
Expression and Functional Activity of the Human Bitter Taste Receptor TAS2R38 in Human Placental Tissues and JEG-3 Cells

 ,
,
Abstract
:1. Introduction
2. Results
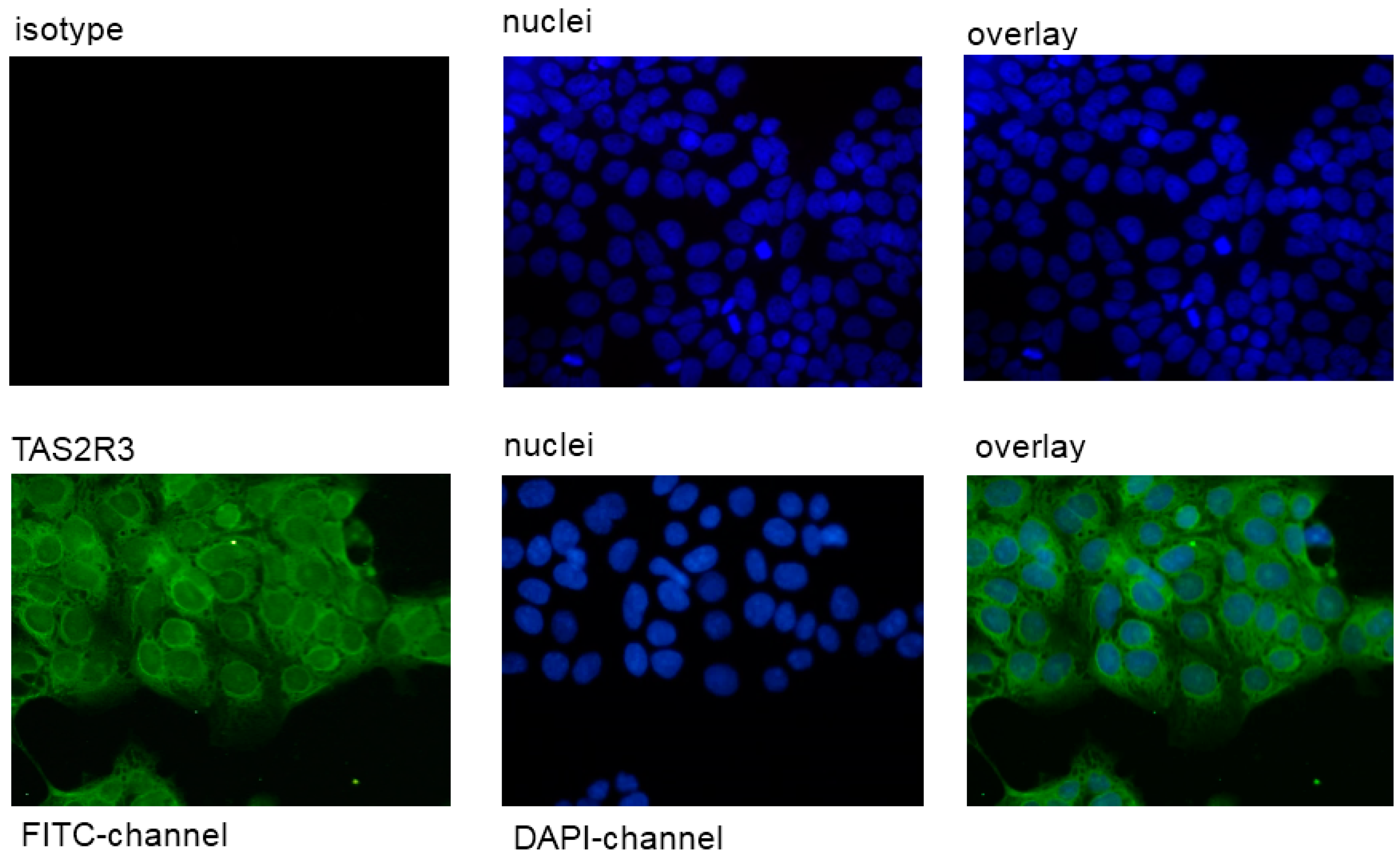
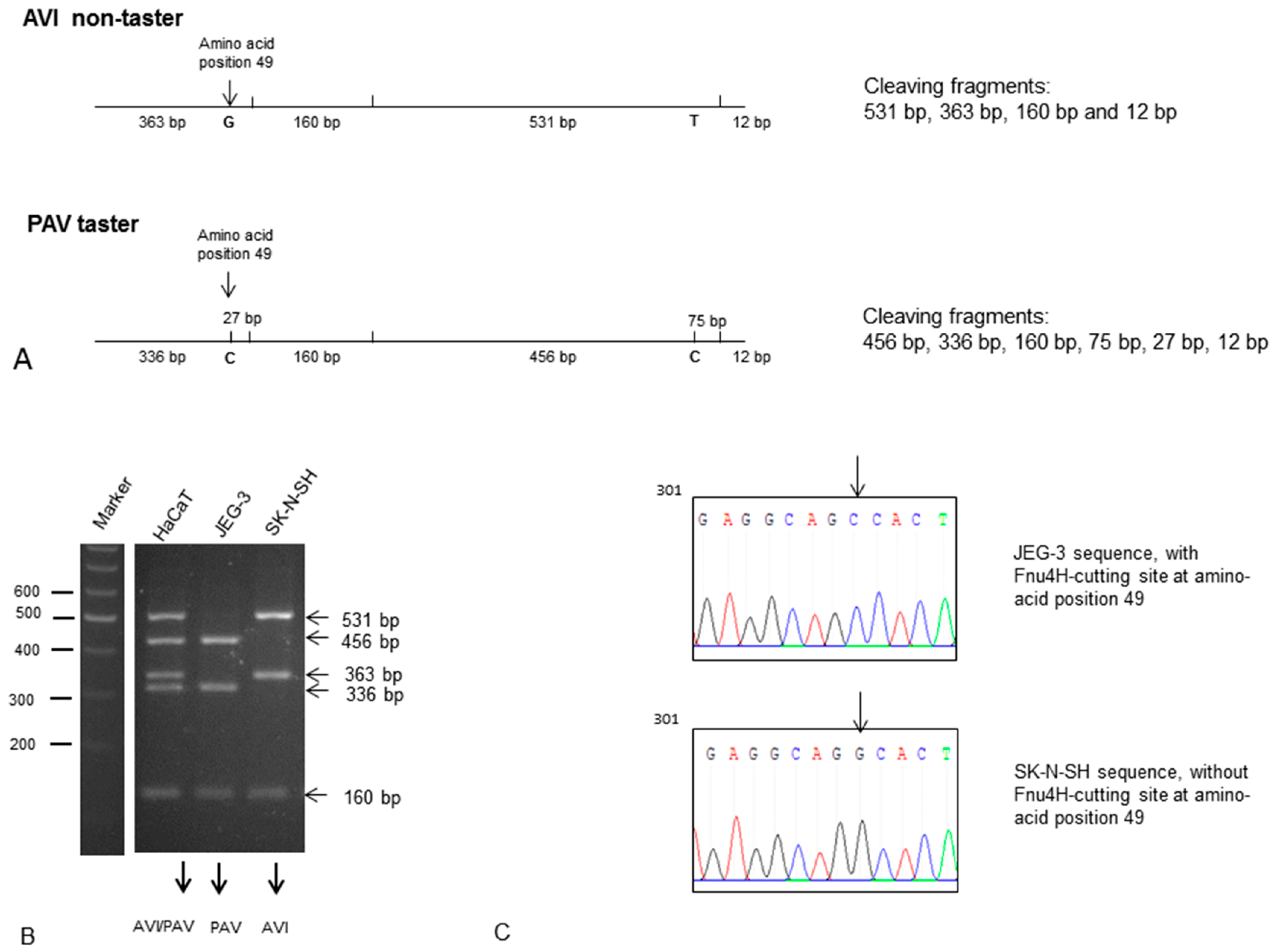
2.1. Expression of TAS2R38 in the Placental Cell Line JEG-3
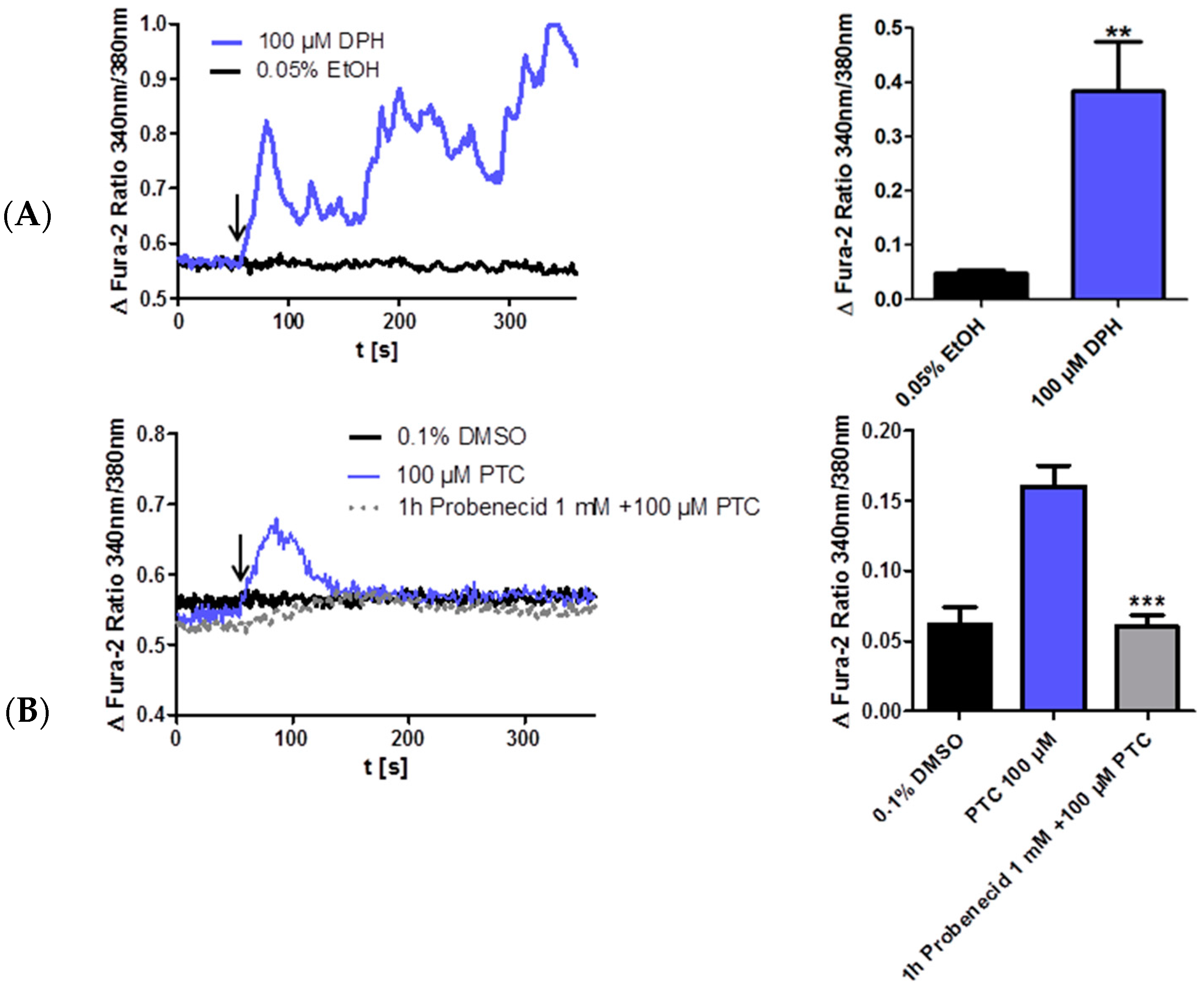
2.2. PTC Induced Calcium Influx in the Taster Cell Line JEG-3
3. Discussion
4. Experimental Section
4.1. Antibodies and Reagents
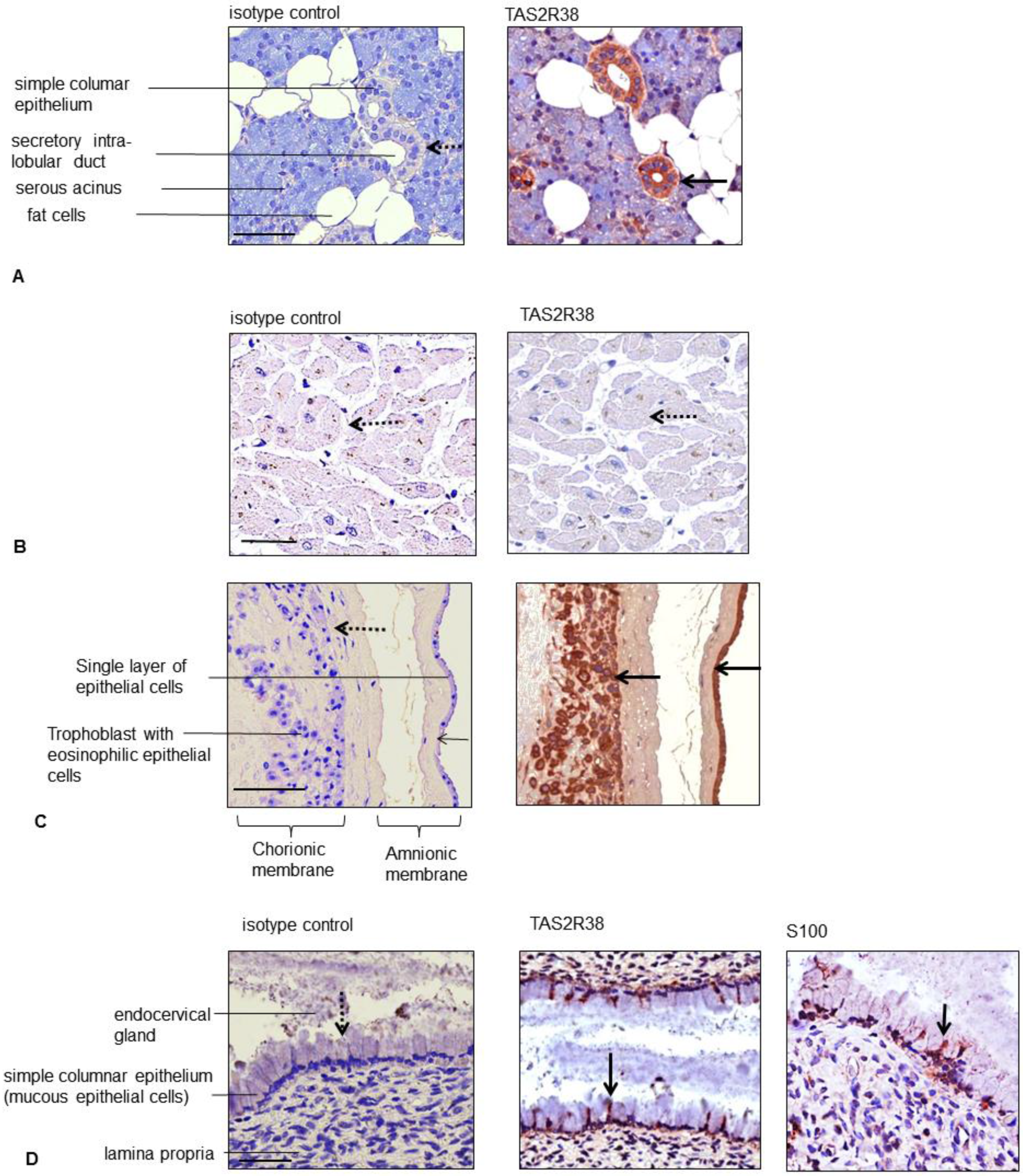
4.2. Immunohistochemistry
4.3. Restriction Enzyme-Based Detection of the TAS2R38 Gene Allele to Distinguish between PTC Taster and Non-Taster
4.4. Immunofluorescence
4.5. Cell Culture
5. Fluorescence Measurements
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yarmolinsky, D.A.; Zuker, C.S.; Ryba, N.J. Common sense about taste: From mammals to insects. Cell 2009, 139, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Adler, E.; Hoon, M.A.; Mueller, K.L.; Chandrashekar, J.; Ryba, N.J.; Zuker, C.S. A novel family of mammalian taste receptors. Cell 2000, 100, 693–702. [Google Scholar] [CrossRef]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; Zuker, C.S.; Ryba, N.J. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef]
- Matsunami, H.; Amrein, H. Taste and pheromone perception in mammals and flies. Genome Biol. 2003, 4, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finger, T.E.; Kinnamon, S.C. Taste Isn’t just for Taste Buds Anymore. F1000 Biol. Rep. 2011, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. Evolutionary biology: The lost appetites. Nature 2012, 486, S16–S17. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.K.; Jorgenson, E.; Coon, H.; Leppert, M.; Risch, N.; Drayna, D. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science 2003, 299, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Gunn, H.C.; Ramos, P.C.; Meyerhof, W.; Wooding, S.P. Genetic, functional, and phenotypic diversity in TAS2R38-mediated bitter taste perception. Chem. Senses 2013, 38, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Biarnes, X.; Marchiori, A.; Giorgetti, A.; Lanzara, C.; Gasparini, P.; Carloni, P.; Born, S.; Brockhoff, A.; Behrens, M.; Meyerhof, W. Insights into the binding of Phenyltiocarbamide (PTC) agonist to its target human TAS2R38 bitter receptor. PLoS ONE 2010, 5, e12394. [Google Scholar] [CrossRef] [PubMed]
- Lipchock, S.V.; Mennella, J.A.; Spielman, A.I.; Reed, D.R. Human bitter perception correlates with bitter receptor messenger RNA expression in taste cells. Am. J. Clin. Nutr. 2013, 98, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.C.; Ranciaro, A.; Froment, A.; Hirbo, J.; Omar, S.; Bodo, J.M.; Nyambo, T.; Lema, G.; Zinshteyn, D.; Drayna, D.; et al. Evolution of functionally diverse alleles associated with PTC bitter taste sensitivity in Africa. Mol. Biol. Evol. 2012, 29, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile cilia of human airway epithelia are chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed]
- Rozengurt, E.; Sternini, C. Taste receptor signaling in the mammalian gut. Curr. Opin. Pharmacol. 2007, 7, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Vrontakis, M.; Parkinson, F.; Chelikani, P. Functional bitter taste receptors are expressed in brain cells. Biochem. Biophys. Res. Commun. 2011, 406, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Elsholz, F.A.; Kersten, A.; Haarhaus, B.; Müller, W.E.; Schempp, C.M. Expression and functional activity of the bitter taste receptors TAS2R1 and TAS2R38 in human keratinocytes. Skin Pharmacol. Physiol. 2015, 28, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Greene, T.A.; Alarcon, S.; Thomas, A.; Berdougo, E.; Doranz, B.J.; Breslin, P.A.; Rucker, J.B. Probenecid inhibits the human bitter taste receptor TAS2R16 and suppresses bitter perception of salicin. PLoS ONE 2011, 6, e20123. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Haarhaus, B.; Kersten, A.; Fiebich, B.; Hug, M.J.; Schempp, C.M. Salicin from willow bark can modulate neurite outgrowth in human neuroblastoma SH-5YSY cells. Phytother. Res. 2015, 29, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Roura, E.; Thomas, W.G. Extrasensory perception: Odorant and taste receptors beyond the nose and mouth. Pharmacol. Ther. 2014, 142, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Born, S.; Redel, U.; Voigt, N.; Schuh, V.; Raguse, J.D.; Meyerhof, W. Immunohistochemical detection of TAS2R38 protein in human taste cells. PLoS ONE 2012, 7, e40304. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.M.; Wong, K.L.; Cheng, K.S.; Kuo, C.S.; Su, T.H.; Chen, Y.W.; Cheng, T.H. Inhibition of voltage-gated K+ channels and Ca2+ channels by diphenidol. Pharmacol. Rep. 2012, 64, 739–744. [Google Scholar] [CrossRef]
- Li, F.; Zhou, M. Depletion of bitter taste transduction leads to massive spermatid loss in transgenic mice. Mol. Hum. Reprod. 2012, 18, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.A.; Liggett, S.B.; Munger, S.D. Extraoral bitter taste receptors as mediators of off-target drug effects. FASEB J. 2012, 26, 4827–4831. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Jones, G.; Zhang, S. Dynamic evolution of bitter taste receptor genes in vertebrates. BMC Evol. Biol. 2009, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.; Wabnitz, G.H.; Kahle, N.A.; Stegmaier, S.; Prior, B.; Giese, T.; Gaida, M.M.; Samstag, Y.; Hansch, G.M. Tasting Pseudomonas aeruginosa Biofilms: Human Neutrophils Express the Bitter Receptor T2R38 as Sensor for the Quorum Sensing Molecule N-(3-Oxododecanoyl)-l-Homoserine Lactone. Front. Immunol. 2015, 6, 369. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Tian, Y.; Fujita, T.; Ikeda, Y.; Kumagai, Y.; Kondo, T.; Tanabe, K.; Nakayama, H.; Horita, S.; Kusuhara, H.; et al. Inhibitory effects of p-aminohippurate and probenecid on the renal clearance of adefovir and benzylpenicillin as probe drugs for organic anion transporter (OAT)1 and OAT3 in humans. Eur. J. Pharm. Sci. 2014, 59, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Witt, M. Carbohydrate histochemistry of vertebrate taste organs. Prog. Histochem. Cytochem. 1996, 30, 1–168. [Google Scholar] [CrossRef]
- Yoshida, R. Hormones and bioactive substances that affect peripheral taste sensitivity. J. Oral Biosci. 2012, 54, 67–72. [Google Scholar] [CrossRef]
- Yoshida, R.; Ohkuri, T.; Jyotaki, M.; Yasuo, T.; Horio, N.; Yasumatsu, K.; Sanematsu, K.; Shigemura, N.; Yamamoto, T.; Margolskee, R.F.; et al. Endocannabinoids selectively enhance sweet taste. Proc. Natl. Acad. Sci. USA 2010, 107, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Dando, R. Endogenous peripheral neuromodulators of the mammalian taste bud. J. Neurophysiol. 2010, 104, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds obtained from Sigma-Aldrich (Taufkirchen, Germany) and are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
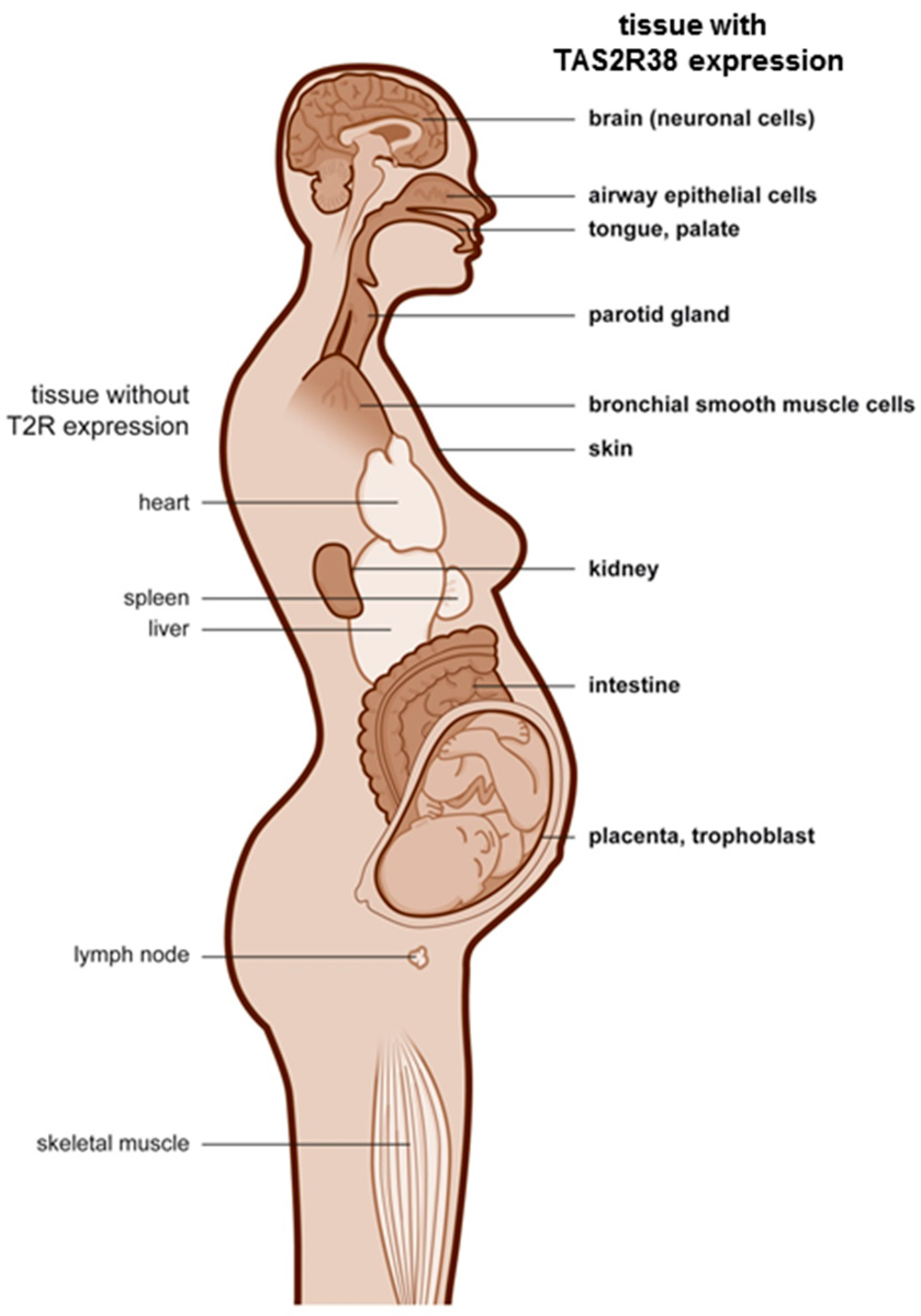
| Blastodermic Layer | Organ System | Cells/Tissues | TAS2R38 Expression |
|---|---|---|---|
| ectodermal | central nervous system | Purkinje cells (Golgi apparatus like staining pattern) in cerebellum | + |
| neurons/glial cells in the spinal cord | - | ||
| ectodermal | skin | keratinocytes, fibroblasts | + |
| mucous membrane (tongue, palate) | + | ||
| ectodermal | placenta | amnion epithelium, | + |
| syncytiotrophoblast, decidua cells | + | ||
| ectodermal | gastrointestinal mucosa and ducts | mucous epithelial cells and gland ductus of: | + |
| ileum, cecum, colon, rectum, parotid gland, kidney, esophagus, stomach, pancreas | + | ||
| breast | - | ||
| endodermal | urogenital system | prostate, exocervix, endocervix | + |
| pro-endometrium (apical) | + (single cells) | ||
| sec-endometrium | - | ||
| myometrium, ovary, liver | - | ||
| endodermal | respiratory ducts | lung | - |
| mesodermal | mesenchymal structures | skeletal muscles, fat tissue, soft tissue, heart | - |
| mesodermal | lymphatic tissue | lymph node, spleen, tonsils, thymus | - |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wölfle, U.; Elsholz, F.A.; Kersten, A.; Haarhaus, B.; Schumacher, U.; Schempp, C.M. Expression and Functional Activity of the Human Bitter Taste Receptor TAS2R38 in Human Placental Tissues and JEG-3 Cells. Molecules 2016, 21, 306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21030306
Wölfle U, Elsholz FA, Kersten A, Haarhaus B, Schumacher U, Schempp CM. Expression and Functional Activity of the Human Bitter Taste Receptor TAS2R38 in Human Placental Tissues and JEG-3 Cells. Molecules. 2016; 21(3):306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21030306
Chicago/Turabian StyleWölfle, Ute, Floriana A. Elsholz, Astrid Kersten, Birgit Haarhaus, Udo Schumacher, and Christoph M. Schempp. 2016. "Expression and Functional Activity of the Human Bitter Taste Receptor TAS2R38 in Human Placental Tissues and JEG-3 Cells" Molecules 21, no. 3: 306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21030306