | 0.1217 |
| SCo2012-680 bp | 2011 | 156 | 0.0013 | 0.1209 |
| SCoT41-1000 bp | 2012 | 368 | 0.0013 | 0.1205 |
| SCoT44-420 bp | 2011 | 408 | 0.0015 | 0.1179 |
| SCoT37-1000 bp | 2011 | 311 | 0.0015 | 0.1172 |
| EST-SSRH41-42-280 bp | 2012 | 22 | 0.0016 | 0.1168 |
| SCoT44-390 bp | 2011 | 409 | 0.0015 | 0.1167 |
| SCoT37-850 bp | 2012 | 312 | 0.0016 | 0.1162 |
| SCo20110-1000 bp | 2012 | 207 | 0.0017 | 0.1153 |
| SCo20110-580 bp | 2011 | 212 | 0.0017 | 0.115 |
| SCo20116-260 bp | 2011 | 250 | 0.0017 | 0.1144 |
| EST-SSRH41-42-280 bp | 2011 | 22 | 0.0019 | 0.1131 |
| SCo20114-580 bp | 2011 | 225 | 0.0019 | 0.113 |
| SCo20125-1000 bp | 2011 | 251 | 0.0019 | 0.113 |
| SCoT37-350 bp | 2012 | 319 | 0.0019 | 0.1129 |
| SCo20110-340 bp | 2012 | 220 | 0.0021 | 0.1115 |
| EST-SSRH59-60-275 bp | 2012 | 53 | 0.0021 | 0.1114 |
| SCo20126-830 bp | 2011 | 266 | 0.0021 | 0.1107 |
| SCoT37-850 bp | 2011 | 312 | 0.0021 | 0.1107 |
| SCo20110-200 bp | 2011 | 223 | 0.0021 | 0.1106 |
| SCoT42-300 bp | 2012 | 395 | 0.0022 | 0.11 |
| SCoT42-860 bp | 2012 | 383 | 0.0023 | 0.109 |
| SCoT44-750 bp | 2011 | 402 | 0.0023 | 0.1089 |
| SCo20125-850 bp | 2011 | 252 | 0.0023 | 0.1087 |
| SCoT34-250 bp | 2012 | 292 | 0.0025 | 0.1079 |
| EST-SSRH59-60-275 bp | 2011 | 53 | 0.0025 | 0.1067 |
| SCo20127-750 bp | 2011 | 273 | 0.0026 | 0.1063 |
| SCo20110-340 bp | 2011 | 220 | 0.0026 | 0.1062 |
| SCoT8-260 bp | 2012 | 201 | 0.0027 | 0.1056 |
| SCo20116-410 bp | 2011 | 248 | 0.0027 | 0.1052 |
| SCoT8-260 bp | 2011 | 201 | 0.0028 | 0.105 |
| EST-SSRH39-40-175 bp | 2012 | 16 | 0.0028 | 0.1049 |
| SCoT36-350 bp | 2012 | 305 | 0.0030 | 0.1035 |
| SCoT44-750 bp | 2012 | 402 | 0.0031 | 0.1034 |
| SCo20110-1000 bp | 2011 | 207 | 0.0031 | 0.1026 |
| EST-SSRH61-62-175 bp | 2011 | 63 | 0.0038 | 0.0985 |
| SCoT42-320 bp | 2012 | 394 | 0.0041 | 0.0977 |
| EST-SSRH47-48-85 bp | 2012 | 34 | 0.0041 | 0.0976 |
| SCoT42-320 bp | 2011 | 394 | 0.0045 | 0.0951 |
| SCoT37-1500 bp | 2012 | 310 | 0.0046 | 0.0951 |
| SCoT40-230 bp | 2011 | 364 | 0.0046 | 0.0947 |
| SCo20126-830 bp | 2012 | 266 | 0.0047 | 0.0946 |
| SCo20125-200 bp | 2012 | 264 | 0.0048 | 0.0944 |
| SCo2012-720 bp | 2011 | 155 | 0.0048 | 0.094 |
| SCoT42-1800 bp | 2011 | 380 | 0.0051 | 0.0929 |
| SCo20127-350 bp | 2012 | 280 | 0.0053 | 0.0923 |
| EST-SSRH77-78-140 bp | 2012 | 119 | 0.0058 | 0.0904 |
| SCo20125-220 bp | 2012 | 263 | 0.0060 | 0.0898 |
| SCoT6-1100 bp | 2012 | 180 | 0.0060 | 0.0897 |
| SCoT39-500 bp | 2011 | 343 | 0.0060 | 0.0896 |
| SCo20126-680 bp | 2011 | 269 | 0.0061 | 0.0892 |
| SCo20125-220 bp | 2011 | 263 | 0.0062 | 0.0888 |
| SCoT6-1100 bp | 2011 | 180 | 0.0063 | 0.0886 |
| SCoT34-450 bp | 2011 | 285 | 0.0064 | 0.0882 |
| SCoT40-250 bp | 2011 | 363 | 0.0065 | 0.0879 |
| SCoT35-500 bp | 2012 | 296 | 0.0067 | 0.0876 |
| SCoT39-100 bp | 2012 | 351 | 0.0069 | 0.087 |
| SCoT34-360 bp | 2011 | 287 | 0.0069 | 0.0867 |
| EST-SSRH77-78-140 bp | 2011 | 119 | 0.0070 | 0.0864 |
| SCo20126-400 bp | 2011 | 271 | 0.0075 | 0.0849 |
| SCo2012-470 bp | 2012 | 159 | 0.0077 | 0.0847 |
| SCoT41-700 bp | 2011 | 371 | 0.0076 | 0.0846 |
| SCoT39-100 bp | 2011 | 351 | 0.0078 | 0.0842 |
| SCo20126-720 bp | 2012 | 268 | 0.0082 | 0.0834 |
| SCoT36-290 bp | 2012 | 307 | 0.0083 | 0.0832 |
| SCoT42-860 bp | 2011 | 383 | 0.0082 | 0.083 |
| SCo2012-470 bp | 2011 | 159 | 0.0083 | 0.0829 |
| SCoT44-390 bp | 2012 | 409 | 0.0085 | 0.0827 |
| SCo20125-850 bp | 2012 | 252 | 0.0085 | 0.0826 |
| SCoT5-300 bp | 2012 | 178 | 0.0086 | 0.0824 |
| EST-SSRH47-48-230 bp | 2012 | 29 | 0.0095 | 0.0805 |
| SCo20126-400 bp | 2012 | 271 | 0.0098 | 0.0798 |
| Location | Year | Alleles | p Value (<0.01) | r2 Value | Mean r2 Value |
|---|---|---|---|---|---|
| SCoT44-390 bp | 2011 | 409 | 0.0015 | 0.1167 | 0.0997 |
| SCoT44-390 bp | 2012 | 409 | 0.0085 | 0.0827 | |
| SCoT44-750 bp | 2011 | 402 | 0.0023 | 0.1089 | 0.1062 |
| SCoT44-750 bp | 2012 | 402 | 0.0031 | 0.1034 | |
| SCoT42-320 bp | 2012 | 394 | 0.0041 | 0.0977 | 0.0964 |
| SCoT42-320 bp | 2011 | 394 | 0.0045 | 0.0951 | |
| SCoT42-860 bp | 2012 | 383 | 0.0023 | 0.1090 | 0.0960 |
| SCoT42-860 bp | 2011 | 383 | 0.0082 | 0.0830 | |
| SCoT39-100 bp | 2012 | 351 | 0.0069 | 0.0870 | 0.0856 |
| SCoT39-100 bp | 2011 | 351 | 0.0078 | 0.0842 | |
| SCoT37-850 bp | 2012 | 312 | 0.0016 | 0.1162 | 0.1135 |
| SCoT37-850 bp | 2011 | 312 | 0.0021 | 0.1107 | |
| SCo20126-400 bp | 2011 | 271 | 0.0075 | 0.0849 | 0.0824 |
| SCo20126-400 bp | 2012 | 271 | 0.0098 | 0.0798 | |
| SCo20126-720 bp | 2011 | 268 | 0.0012 | 0.1217 | 0.1026 |
| SCo20126-720 bp | 2012 | 268 | 0.0082 | 0.0834 | |
| SCo20126-830 bp | 2011 | 266 | 0.0021 | 0.1107 | 0.1027 |
| SCo20126-830 bp | 2012 | 266 | 0.0047 | 0.0946 | |
| SCo20125-220 bp | 2012 | 263 | 0.0060 | 0.0898 | 0.0893 |
| SCo20125-220 bp | 2011 | 263 | 0.0062 | 0.0888 | |
| SCo20125-850 bp | 2011 | 252 | 0.0023 | 0.1087 | 0.0957 |
| SCo20125-850 bp | 2012 | 252 | 0.0085 | 0.0826 | |
| SCo20110-340 bp | 2012 | 220 | 0.0021 | 0.1115 | 0.1089 |
| SCo20110-340 bp | 2011 | 220 | 0.0026 | 0.1062 | |
| SCo20110-1000 bp | 2012 | 207 | 0.0017 | 0.1153 | 0.1090 |
| SCo20110-1000 bp | 2011 | 207 | 0.0031 | 0.1026 | |
| SCoT8-260 bp | 2012 | 201 | 0.0027 | 0.1056 | 0.1053 |
| SCoT8-260 bp | 2011 | 201 | 0.0028 | 0.1050 | |
| SCoT6-1100 bp | 2012 | 180 | 0.0060 | 0.0897 | 0.0892 |
| SCoT6-1100 bp | 2011 | 180 | 0.0063 | 0.0886 | |
| SCo2012-470 bp | 2012 | 159 | 0.0077 | 0.0847 | 0.0838 |
| SCo2012-470 bp | 2011 | 159 | 0.0083 | 0.0829 | |
| SCo2012-680 bp | 2012 | 156 | 0.0010 | 0.1253 | 0.1231 |
| SCo2012-680 bp | 2011 | 156 | 0.0013 | 0.1209 | |
| EST-SSRH77-78-140 bp | 2012 | 119 | 0.0058 | 0.0904 | 0.0884 |
| EST-SSRH77-78-140 bp | 2011 | 119 | 0.0070 | 0.0864 | |
| EST-SSRH59-60-275 bp | 2012 | 53 | 0.0021 | 0.1114 | 0.1091 |
| EST-SSRH59-60-275 bp | 2011 | 53 | 0.0025 | 0.1067 | |
| EST-SSRH41-42-280 bp | 2012 | 22 | 0.0016 | 0.1168 | 0.1150 |
| EST-SSRH41-42-280 bp | 2011 | 22 | 0.0019 | 0.1131 |
3. Discussion
4. Materials and Methods
4.1. Experimental Materials and the Rust Trait
4.2. DNA Extraction and EST-SSR and SCoT Amplification
4.3. Genetic Diversity and Cluster Analysis
4.4. Markers Efficiency Analysis
4.5. Population Genetic Structure Analysis
4.6. Evaluation of Linkage Disequilibrium
4.7. Association Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stewart, A.V.; Ellison, N.W. The Genus Dactylis. In Wealth of Wild Species: Role in Plant Genome Elucidation and Improvement; Springer: New York, NY, USA, 2010; Volume 2. [Google Scholar]
- Casler, M.D.; Fales, S.L.; McElroy, A.R.; Hall, M.H.; Hoffman, L.D.; Leath, K.T. Genetic progress from 40 years of orchardgrass breeding in north america measured under hay management. Crop Sci. 2000, 40, 1019–1025. [Google Scholar] [CrossRef]
- Mitui, K. Handbook of Herbage Crops and Grasslands; Youkendo: Tokyo, Japan, 1981. [Google Scholar]
- Tajimi, A. Tetrasomic inheritance of stem rust resistance on orchardgra. J. Jpn. Grassl. Sci. 1974, 20, 26–30. [Google Scholar]
- Edwards, M.T.; Sleper, D.A.; Loegering, W.Q. Histology of healthy and diseased orchardgrass leaves subjected to digestion in rumen fluid. Crop Sci. 1981, 21, 341–343. [Google Scholar] [CrossRef]
- Ittu, M.; Kellner, E. Studies on the response to black rust of varieties of cocksfoot (Dactylis glomerata L.). Analele Inst. Cercet. Pl. Technol. 1977, 42, 23–29. [Google Scholar]
- Miller, T.L.; Carlson, I.T. Breeding for rust resistance in orchardgrass by phenotypic and phenotypic-genotypic selection. Crop Sci. 1982, 22, 1218–1221. [Google Scholar] [CrossRef]
- Yan, H.D.; Zeng, B.; Zhang, X.Q.; Cheng, L.; Miller, S.; Huang, L.K. Screening orchardgrass (Dactylis glomerata L.) germplasm for high rust resistance and high summer survival rate in a subtropical climate. Grassl. Sci. 2013, 59, 205–210. [Google Scholar] [CrossRef]
- Fufa, H.; Baenziger, P.S.; Beecher, B.S.; Dweikat, I.; Graybosch, R.A.; Eskridge, K.M. Comparison of phenotypic and molecular marker-based classifications of hard red winter wheat cultivars. Euphytica 2005, 145, 133–146. [Google Scholar] [CrossRef]
- Ali, M.L.; Rajewski, J.F.; Baenziger, P.S.; Gill, K.S.; Eskridge, K.M.; Dweikat, I. Assessment of genetic diversity and relationship among a collection of us sweet sorghum germplasm by ssr markers. Mol. Breed. 2008, 21, 497–509. [Google Scholar] [CrossRef]
- Lande, R.; Thompson, R. Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 1990, 124, 743–756. [Google Scholar]
- Ribaut, J.M.; Hoisington, D. Marker-assisted selection: New tools and strategies. Trends Plant Sci. 1998, 3, 236–239. [Google Scholar] [CrossRef]
- Eleuch, L.; Jilal, A.; Grando, S.; Ceccarelli, S.; von Korff Schmising, M.; Tsujimoto, H.; Hajer, A.; Daaloul, A.; Baum, M. Genetic diversity and association analysis for salinity tolerance, heading date and plant height of barley germplasm using simple sequence repeat markers. J. Integr. Plant Biol. 2008, 50, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.X.; Jenkins, J.N.; McCarty, J.C.; Zhong, M.; Swindle, M. Aflp marker associations with agronomic and fiber traits in cotton. Euphytica 2007, 153, 153–163. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, S.M.; Chen, F.D.; Fang, W.M.; Deng, Y.M.; Chang, Q.S.; Liu, P.S. Genetic analysis and associated srap markers for flowering traits of chrysanthemum (Chrysanthemum morifolium). Euphytica 2011, 177, 15–24. [Google Scholar] [CrossRef]
- Jung, G.; Coyne, D.P.; Skroch, P.W.; Nienhuis, J.; Arnaud-Santana, E.; Bokosi, J.; Ariyarathne, H.M.; Steadman, J.R.; Beaver, J.S.; Kaeppler, S.M. Molecular markers associated with plant architecture and resistance to common blight, web blight, and rust in common beans. J. Am. Soc. Hortic. Sci. 1996, 121, 794–803. [Google Scholar]
- Maccaferri, M.; Sanguineti, M.C.; Mantovani, P.; Demontis, A.; Massi, A.; Ammar, K.; Kolmer, J.A.; Czembor, J.H.; Ezrati, S.; Tuberosa, R. Association mapping of leaf rust response in durum wheat. Mol. Breed. 2010, 26, 189–228. [Google Scholar] [CrossRef]
- La Mantia, J.; Klápště, J.; El-Kassaby, Y.A.; Azam, S.; Guy, R.D.; Douglas, C.J.; Mansfield, S.D.; Hamelin, R. Association analysis identifies melampsora × columbiana poplar leaf rust resistance snps. PLoS ONE 2013, 8, e78423. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.Y.; Mackill, D.J. Start codon targeted (scot) polymorphism: A simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Gupta, P.K.; Rustgi, S.; Sharma, S.; Singh, R.; Kumar, N.; Balyan, H.S. Transferable est-ssr markers for the study of polymorphism and genetic diversity in bread wheat. Mol. Genet. Genom. 2003, 270, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Chabane, K.; Hendre, P.S.; Aggarwal, R.K.; Graner, A. Comparative assessment of est-ssr, est-snp and aflp markers for evaluation of genetic diversity and conservation of genetic resources using wild, cultivated and elite barleys. Plant Sci. 2007, 173, 638–649. [Google Scholar] [CrossRef]
- Luo, C.; He, X.H.; Chen, H.; Hu, Y.; Ou, S.J. Genetic relationship and diversity of Mangifera indica L.: Revealed through scot analysis. Genet. Resour. Crop Evol. 2012, 59, 1505–1515. [Google Scholar] [CrossRef]
- Simko, I. Development of est-ssr markers for the study of population structure in lettuce (Lactuca sativa L.). J. Hered. 2009, 100, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.V.; Rangel, P.N.; Brondani, C.; Martins, W.S.; Melo, L.C.; Carneiro, M.S.; Borba, T.C.O.; Brondani, R.P.V. The characterization of a new set of est-derived simple sequence repeat (ssr) markers as a resource for the genetic analysis of Phaseolus vulgaris. BMC Genet. 2011, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Milbourne, D.; Meyer, R.; Bradshaw, J.E.; Baird, E.; Bonar, N.; Provan, J.; Powell, W.; Waugh, R. Comparison of PCR-based marker systems for the analysis of genetic relationships in cultivated potato. Mol. Breed. 1997, 3, 127–136. [Google Scholar] [CrossRef]
- Gorji, A.M.; Poczai, P.; Polgar, Z.; Taller, J. Efficiency of arbitrarily amplified dominant markers (SCOT, ISSR and RAPD) for diagnostic fingerprinting in tetraploid potato. Am. Potato J. Res. 2011, 88, 226–237. [Google Scholar] [CrossRef]
- Alikhani, L.; Rahmani, M.-S.; Shabanian, N.; Badakhshan, H.; Khadivi-Khub, A. Genetic variability and structure of quercus brantii assessed by issr, irap and scot markers. Gene 2014, 552, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; He, X.H.; Chen, H.; Ou, S.J.; Gao, M.P.; Brown, J.S.; Tondo, C.T.; Schnell, R.J. Genetic diversity of mango cultivars estimated using scot and issr markers. Biochem. Syst. Ecol. 2011, 39, 676–684. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Kumaria, S.; Kumar, S.; Tandon, P. Start codon targeted (SCOT) marker reveals genetic diversity of dendrobium nobile lindl., an endangered medicinal orchid species. Gene 2013, 529, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.H.; Zhu, Y.Q.; Tong, Z.H.; Xu, Z.Y.; Jiang, X.F.; Huang, C.H. Analysis of genetic diversity and relationships among genus lycoris based on start codon targeted (SCOT) marker. Biochem. Syst. Ecol. 2014, 57, 221–226. [Google Scholar] [CrossRef]
- Xiong, F.Q.; Zhong, R.C.; Han, Z.Q.; Jiang, J.; He, L.Q.; Zhuang, W.J.; Tang, R.H. Start codon targeted polymorphism for evaluation of functional genetic variation and relationships in cultivated peanut (Arachis hypogaea L.) genotypes. Mol. Biol. Rep. 2011, 38, 3487–3494. [Google Scholar] [CrossRef] [PubMed]
- Flint-Garcia, S.A.; Thornsberry, J.M.; IV, B. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Rustgi, S.; Kulwal, P.L. Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 2005, 57, 461–485. [Google Scholar] [CrossRef] [PubMed]
- Jun, T.H.; Van, K.; Kim, M.Y.; Lee, S.H.; Walker, D.R. Association analysis using ssr markers to find qtl for seed protein content in soybean. Euphytica 2008, 162, 179–175. [Google Scholar] [CrossRef]
- Ostrowski, M.; David, J.; Santoni, S.; Mckhann, H.; Reboud, X.; Le Corre, V.; Camilleri, C.; Brunel, D.; Bouchez, D.; Faure, B. Evidence for a large-scale population structure among accessions of Arabidopsis thaliana: Possible causes and consequences for the distribution of linkage disequilibrium. Mol. Ecol. 2006, 15, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Malysheva-Otto, L.V.; Ganal, M.W.; Röder, M.S. Analysis of molecular diversity, population structure and linkage disequilibrium in a worldwide survey of cultivated barley germplasm (Hordeum vulgare L.). BMC Genet. 2006, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Azmach, G.; Gedil, M.; Menkir, A.; Spillane, C. Marker-trait association analysis of functional gene markers for provitamin a levels across diverse tropical yellow maize inbred lines. BMC Plant Biol. 2013, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.L.; Liu, S.; Hall, M.D.; Brooks, W.S.; Chao, S.; Muehlbauer, G.J.; Baik, B.K.; Steffenson, B.; Griffey, C.A. Marker-trait associations in virginia tech winter barley identified using genome-wide mapping. Theor. Appl. Genet. 2013, 126, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Sukumaran, S.; Barkley, N.A.; Chen, Z.; Chen, C.Y.; Guo, B.; Pittman, R.N.; Stalker, H.T.; Holbrook, C.C.; Pederson, G.A. Population structure and marker–trait association analysis of the us peanut (Arachis hypogaea L.) mini-core collection. Theor. Appl. Genet. 2011, 123, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Muylle, H.; Baert, J.; van Bockstaele, E.; Moerkerke, B.; Goetghebeur, E.; Roldàn-Ruiz, I. Identification of molecular markers linked with crown rust (Puccinia coronata f. Sp. Lolii) resistance in perennial ryegrass (Lolium perenne) using aflp markers and a bulked segregant approach. Euphytica 2005, 143, 135–144. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.C.; Boyle, T.B.J.; Ye, Z.H.; Mao, J.X. Popgene, the User-friendly Shareware for Population Genetic Analysis; Edmonton, Molecular Biology and Biotechnology Centre, University of Alberta: Edmonton, AL, Canada, 1997. [Google Scholar]
- Rohlf, F.J. Ntsys-pc: Numerical Taxonomy and Multivariate Analysis System; State University of New York: Department of Ecology and Evolution, State University of New York, Stony Brook, NY, 1992. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–475. [Google Scholar]
- Zhang, F.; Ge, S. Data analysis in population genetics. I. Analysis of rapd data with amova. Biodivers. Sci. 2002, 10, 438–444. [Google Scholar]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of rflp, rapd, aflp and ssr (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Archak, S.; Gaikwad, A.B.; Gautam, D.; Rao, E.V.V.B.; Swamy, K.R.M.; Karihaloo, J.L. Comparative assessment of DNA fingerprinting techniques (RAPD, ISSR and AFLP) for genetic analysis of cashew (Anacardium occidentale L.) accessions of india. Genome 2003, 46, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. Tassel: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not Available.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Zhang, Y.; Zeng, B.; Yin, G.; Zhang, X.; Ji, Y.; Huang, L.; Jiang, X.; Liu, X.; Peng, Y.; et al. Genetic Diversity and Association of EST-SSR and SCoT Markers with Rust Traits in Orchardgrass (Dactylis glomerata L.). Molecules 2016, 21, 66. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21010066
Yan H, Zhang Y, Zeng B, Yin G, Zhang X, Ji Y, Huang L, Jiang X, Liu X, Peng Y, et al. Genetic Diversity and Association of EST-SSR and SCoT Markers with Rust Traits in Orchardgrass (Dactylis glomerata L.). Molecules. 2016; 21(1):66. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21010066
Chicago/Turabian StyleYan, Haidong, Yu Zhang, Bing Zeng, Guohua Yin, Xinquan Zhang, Yang Ji, Linkai Huang, Xiaomei Jiang, Xinchun Liu, Yan Peng, and et al. 2016. "Genetic Diversity and Association of EST-SSR and SCoT Markers with Rust Traits in Orchardgrass (Dactylis glomerata L.)" Molecules 21, no. 1: 66. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21010066