
Elucidation of Differential Accumulation of 1-Phenylethanol in Flowers and Leaves of Tea (Camellia sinensis) Plants

Abstract
:1. Introduction
2. Results and Discussion
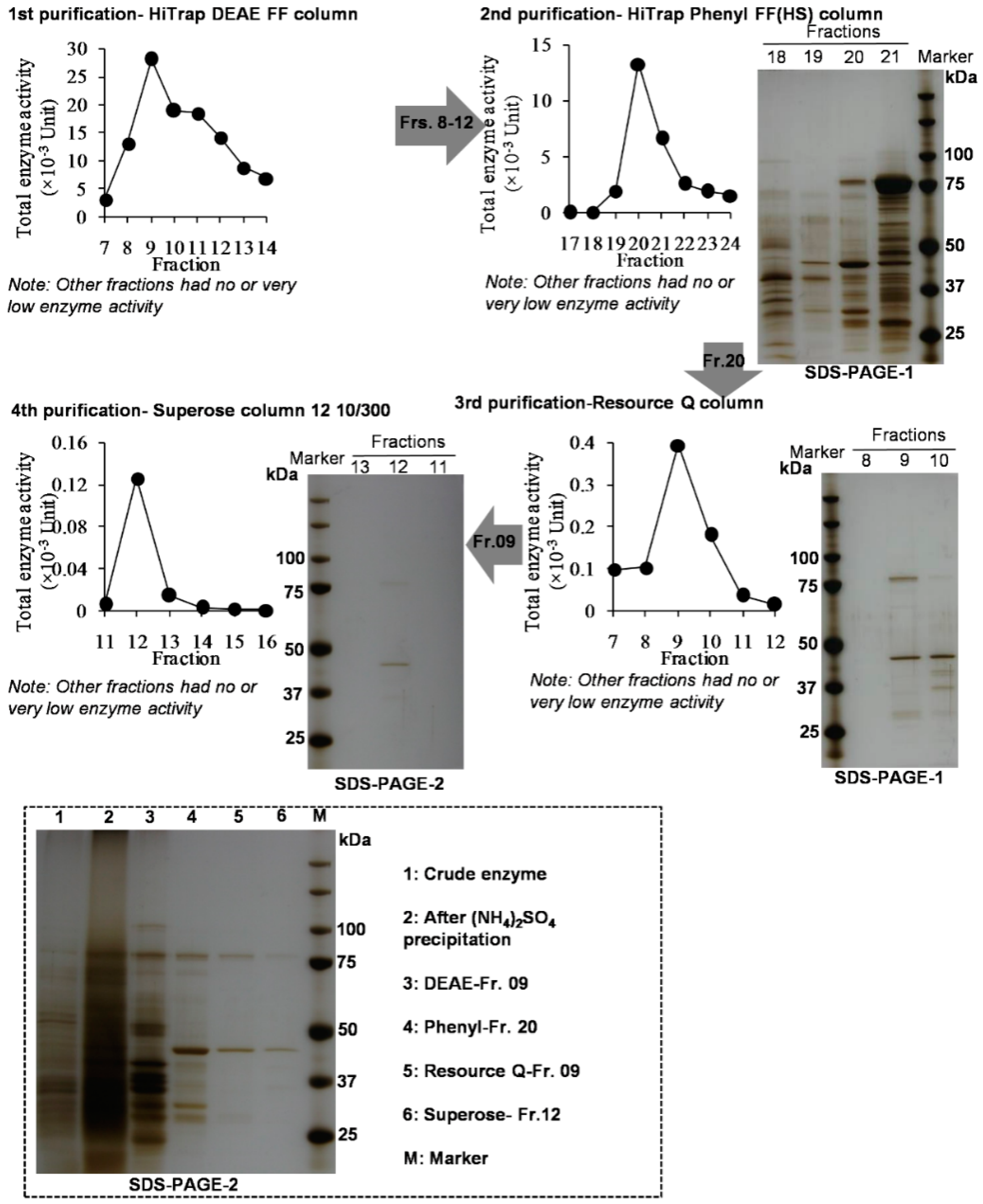
2.1. Tea Flowers Contained Two Types of Enzymes being Responsible for Formations of (R)-1PE and (S)-1PE, Respectively
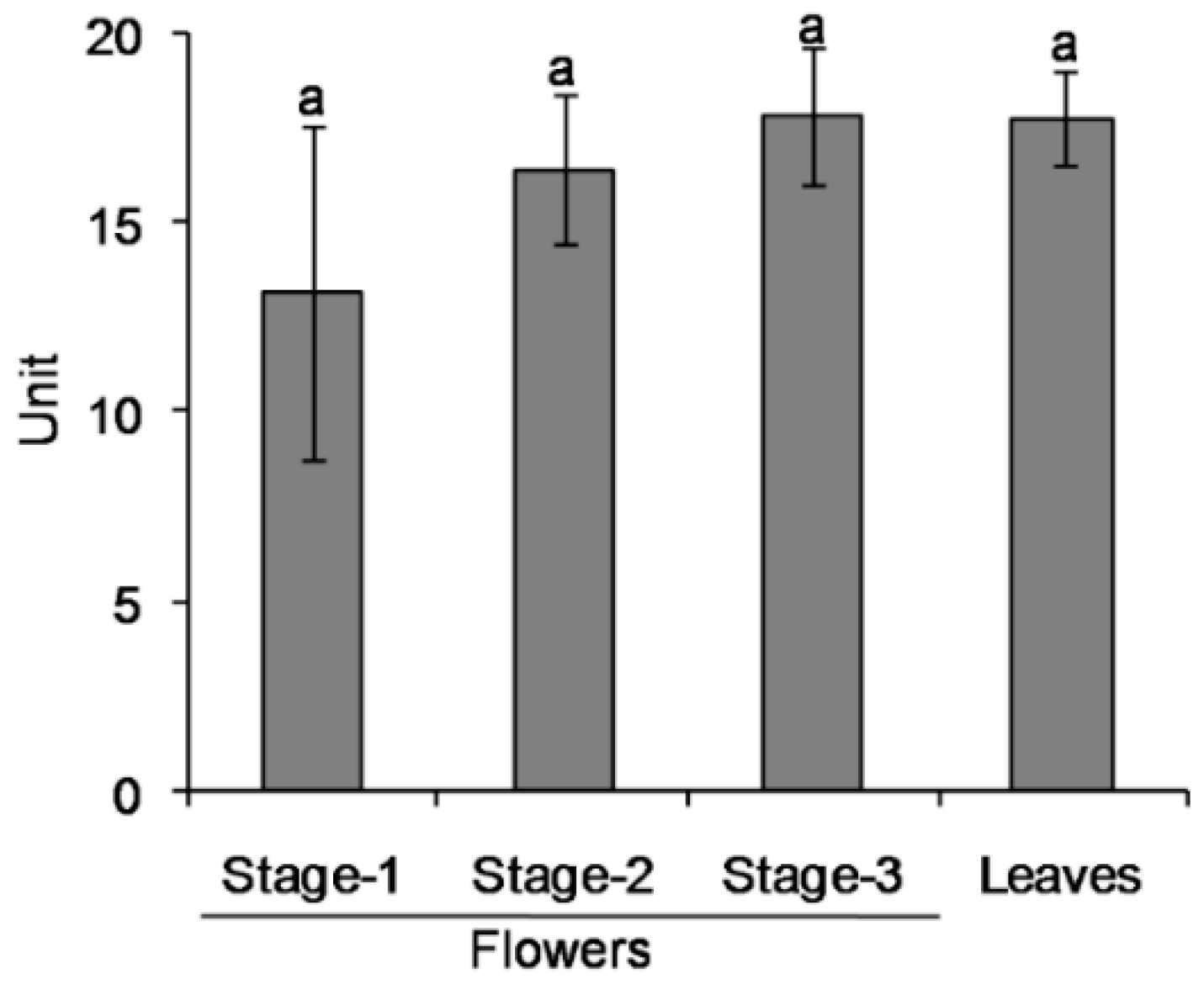
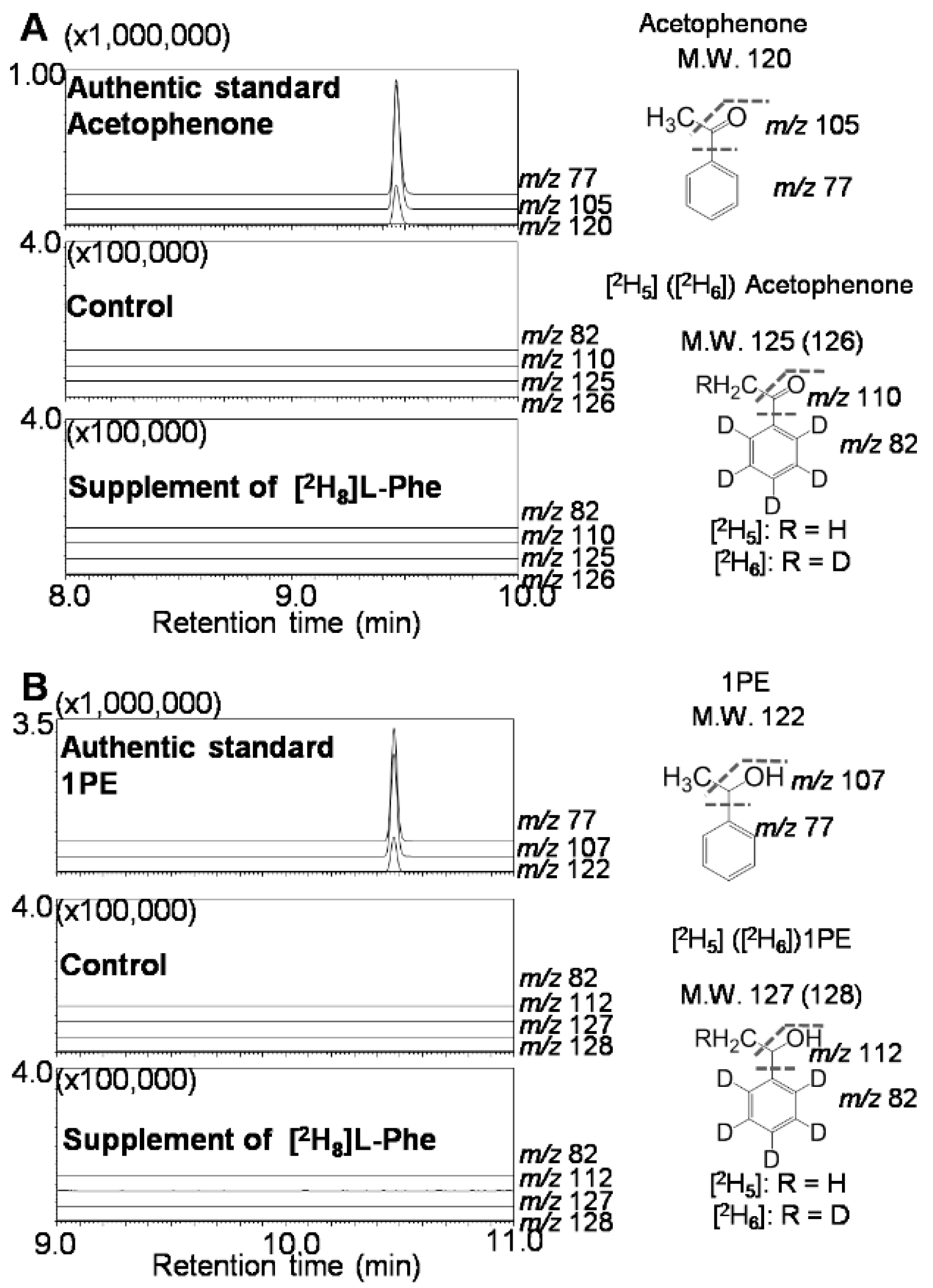
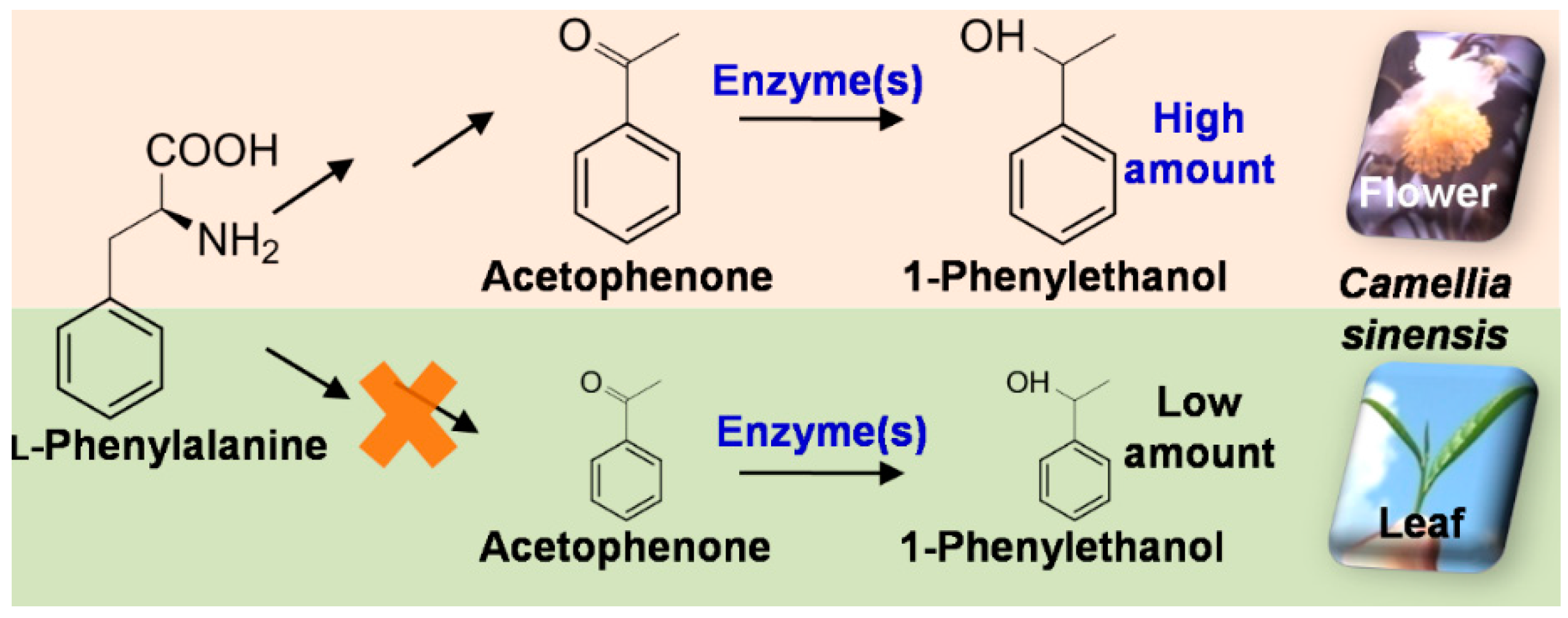
2.2. Differential Distribution of 1PE Between Tea Leaves and Tea Flowers was Due to Availability of Acetophenone
3. Materials and Methods
3.1. Plant Materials
3.2. Determination of Stereochemistry of 1PE from Different Tissues of Tea Flowers
3.3. Extraction of Crude Enzymes Involved in the Pathway Leading from Acetophenone to 1PE from Tea Flowers and Tea Leaves
3.4. Assay of the Crude Enzymes from Tea Flowers and Tea Leaves
3.5. Isolation and Partial Purification of the Enzymes Involved in the Pathway Leading from Acetophenone to 1PE from Tea Flowers
3.6. Supplement of Stable Isotope-Labeled Compound to Tea Leaves and Identification of Products
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CHAPS | 3-[(3-Cholamidopropyl)-dimethylamino]-1-propanesulfonate |
| DTT | dl-Dithiothreitol |
| EDTA | Ethylenediamine tetraacetic acid |
| 1PE | 1-Phenylethanol |
| l-Phe | l-Phenylalanine |
| PVPP | Polyvinylpolypyrrolidone |
| VPBs | Volatile phenylpropanoids and benzenoids |
References
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Negre-Zakharov, F.; Long, M.C.; Dudareva, N. Floral scents and fruit aromas inspired by nature. In Plant-Derived Natural Products, Synthesis, Function, and Application; Osbourn, A.E., Lanzotti, V., Eds.; Springer: Berlin, Germany, 2009; pp. 405–431. [Google Scholar]
- Dong, F.; Fu, X.M.; Watanabe, N.; Su, X.G.; Yang, Z.Y. Recent advances in the emission and functions of plant vegetative volatiles. Molecules 2016, 21, 124. [Google Scholar] [CrossRef] [PubMed]
- Rasch, B.; Büchel, C.; Gais, S.; Born, J. Odor cues during slow-wave sleep prompt declarative memory consolidation. Science 2007, 315, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Pichersky, E. Floral scent metabolic pathways: Their regulation and evolution. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; Taylor & Francis Group: Boca Raton, FL, USA; London, UK, 2006; pp. 55–78. [Google Scholar]
- Cheng, S.; Fu, X.M.; Mei, X.; Zhou, Y.; Du, B.; Watanabe, N.; Yang, Z.Y. Regulation of biosynthesis and emission of volatile phenylpropanoids/benzenoids in petunia× hybrida flowers by multi-factors of circadian clock, light, and temperature. Plant Physiol. Bioch. 2016, 107. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P. The evolution of floral scent and insect chemical communication. Ecol. Lett. 2010, 13, 643–656. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C.; Chen, F.; Pichersky, E. Characterization of an acyltransferase capable of synthesizing benzylbenzoate and other volatile esters in flowers and damaged leaves of Clarkia breweri. Plant Physiol. 2002, 130, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Scalliet, G.; Lionnet, C.; Bechec, M.L.; Dutron, L.; Magnard, J.L.; Baudino, S.; Bergougnoux, V.; Jullien, F.; Chambrier, P.; Vergne, P.; et al. Role of petal-specific orcinol O-methyltransferases in the evolution of rose scent. Plant Physiol. 2006, 140, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Koeduka, T.; Fridman, E.; Gang, D.R.; Vassão, D.G.; Jackson, B.L.; Kish, C.M.; Orlova, I.; Spassova, S.M.; Lewis, N.G.; Noel, J.P.; et al. Eugenol and isoeugenol, characteristic aromatic constituents of spices, are biosynthesized via reduction of a coniferyl alcohol ester. Proc. Natl. Acad. Sci. USA 2006, 103, 10128–10133. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Kobayashi, H.; Sakai, M.; Hirata, H.; Asai, T.; Ohnishi, T.; Baldermann, S.; Watanabe, N. Functional characterization of rose phenylacetaldehyde reductase (PAR), an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol. J. Plant Physiol. 2011, 168, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: nature’s diversity and ingenuity. Science 2006, 331, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Boatright, J.; Negre, F.; Chen, X.L.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004, 135, 1993–2011. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Sakai, M.; Sayama, H.; Shimeno, T.; Yamaguchi, K.; Watanabe, N. Elucidation of the biochemical pathway of 2-phenylethanol from shikimic acid using isolated protoplasts of rose flowers. J. Plant Physiol. 2009, 166, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Yang, Z.Y.; Baldermann, S.; Kajitani, Y.; Ota, S.; Kasuga, H.; Imazeki, Y.; Ohnishi, T.; Watanabe, N. Characterization of l-phenylalanine metabolism to acetophenone and 1-phenylethanol in the flowers of Camellia sinensis using stable isotope labeling. J. Plant Physiol. 2012, 169, 217–225. [Google Scholar] [PubMed]
- Zhou, Y.; Zhang, L.; Gui, J.D.; Dong, F.; Cheng, S.H.; Mei, X.; Zhang, L.Y.; Li, Y.Q.; Su, X.G.; Baldermann, S.; et al. Molecular cloning and characterization of a short chain dehydrogenase showing activity with volatile compounds isolated from Camellia sinensis. Plant Mol. Biol. Rep. 2015, 33, 253–263. [Google Scholar] [CrossRef]
- Höffken, H.W.; Duong, M.; Friedrich, T.; Breuer, M.; Hauer, B.; Reinhardt, R.; Rabus, R.; Heider, J. Crystal structure and enzyme kinetics of the (S)-specific 1-phenylethanol dehydrogenase of the denitrifying bacterium Strain EbN1. Biochemistry 2006, 45, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Nakata, Y.; Cao, C.; Sugiyama, Y.; Asanuma, Y.; Kanamaru, S.; Matsuda, T. Acetophenone reductase with extreme stability against a high concentration of organic compounds or an elevated temperature. Appl. Microbiol. Biotechnol. 2013, 97, 10413–10421. [Google Scholar] [CrossRef] [PubMed]
- Connor, S.O. Methods for molecular identification of biosynthetic enzymes in plants. In Plant-Derived Natural Products, Synthesis, Function, and Application; Osbourn, A.E., Lanzotti, V., Eds.; Springer: Berlin, Germany, 2009; pp. 165–179. [Google Scholar]
- Dudzik, A.; Snoch, W.; Borowiecki, P.; Opalinska-Piskorz, J.; Witko, M.; Heider, J.; Szaleniec, M. Asymmetric reduction of ketones and β-keto esters by (S)-1-phenylethanol dehydrogenase from denitrifying bacterium Aromatoleum aromaticum. Appl. Microbiol. Biotechnol. 2015, 99, 5055–5069. [Google Scholar] [CrossRef] [PubMed]
- Hummel, W. Reduction of acetophenone to R (+)-phenylethanol by a new alcohol dehydrogenase from Lactobacillus kefir. Appl. Microbiol. Biotechnol. 1990, 34, 15–19. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Kobayashi, E.; Katsuno, T.; Asanuma, T.; Fujimori, T.; Ishikawa, T.; Tomomura, M.; Mochizuki, K.; Watase, T.; Nakamura, Y.; et al. Characterisation of volatile and non-volatile metabolites in etiolated leaves of tea (Camellia sinensis) plants in the dark. Food Chem. 2012, 135, 2268–2276. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Baldermann, S.; Watanabe, N. Recent studies of the volatile compounds in tea. Food Res. Int. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Fu, X.M.; Chen, Y.Y.; Mei, X.; Katsuno, T.; Kobayashi, E.; Dong, F.; Watanabe, N.; Yang, Z.Y. Regulation of formation of volatile compounds of tea (Camellia sinensis) leaves by single light wavelength. Sci. Rep. 2015, 5, 16858. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Dong, F.; Kunimasa, A.; Zhang, Y.; Cheng, S.; Lu, J.; Zhang, L.; Murata, A.; Mayer, F.; Fleischmann, P.; et al. Occurrence of glycosidically conjugated 1-phenylethanol and its hydrolase β-primeverosidase in tea (camellia sinensis) flowers. J. Agric. Food Chem. 2014, 62, 8042–8050. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Fu, X.M.; Mei, X.; Zhou, Y.; Du, B.; Tu, Y.Y.; Yang, Z.Y. Characterization of functional proteases from flowers of tea (Camellia sinensis) plants. J. Funct. Foods 2016, 25, 149–159. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds including acetophenone, (R)/(S)-1-phenylethanol, [2H5]ring-acetophenone, and l-[2H8]phenylalanine are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Floral Organs | (R)-1PE | (S)-1PE |
|---|---|---|
| Filaments | 97.2 ± 0.3 a | 2.8 ± 0.3 a |
| Anthers | 92.6 ± 0.1 bc | 7.4 ± 0.1 bc |
| Petals | 90.1 ± 1.8 c | 9.9 ± 1.8 c |
| Receptacles | 86.0 ± 1.9 d | 14.0 ± 1.9 d |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, F.; Zhou, Y.; Zeng, L.; Peng, Q.; Chen, Y.; Zhang, L.; Su, X.; Watanabe, N.; Yang, Z. Elucidation of Differential Accumulation of 1-Phenylethanol in Flowers and Leaves of Tea (Camellia sinensis) Plants. Molecules 2016, 21, 1106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21091106
Dong F, Zhou Y, Zeng L, Peng Q, Chen Y, Zhang L, Su X, Watanabe N, Yang Z. Elucidation of Differential Accumulation of 1-Phenylethanol in Flowers and Leaves of Tea (Camellia sinensis) Plants. Molecules. 2016; 21(9):1106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21091106
Chicago/Turabian StyleDong, Fang, Ying Zhou, Lanting Zeng, Qiyuan Peng, Yiyong Chen, Ling Zhang, Xinguo Su, Naoharu Watanabe, and Ziyin Yang. 2016. "Elucidation of Differential Accumulation of 1-Phenylethanol in Flowers and Leaves of Tea (Camellia sinensis) Plants" Molecules 21, no. 9: 1106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21091106