Saponins from Chinese Medicines as Anticancer Agents

Abstract
:1. Introduction
2. Triterpenoid Saponins
2.1. Ginsenosides
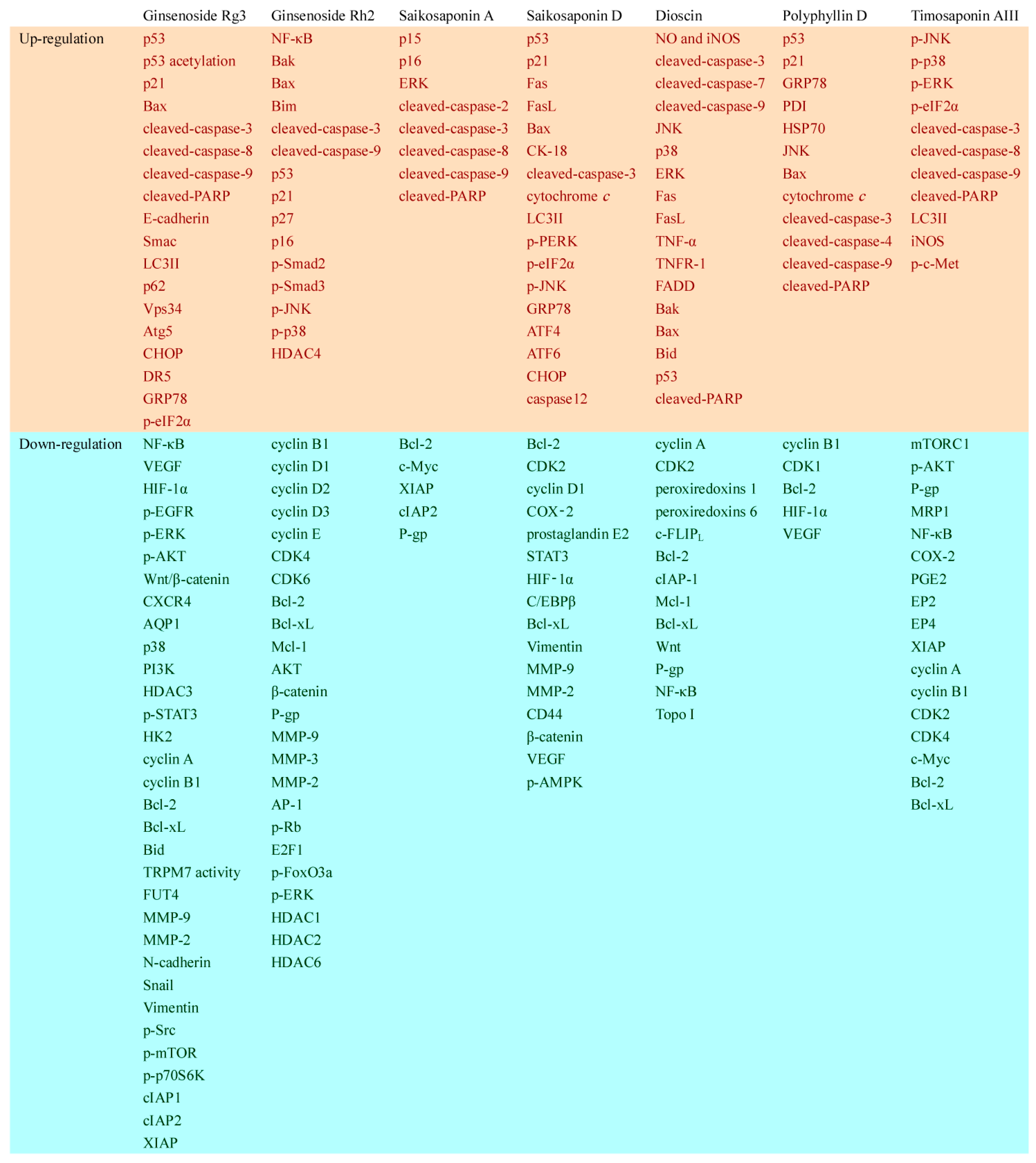
2.1.1. Ginsenoside Rg3
2.1.2. Ginsenoside Rh2
2.1.3. Other Ginsenosides
2.2. Saikosaponins
2.2.1. Saikosaponin A
2.2.2. Saikosaponin D
3. Steroid Saponins
3.1. Dioscin
3.2. Polyphyllin D
3.3. Timosaponin AIII
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lu, J.J.; Dang, Y.Y.; Huang, M.; Xu, W.S.; Chen, X.P.; Wang, Y.T. Anti-cancer properties of terpenoids isolated from Rhizoma Curcumae—A review. J. Ethnopharmacol. 2012, 143, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.J.; Bao, J.L.; Wu, G.S.; Xu, W.S.; Huang, M.Q.; Chen, X.P.; Wang, Y.T. Quinones derived from plant secondary metabolites as anti-cancer agents. Anti-Cancer Agents Med. Chem. 2013, 13, 456–463. [Google Scholar] [CrossRef]
- Lu, J.J.; Bao, J.L.; Chen, X.P.; Huang, M.; Wang, Y.T. Alkaloids isolated from natural herbs as the anticancer agents. Evid.-Based Complement. Altern. Med. 2012, 2012, 485042. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zhao, M.; Tang, L.; Zhang, C.; Song, L.; Wang, W. Ginsenoside Rg1 attenuates hypoxia and hypercapnia-induced vasoconstriction in isolated rat pulmonary arterial rings by reducing the expression of p38. J. Thorac. Dis. 2016, 8, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Yi, X.; Wang, Y.; He, X. Antiproliferative and anti-inflammatory polyhydroxylated spirostanol saponins from Tupistra chinensis. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Shen, X.; Li, S.; Li, Y.; Si, H.; Fan, J.; Li, J.; Gao, E.; Liu, S. Structure-activity relationships of 3-O-β-chacotriosyl oleanane-type triterpenoids as potential H5N1 entry inhibitors. Eur. J. Med. Chem. 2016, 119, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Maryam, A.; Zhang, H.; Mehmood, T.; Ma, T. Killing cancer with platycodin D through multiple mechanisms. J. Cell. Mol. Med. 2016, 20, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.F.; Huang, S.J.; Huang, C.C.; Liu, P.S.; Lin, K.I.; Liu, C.W.; Hsieh, W.C.; Shiu, L.Y.; Chen, C.H. Saikosaponin D induces cell death through caspase-3-dependent, caspase-3-independent and mitochondrial pathways in mammalian hepatic stellate cells. BMC Cancer 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Gao, F.; Sun, X.; Bi, H.; Zhu, Y. Paris saponin vii suppresses osteosarcoma cell migration and invasion by inhibiting MMP-2/9 production via the p38 mapk signaling pathway. Mol. Med. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.W.; Song, F.J.; Li, N.; Dong, X.; Jiang, Y.; Tu, P.F. ASC, a bioactive steroidal saponin from Ophitopogin japonicas, inhibits angiogenesis through interruption of SRC tyrosine kinase-dependent matrix metalloproteinase pathway. Basic Clin. Pharmacol. Toxicol. 2015, 116, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jiang, H.; Li, J.; Xu, J.; Fei, Z. Anticancer effects of paris saponins by apoptosis and PI3k/AKT pathway in gefitinib-resistant non-small cell lung cancer. Med. Sci. Monit. 2016, 22, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.J.; Song, S.C.; Du, L.W.; Zhou, G.H.; Ma, S.L.; Li, J.H.; Feng, J.G.; Zhu, X.H.; Jiang, H. Paris saponins enhance radiosensitivity in a gefitinib-resistant lung adenocarcinoma cell line by inducing apoptosis and G2/M cell cycle phase arrest. Mol. Med. Rep. 2016, 13, 2878–2884. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.F.; Zhang, J.T. New achievements in ginseng research and its future prospects. Chin. J. Integr. Med. 2009, 15, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Anderson, S.; Du, W.; He, T.C.; Yuan, C.S. Red ginseng and cancer treatment. Chin. J. Natl. Med. 2016, 14, 7–16. [Google Scholar]
- Wang, X.Y.; Wang, Y.G.; Wang, Y.F. Ginsenoside Rb1, Rg1 and three extracts of traditional chinese medicine attenuate ultraviolet B-induced G1 growth arrest in hacat cells and dermal fibroblasts involve down-regulating the expression of p16, p21 and p53. Photodermatol. Photoimmunol. Photomed. 2011, 27, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, J.; Sakaguchi, H.; Aoki, I.; Toyoki, H.; Khatun, S.; Tamaya, T. Inhibitory effect of ginsenoside-Rb2 on invasiveness of uterine endometrial cancer cells to the basement membrane. Eur. J. Gynaecol. Oncol. 2001, 22, 339–341. [Google Scholar] [PubMed]
- Lee, Y.J.; Jin, Y.R.; Lim, W.C.; Ji, S.M.; Cho, J.Y.; Ban, J.J.; Lee, S.K. Ginsenoside Rc and Re stimulate c-fos expression in MCF-7 human breast carcinoma cells. Arch. Pharm. Res. 2003, 26, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J. Involvement of melastatin type transient receptor potential 7 channels in ginsenoside Rd-induced apoptosis in gastric and breast cancer cells. J. Ginseng Res. 2013, 37, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Joo, E.J.; Chun, J.; Ha, Y.W.; Lee, J.H.; Han, Y.; Kim, Y.S. Induction of apoptosis by ginsenoside Rk1 in SK-MEL-2-human melanoma. Arch. Pharm. Res. 2012, 35, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Han, I.H.; Kim, Y.J.; Yamabe, N.; Lee, D.; Hwang, G.S.; Oh, M.; Choi, K.C.; Kim, S.N.; Ham, J.; et al. Anticarcinogenic effects of products of heat-processed ginsenoside Re, a major constituent of ginseng berry, on human gastric cancer cells. J. Agric. Food Chem. 2014, 62, 2830–2836. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Siddiqi, M.H.; Aceituno, V.C.; Simu, S.Y.; Yang, D.C. Suppression of MAPKs/NF-κB activation induces intestinal anti-inflammatory action of ginsenoside Rf in Ht-29 and raw264.7 cells. Immunol. Investig. 2016. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.M.; Bai, X.; Jiang, H.F.; He, P.; Wang, J.H. 20-(S)-ginsenoside Rg3 induces apoptotic cell death in human leukemic U937 and HL-60 cells through PI3K/AKT pathways. Anti-Cancer Drugs 2014, 25, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Bae, D.K.; Jeon, J.H.; Lee, J.; Oh, N.; Yang, G.; Yang, Y.H.; Kim, T.K.; Song, J.; Lee, S.H.; et al. Immunopotentiation and antitumor effects of a ginsenoside Rg3-fortified red ginseng preparation in mice bearing H460 lung cancer cells. Environ. Toxicol. Pharmacol. 2011, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Huo, B.; Lv, Y.; Wang, Y.; Liu, W. Ginsenoside Rg3 enhances the inhibitory effects of chemotherapy on esophageal squamous cell carcinoma in mice. Mol. Clin. Oncol. 2014, 2, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Park, E.H.; Kim, Y.J.; Yamabe, N.; Park, S.H.; Kim, H.K.; Jang, H.J.; Kim, J.H.; Cheon, G.J.; Ham, J.; Kang, K.S. Stereospecific anticancer effects of ginsenoside Rg3 epimers isolated from heat-processed american ginseng on human gastric cancer cell. J. Ginseng Res. 2014, 38, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, S.Y.; Yuk, D.Y.; Moon, D.C.; Choi, S.S.; Kim, Y.; Han, S.B.; Oh, K.W.; Hong, J.T. Inhibition of NF-κB by ginsenoside Rg3 enhances the susceptibility of colon cancer cells to docetaxel. Arch. Pharm. Res. 2009, 32, 755–765. [Google Scholar] [CrossRef] [PubMed]
- He, B.C.; Gao, J.L.; Luo, X.; Luo, J.; Shen, J.; Wang, L.; Zhou, Q.; Wang, Y.T.; Luu, H.H.; Haydon, R.C.; et al. Ginsenoside Rg3 inhibits colorectal tumor growth through the down-regulation of WNT/SS-catenin signaling. Int. J. Oncol. 2011, 38, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kim, G.T.; Roh, S.H.; Song, J.S.; Kim, H.J.; Hong, S.S.; Kwon, S.W.; Park, J.H. Proteomic analysis of the anti-cancer effect of 20S-ginsenoside Rg3 in human colon cancer cell lines. Biosci. Biotechnol. Biochem. 2009, 73, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.W.; Chen, X.M.; Chen, X.H.; Zheng, S.S. Ginsenoside Rg3 inhibit hepatocellular carcinoma growth via intrinsic apoptotic pathway. World J. Gastroenterol. 2011, 17, 3605–3613. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.H.; Kim, H.; Hong, M.J.; Yang, M.H.; Kim, J.W.; Yoo, H.; Yang, H.; Park, J.H.; Sung, S.H.; Kim, H.P.; et al. Stereoisomer-specific anticancer activities of ginsenoside Rg3 and RH2 in HepG2 cells: Disparity in cytotoxicity and autophagy-inducing effects due to 20(S)-epimers. Biol. Pharma. Bull. 2015, 38, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.J.; Zhang, M.Z.; Wang, L.X. Gensenoside Rg3 inhibits hypoxia-induced VEGF expression in human cancer cells. Cell. Physiol. Biochem. 2010, 26, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, S.; Ho, J.N.; Byun, S.S.; Hong, S.K.; Lee, S.E.; Lee, E. Synergistic antitumor effect of ginsenoside Rg3 and cisplatin in cisplatinresistant bladder tumor cell line. Oncol. Rep. 2014, 32, 1803–1808. [Google Scholar] [PubMed]
- Kim, B.M.; Kim, D.H.; Park, J.H.; Na, H.K.; Surh, Y.J. Ginsenoside Rg3 induces apoptosis of human breast cancer (MDA-MB-231) cells. J. Cancer Prev. 2013, 18, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhao, L.; Zhang, Y.; Chen, W.; Liu, D.; Hou, H.; Ding, L.; Li, X. Ginsenoside 20(S)-Rg3 targets HIF-1α to block hypoxia-induced epithelial-mesenchymal transition in ovarian cancer cells. PLoS ONE 2014, 9, e103887. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, S.Y.; Cho, J.S.; Son, S.M.; Choi, S.S.; Yun, Y.P.; Yoo, H.S.; Yoon do, Y.; Oh, K.W.; Han, S.B.; et al. Combination of ginsenoside Rg3 with docetaxel enhances the susceptibility of prostate cancer cells via inhibition of NF-κB. Eur. J. Pharmacol. 2010, 631. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, E.H.; Ko, S.R.; Choi, K.J.; Park, J.H.; Im, D.S. Effects of ginsenosides Rg3 and Rh2 on the proliferation of prostate cancer cells. Arch. Pharm. Res. 2004, 27, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Tian, L.L.; Zhang, Y.M.; Wang, X.Q.; Yan, Q.; Liu, J.W. Ginsenoside Rg3 suppresses FUT4 expression through inhibiting NF-κB/p65 signaling pathway to promote melanoma cell death. Int. J. Oncol. 2015, 47, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, H.; Ou-Yang, X.; He, X. Research on the antitumor effect of ginsenoside Rg3 in B16 melanoma cells. Melanoma Res. 2008, 18, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Fu, Y.S.; Aziz, F.; Wang, X.Q.; Yan, Q.; Liu, J.W. Ginsenoside Rg3 inhibits melanoma cell proliferation through down-regulation of histone deacetylase 3 (HDAC3) and increase of p53 acetylation. PLoS ONE 2014, 9, e115401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kang, X.; Zhao, W. Antiangiogenic effect of low-dose cyclophosphamide combined with ginsenoside Rg3 on lewis lung carcinoma. Biochem. Biophys. Res. Commun. 2006, 342, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.M.; Xin, Y.; Cui, M.H.; Jiang, X.; Gu, L.P. Inhibitory effect of ginsenoside Rg3 combined with cyclophosphamide on growth and angiogenesis of ovarian cancer. Chin. Med. J. 2007, 120, 584–588. [Google Scholar] [PubMed]
- Zhang, Q.H.; Wu, C.F.; Duan, L.; Yang, J.Y. Protective effects of ginsenoside Rg3 against cyclophosphamide-induced DNA damage and cell apoptosis in mice. Arch. Toxicol. 2008, 82, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kang, X.; Yang, B.; Wang, J.; Yang, F. Antiangiogenic effect of capecitabine combined with ginsenoside Rg3 on breast cancer in mice. Cancer Biother. Radiopharm. 2008, 23, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.G.; Huang, Y.; Cui, D.D.; Huang, X.B.; Mao, S.H.; Ji, L.L.; Song, H.B.; Yi, C. Inhibitory effect of ginsenoside Rg3 combined with gemcitabine on angiogenesis and growth of lung cancer in mice. BMC Cancer 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.K.; Park, K.K.; Chung, A.S.; Chung, W.Y. Ginsenoside Rg3 enhances the chemosensitivity of tumors to cisplatin by reducing the basal level of nuclear factor erythroid 2-related factor 2-mediated heme oxygenase-1/NAD(P)H quinone oxidoreductase-1 and prevents normal tissue damage by scavenging cisplatin-induced intracellular reactive oxygen species. Food Chem. Toxicol. 2012, 50, 2565–2574. [Google Scholar] [PubMed]
- Kim, D.G.; Jung, K.H.; Lee, D.G.; Yoon, J.H.; Choi, K.S.; Kwon, S.W.; Shen, H.M.; Morgan, M.J.; Hong, S.S.; Kim, Y.S. 20(S)-ginsenoside Rg3 is a novel inhibitor of autophagy and sensitizes hepatocellular carcinoma to doxorubicin. Oncotarget 2014, 5, 4438–4451. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Seong, S.; Kim, S.Y. Synergistic effect of ginsenoside Rg3 with verapamil on the modulation of multidrug resistance in human acute myeloid leukemia cells. Oncol. Lett. 2014, 7, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jung, K.H.; Morgan, M.J.; Kang, Y.R.; Lee, H.S.; Koo, G.B.; Hong, S.S.; Kwon, S.W.; Kim, Y.S. Sensitization of trail-induced cell death by 20(S)-ginsenoside Rg3 via chop-mediated DR5 upregulation in human hepatocellular carcinoma cells. Mol. Cancer Ther. 2013, 12, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Song, Y.M.; Wang, B.; Zhang, F.R.; Yang, R.; Wang, H.Q.; Zhang, G.J. Ginsenoside Rg3 sensitizes human non-small cell lung cancer cells to γ-radiation by targeting the nuclear factor-kappab pathway. Mol. Med. Rep. 2015, 12, 609–614. [Google Scholar] [PubMed]
- Zeng, D.; Wang, J.; Kong, P.; Chang, C.; Li, J.; Li, J. Ginsenoside Rg3 inhibits HIF-1α and vegf expression in patient with acute leukemia via inhibiting the activation of PI3K/AKT and ERK1/2 pathways. Int. J. Clin. Exp. Pathol. 2014, 7, 2172–2178. [Google Scholar] [PubMed]
- Wu, R.; Ru, Q.; Chen, L.; Ma, B.; Li, C. Stereospecificity of ginsenoside Rg3 in the promotion of cellular immunity in hepatoma H22-bearing mice. J. Food Sci. 2014, 79, H1430–H1435. [Google Scholar] [CrossRef] [PubMed]
- Joo, E.J.; Chun, J.; Ha, Y.W.; Ko, H.J.; Xu, M.Y.; Kim, Y.S. Novel roles of ginsenoside Rg3 in apoptosis through downregulation of epidermal growth factor receptor. Chem. Biol. Interact. 2015, 233, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Aziz, F.; Tian, L.L.; Wang, X.Q.; Yan, Q.; Liu, J.W. Ginsenoside Rg3-induced EGFR/MAPK pathway deactivation inhibits melanoma cell proliferation by decreasing FUT4/ley expression. Int. J. Oncol. 2015, 46, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Kim, D.H.; Park, J.H.; Surh, Y.J.; Na, H.K. Ginsenoside Rg3 inhibits constitutive activation of NF-κB signaling in human breast cancer (MDA-MB-231) cells: ERK and AKT as potential upstream targets. J. Cancer Prev. 2014, 19, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.P.; Qian, L.L.; Jiang, H.; Chen, J.H. Ginsenoside Rg3 inhibits CXCR4 expression and related migrations in a breast cancer cell line. Int. J. Clin. Oncol. 2011, 16, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.Y.; Guo, H.; Han, J.; Hao, F.; An, Y.; Xu, Y.; Xiaokaiti, Y.; Pan, Y.; Li, X.J. Ginsenoside Rg3 attenuates cell migration via inhibition of aquaporin 1 expression in PC-3M prostate cancer cells. Eur. J. Pharmacol. 2012, 683, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Nao, J.F.; Zhang, M.; He, P. 20(S)-ginsenoside Rg3 promotes apoptosis in human ovarian cancer HO-8910 cells through PI3K/AKT and XIAP pathways. Tumour Biol. 2014, 35, 11985–11994. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Choi, W.I.; Jeon, B.N.; Choi, K.C.; Kim, K.; Kim, T.J.; Ham, J.; Jang, H.J.; Kang, K.S.; Ko, H. Stereospecific effects of ginsenoside 20-Rg3 inhibits TGF-β1-induced epithelial-mesenchymal transition and suppresses lung cancer migration, invasion and anoikis resistance. Toxicology 2014, 322, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, T.; Zhao, L.; Chen, W.; Hou, H.; Ye, Z.; Li, X. Ginsenoside 20(S) Rg3 inhibits the warburg effect through STAT3 pathways in ovarian cancer cells. Int. J. Oncol. 2015, 46, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, P.; Zeng, H.Q.; Lou, S.F.; Wang, D.X. Ginsenoside Rg3 induces apoptosis in human multiple myeloma cells via the activation of Bcl-2-associated x protein. Mol. Med. Rep. 2015, 12, 3557–3562. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Choi, P.; Lee, D.; Kim, T.; Jung, E.B.; Hwang, B.S.; Kang, K.S.; Ham, J. Effect of amino acids on the generation of ginsenoside Rg3 epimers by heat processing and the anticancer activities of epimers in A2780 human ovarian cancer cells. Evid. Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Aziz, F.; Wang, X.; Liu, J.; Yan, Q. Ginsenoside Rg3 induces FUT4-mediated apoptosis in h. Pylori CAGA-treated gastric cancer cells by regulating SP1 and HSF1 expressions. Toxicol. in Vitro 2016, 31, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Nah, S.Y.; Jeon, J.H.; So, I.; Kim, S.J. Transient receptor potential melastatin 7 channels are involved in ginsenoside Rg3-induced apoptosis in gastric cancer cells. Basic Clin. Pharmacol. Toxicol. 2011, 109, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Shen, D.; Li, X.; Shan, X.; Wang, X.; Yan, Q.; Liu, J. Ginsenoside Rg3 inhibits epithelial-mesenchymal transition (EMT) and invasion of lung cancer by down-regulating FUT4. Oncotarget 2016, 7, 1619–1632. [Google Scholar] [PubMed]
- Guo, J.Q.; Zheng, Q.H.; Chen, H.; Chen, L.; Xu, J.B.; Chen, M.Y.; Lu, D.; Wang, Z.H.; Tong, H.F.; Lin, S. Ginsenoside Rg3 inhibition of vasculogenic mimicry in pancreatic cancer through downregulation of vecadherin/EPHA2/MMP9/MMP2 expression. Int. J. Oncol. 2014, 45, 1065–1072. [Google Scholar] [PubMed]
- Xu, T.M.; Cui, M.H.; Xin, Y.; Gu, L.P.; Jiang, X.; Su, M.M.; Wang, D.D.; Wang, W.J. Inhibitory effect of ginsenoside Rg3 on ovarian cancer metastasis. Chin. Med. J. 2008, 121, 1394–1397. [Google Scholar] [PubMed]
- Kim, J.W.; Jung, S.Y.; Kwon, Y.H.; Lee, S.H.; Lee, J.H.; Lee, B.Y.; Kwon, S.M. Ginsenoside Rg3 inhibits endothelial progenitor cell differentiation through attenuation of VEGF-dependent AKT/ENOS signaling. Phytother. Res. 2012, 26, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Jung, S.Y.; Kwon, Y.H.; Lee, J.H.; Lee, Y.M.; Lee, B.Y.; Kwon, S.M. Ginsenoside Rg3 attenuates tumor angiogenesis via inhibiting bioactivities of endothelial progenitor cells. Cancer Biol. Ther. 2012, 13, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Son, K.J.; Choi, K.R.; Lee, S.J.; Lee, H. Immunogenic cell death induced by ginsenoside Rg3: Significance in dendritic cell-based anti-tumor immunotherapy. Immune Netw. 2016, 16, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kitts, D.D.; Popovich, D.G.; Hu, C. Characterizing the mechanism for ginsenoside-induced cytotoxicity in cultured leukemia (THP-1) cells. Can. J. Physiol. Pharmacol. 2007, 85, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Peng, K.; Wang, L.; Wen, B.; Zhou, L.; Luo, T.; Su, M.; Li, J.; Luo, Z. Ginsenoside Rh2 inhibits proliferation and induces apoptosis in human leukemia cells via TNF-α signaling pathway. Acta Biochim. Biophys. Sin. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.C.; Yang, S.M.; Huang, C.Y.; Chen, J.C.; Chang, W.M.; Hsu, S.L. Molecular mechanisms of ginsenoside Rh2-mediated G1 growth arrest and apoptosis in human lung adenocarcinoma A549 cells. Cancer Chemother. Pharmacol. 2005, 55, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhao, J.; Wang, C.Z.; Searle, J.; He, T.C.; Yuan, C.S.; Du, W. Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett. 2011, 301, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Oh, J.Y.; Kim, S.J. Ginsenoside Rh2 induces Bcl-2 family proteins-mediated apoptosis in vitro and in xenografts in vivo models. J. Cell. Biochem. 2011, 112, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Kim, T.W.; Singh, S.V. Ginsenoside Rh2-mediated G1 phase cell cycle arrest in human breast cancer cells is caused by p15 INK4B and p27 KIP1-dependent inhibition of cyclin-dependent kinases. Pharm. Res. 2009, 26, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Sasa, H.; Kita, T.; Hirata, J.; Tode, T.; Nagata, I. Inhibition of human ovarian cancer cell proliferation in vitro by ginsenoside Rh2 and adjuvant effects to cisplatin in vivo. Anti-Cancer Drugs 1991, 2, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Musende, A.G.; Eberding, A.; Wood, C.; Adomat, H.; Fazli, L.; Hurtado-Coll, A.; Jia, W.; Bally, M.B.; Guns, E.T. Pre-clinical evaluation of Rh2 in PC-3 human xenograft model for prostate cancer in vivo: Formulation, pharmacokinetics, biodistribution and efficacy. Cancer Chemother. Pharmacol. 2009, 64, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Eberding, A.; Madera, C.; Fazli, L.; Jia, W.; Goldenberg, L.; Gleave, M.; Guns, E.S. Rh2 synergistically enhances paclitaxel or mitoxantrone in prostate cancer models. J. Urol. 2006, 175, 1926–1931. [Google Scholar] [CrossRef]
- Kim, Y.S.; Jin, S.H. Ginsenoside Rh2 induces apoptosis via activation of caspase-1 and -3 and up-regulation of bax in human neuroblastoma. Arch. Pharm. Res. 2004, 27, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, D.H.; Han, S.J.; Hyun, J.W.; Kim, H.S. Repression of matrix metalloproteinase gene expression by ginsenoside Rh2 in human astroglioma cells. Biochem. Pharmacol. 2007, 74, 1642–1651. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.F.; Wang, B.X.; Tashiro, S.; Li, T.J.; Ma, J.S.; Ikejima, T. Apoptotic effects of ginsenoside Rh2 on human malignant melanoma A375-S2 cells. Acta Pharmacol. Sin. 2002, 23, 315–322. [Google Scholar] [PubMed]
- Park, E.K.; Lee, E.J.; Lee, S.H.; Koo, K.H.; Sung, J.Y.; Hwang, E.H.; Park, J.H.; Kim, C.W.; Jeong, K.C.; Park, B.K.; et al. Induction of apoptosis by the ginsenoside Rh2 by internalization of lipid rafts and caveolae and inactivation of AKT. Br. J. Pharmacol. 2010, 160, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, M.; Li, P.; Wu, Y.; Chang, C.; Qiu, Y.; Cao, L.; Liu, Z.; Jia, C. Ginsenoside Rh2 inhibits cancer stem-like cells in skin squamous cell carcinoma. Cell. Physiol. Biochem. 2015, 36, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, Q.; Liu, K.; Li, G.; Zheng, R. Ginsenoside Rh2 enhances antitumour activity and decreases genotoxic effect of cyclophosphamide. Basic Clin. Pharmacol. Toxicol. 2006, 98, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Musende, A.G.; Eberding, A.; Jia, W.; Ramsay, E.; Bally, M.B.; Guns, E.T. Rh2 or its aglycone appd in combination with docetaxel for treatment of prostate cancer. Prostate 2010, 70, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Li, J.; Xia, J.; Jiang, R.; Zuo, G.W.; Li, X.P.; Chen, Y.; Xiong, W.; Chen, D.L. Ginsenoside 20(S)-Rh2 as potent natural histone deacetylase inhibitors suppressing the growth of human leukemia cells. Chem. Biol. Interact. 2015, 242, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Rong, N.; Liu, L.J.; Xu, X.L.; Liu, J.T.; Jin, F.X.; Wang, C.M. Antitumoral activity of (20R)- and (20S)-ginsenoside Rh2 on transplanted hepatocellular carcinoma in mice. Planta Med. 2016, 82, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hong, B.; Wu, S.; Niu, T. Inhibition of prostatic cancer growth by ginsenoside Rh2. Tumour Biol. 2015, 36, 2377–2381. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gao, Y.; Ma, W.; Guo, W.; Zhou, G.; Cheng, T.; Liu, Y. EGFR signaling-dependent inhibition of glioblastoma growth by ginsenoside Rh2. Tumour Biol. 2014, 35, 5593–5598. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhao, T.; Liu, H.; Zhang, L. Ginsenoside Rh2 inhibits hepatocellular carcinoma through β-catenin and autophagy. Sci. Rep. 2016, 6, 19383. [Google Scholar] [PubMed]
- Zhang, J.; Zhou, F.; Wu, X.; Gu, Y.; Ai, H.; Zheng, Y.; Li, Y.; Zhang, X.; Hao, G.; Sun, J.; et al. 20(S)-ginsenoside Rh2 noncompetitively inhibits p-glycoprotein in vitro and in vivo: A case for herb-drug interactions. Drug Metab. Dispos. 2010, 38, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xiao, X.; Xu, L.; Zhu, L.; Tan, L.; Tang, H.; Zhang, Y.; Xie, Q.; Yao, S. A dynamic study on reversal of multidrug resistance by ginsenoside Rh2 in adriamycin-resistant human breast cancer MCF-7 cells. Talanta 2012, 88, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, F.; Wu, X.; Zhang, X.; Chen, Y.; Zha, B.S.; Niu, F.; Lu, M.; Hao, G.; Sun, Y.; et al. Cellular pharmacokinetic mechanisms of adriamycin resistance and its modulation by 20(S)-ginsenoside Rh2 in MCF-7/ADR cells. Br. J. Pharmacol. 2012, 165, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Li, J.; Feng, Z.; Zhao, L.; Luo, L.; You, Z.; Li, D.; Xia, J.; Zuo, G.; Chen, D. Effect of ginsenoside Rh2 on the migratory ability of HEPG2 liver carcinoma cells: Recruiting histone deacetylase and inhibiting activator protein 1 transcription factors. Mol. Med. Rep. 2014, 10, 1779–1785. [Google Scholar] [PubMed]
- Dong, H.; Bai, L.P.; Wong, V.K.; Zhou, H.; Wang, J.R.; Liu, Y.; Jiang, Z.H.; Liu, L. The in vitro structure-related anti-cancer activity of ginsenosides and their derivatives. Molecules 2011, 16, 10619–10630. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Choi, Y.J.; Lee, S.G. Ginsenoside Rh1 suppresses matrix metalloproteinase-1 expression through inhibition of activator protein-1 and mitogen-activated protein kinase signaling pathway in human hepatocellular carcinoma cells. Eur. J. Pharmacol. 2012, 679, 24–33. [Google Scholar] [PubMed]
- Ha, S.E.; Shin, D.H.; Kim, H.D.; Shim, S.M.; Kim, H.S.; Kim, B.H.; Lee, J.S.; Park, J.K. Effects of ginsenoside Rg2 on the ultraviolet B-induced DNA damage responses in hacat cells. Naunyn Schmiedebergs Arch. Pharmacol. 2010, 382, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Yu, X.; Guo, D.; Yu, B.; Li, L.; Liao, Q.; Xing, R. Ginsenoside Rg1 attenuates invasion and migration by inhibiting transforming growth factor-β-induced epithelial to mesenchymal transition in HepG2 cells. Mol. Med. Rep. 2015, 11, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Qi, B.; Yuan, D.; Dong, S.; Guo, D.; Zhang, C.; Yu, M. Suppression of PMA-induced tumor cell invasion and migration by ginsenoside Rg1 via the inhibition of NF-κB-dependent MMP-9 expression. Oncol. Rep. 2014, 32, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wei, Q.; Zuo, G.W.; Xia, J.; You, Z.M.; Li, C.L.; Chen, D.L. Ginsenoside Rg1 induces apoptosis through inhibition of the EPOR-mediated JAK2/STAT5 signalling pathway in the TF-1/ EPO human leukemia cell line. Asian Pac. J. Cancer Prev. 2014, 15, 2453–2459. [Google Scholar] [CrossRef] [PubMed]
- Bermejo Benito, P.; Abad Martinez, M.J.; Silvan Sen, A.M.; Sanz Gomez, A.; Fernandez Matellano, L.; Sanchez Contreras, S.; Diaz Lanza, A.M. In vivo and in vitro antiinflammatory activity of saikosaponins. Life Sci. 1998, 63, 1147–1156. [Google Scholar] [CrossRef]
- Chiang, L.C.; Ng, L.T.; Liu, L.T.; Shieh, D.E.; Lin, C.C. Cytotoxicity and anti-hepatitis B virus activities of saikosaponins from Bupleurum species. Planta Medica 2003, 69, 705–709. [Google Scholar] [PubMed]
- Cheng, P.W.; Ng, L.T.; Chiang, L.C.; Lin, C.C. Antiviral effects of saikosaponins on human coronavirus 229E in vitro. Clin. Exp. Pharmacol. Physiol. 2006, 33, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Kong, J.; Wang, D.; Lien, L.L.; Lien, E.J. A survey of chinese herbal ingredients with liver protection activities. Chin. Med. 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Sato, N.; Yoshida, M.; Satoh, N.; Ueda, S. Effects of bupleurum scorzoneraefolium, bupleurum falcatum, and saponins on nephrotoxic serum nephritis in mice. J. Ethnopharmacol. 2008, 116, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Yano, H.; Mizoguchi, A.; Fukuda, K.; Haramaki, M.; Ogasawara, S.; Momosaki, S.; Kojiro, M. The herbal medicine SHO-SAIKO-TO inhibits proliferation of cancer cell lines by inducing apoptosis and arrest at the G0/G1 phase. Cancer Res. 1994, 54, 448–454. [Google Scholar] [PubMed]
- Kao, S.T.; Yeh, C.C.; Hsieh, C.C.; Yang, M.D.; Lee, M.R.; Liu, H.S.; Lin, J.G. The chinese medicine Bu-Zhong-Yi-Qi-Tang inhibited proliferation of hepatoma cell lines by inducing apoptosis via G0/G1 arrest. Life Sci. 2001, 69, 1485–1496. [Google Scholar] [CrossRef]
- Tatsuta, M.; Iishi, H.; Baba, M.; Nakaizumi, A.; Uehara, H. Inhibition by Xiao-Chai-Hu-Tang (TJ-9) of development of hepatic FOCI induced by n-nitrosomorpholine in sprague-dawley rats. Jpn. J. Cancer Res. 1991, 82, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Shyu, K.G.; Tsai, S.C.; Wang, B.W.; Liu, Y.C.; Lee, C.C. Saikosaponin c induces endothelial cells growth, migration and capillary tube formation. Life Sci. 2004, 76, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z.; Fujikawa-Yamamoto, K.; Tanino, M.; Teraoka, K.; Yamagishi, H.; Odashima, S. Saikosaponin B2-induced apoptosis of cultured B16 melanoma cell line through down-regulation of PKC activity. Biochem. Biophys. Res. Commun. 1996, 219, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Wu, J.; Liu, L.Y.; Wu, Y.Y.; Li, L.Z.; Huang, X.X.; Liu, Q.B.; Yang, J.Y.; Song, S.J.; Wu, C.F. Cytotoxic triterpenoid glycosides (saikosaponins) from the roots of Bupleurum chinense. Bioorg. Med. Chem. Lett. 2015, 25, 3887–3892. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Xu, M.Y.; Shehzad, O.; Seo, E.K.; Kim, Y.S. Separation of triterpenoid saponins from the root of Bupleurum falcatum by counter current chromatography: The relationship between the partition coefficients and solvent system composition. J. Sep. Sci. 2014, 37, 3587–3594. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.S.; Hsu, H.Y. Involvement of p-15INK4b and p-16INK4a gene expression in saikosaponin a and TPA-induced growth inhibition of HepG2 cells. Biochem. Biophys. Res. Commun. 2001, 285, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Murakami, T.; Kimura, Y.; Takahashi, M.; Okita, K. Saikosaponin a-induced cell death of a human hepatoma cell line (HUH-7): The significance of the sub-G1 peak in a DNA histogram. Pathol. Int. 1995, 45, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Chang, N.W.; Chung, J.G.; Chen, K.C. Saikosaponin—A induces apoptotic mechanism in human breast MDA-MB-231 and MCF-7 cancer cells. Am. J. Chin. Med. 2003, 31, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.J.; Chen, I.L.; Horng, L.Y.; Wu, R.T. Induction of differentiation in rat c6 glioma cells with saikosaponins. Phytother. Res. 2002, 16, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Hong, S.H. Sequential caspase-2 and caspase-8 activation is essential for saikosaponin a-induced apoptosis of human colon carcinoma cell lines. Apoptosis 2011, 16, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.P.; Chen, Z.D. Saikosaponin A, an active glycoside from Radix bupleuri, reverses P-glycoprotein-mediated multidrug resistance in MCF-7/ADR cells and HepG2/adm cells. Xenobiotica 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Kuo, P.L.; Lin, C.C. The proliferative inhibition and apoptotic mechanism of Saikosaponin D in human non-small cell lung cancer A549 cells. Life Sci. 2004, 75, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Motoo, Y.; Sawabu, N. Antitumor effects of saikosaponins, baicalin and baicalein on human hepatoma cell lines. Cancer Lett. 1994, 86, 91–95. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Kuo, P.L.; Chiang, L.C.; Lin, C.C. Involvement of p53, nuclear factor kappab and FAS/FAS ligand in induction of apoptosis and cell cycle arrest by saikosaponin D in human hepatoma cell lines. Cancer Lett. 2004, 213, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Dang, S.; Cheng, Y.; Zhang, X.; Li, M.; Li, Y.; Li, S. Effects of saikosaponin-d on syndecan-2, matrix metalloproteinases and tissue inhibitor of metalloproteinases-2 in rats with hepatocellular carcinoma. J. Tradit. Chin. Med. 2012, 32, 415–422. [Google Scholar] [CrossRef]
- Yao, M.; Yang, J.; Cao, L.; Zhang, L.; Qu, S.; Gao, H. Saikosaponind inhibits proliferation of DU145 human prostate cancer cells by inducing apoptosis and arresting the cell cycle at G0/G1 phase. Mol. Med. Rep. 2014, 10, 365–372. [Google Scholar] [PubMed]
- Liu, R.Y.; Li, J.P. Saikosaponin-d inhibits proliferation of human undifferentiated thyroid carcinoma cells through induction of apoptosis and cell cycle arrest. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2435–2443. [Google Scholar] [PubMed]
- Wang, B.F.; Wang, X.J.; Kang, H.F.; Bai, M.H.; Guan, H.T.; Wang, Z.W.; Zan, Y.; Song, L.Q.; Min, W.L.; Lin, S.; et al. Saikosaponin-d enhances radiosensitivity of hepatoma cells under hypoxic conditions by inhibiting hypoxia-inducible factor-1α. Cell. Physiol. Biochem. 2014, 33, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.L.; He, S.X.; Ren, M.D.; Wang, Y.L.; Zhang, Y.X.; Liu, E.Q. Chemopreventive effect of saikosaponin-d on diethylinitrosamine-induced hepatocarcinogenesis: Involvement of CCAAT/enhancer binding protein beta and cyclooxygenase-2. Mol. Med. Rep. 2012, 5, 637–644. [Google Scholar] [PubMed]
- He, S.; Lu, G.; Hou, H.; Zhao, Z.; Zhu, Z.; Lu, X.; Chen, J.; Wang, Z. Saikosaponind suppresses the expression of cyclooxygenase2 through the phosphosignal transducer and activator of transcription 3/hypoxiainducible factor1alpha pathway in hepatocellular carcinoma cells. Mol. Med. Rep. 2014, 10, 2556–2562. [Google Scholar] [PubMed]
- Wang, B.F.; Lin, S.; Bai, M.H.; Song, L.Q.; Min, W.L.; Wang, M.; Yang, P.; Ma, H.B.; Wang, X.J. Effects of SSD combined with radiation on inhibiting SMMC-7721 hepatoma cell growth. Med. Sci. Monit. 2014, 20, 1340–1344. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.K.; Li, T.; Law, B.Y.; Ma, E.D.; Yip, N.C.; Michelangeli, F.; Law, C.K.; Zhang, M.M.; Lam, K.Y.; Chan, P.L.; et al. Saikosaponin-d, a novel serca inhibitor, induces autophagic cell death in apoptosis-defective cells. Cell Death Dis. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Zhang, H.J.; Jiang, Y.D.; Wu, P.; Qi, H.; Cai, C.; Zheng, S.B.; Dang, Q. Saikosaponin-d: A potential chemotherapeutics in castration resistant prostate cancer by suppressing cancer metastases and cancer stem cell phenotypes. Biochem. Biophys. Res. Commun. 2016, 474, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Xu, Y.; Xu, L.; Cong, X.; Yin, L.; Li, H.; Peng, J. Mechanism investigation of dioscin against CCl4-induced acute liver damage in mice. Environ. Toxicol. Pharmacol. 2012, 34, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Sun, X.; Yin, L.; Han, X.; Xu, L.; Qi, Y.; Xu, Y.; Li, H.; Lin, Y.; Liu, K.; et al. Dioscin ameliorates cerebral ischemia/reperfusion injury through the downregulation of TLR4 signaling via hmgb-1 inhibition. Free Radic. Biol. Med. 2015, 84, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Xu, H.; Peng, J.; Wang, C.; Jin, Y.; Liu, K.; Sun, H.; Qin, J. Potent anti-inflammatory effect of dioscin mediated by suppression of TNF-α-induced vcam-1, icam-1and el expression via the NF-κB pathway. Biochimie 2015, 110, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Wang, Z.; Ju, Y.; Zhou, J.B.; Wang, Y.; Wong, R.N. The mitotic-arresting and apoptosis-inducing effects of diosgenyl saponins on human leukemia cell lines. Biol. Pharm. Bull. 2004, 27, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, J.; Ju, Y.; Zhang, H.; Liu, M.; Li, X. Effects of two saponins extracted from the Polygonatum Zanlanscianense pamp on the human leukemia (HL-60) cells. Biol. Pharm. Bull. 2001, 24, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.J.; Tsai, T.L.; Hsieh, Y.S.; Wang, C.J.; Chiou, H.L. Dioscin-induced autophagy mitigates cell apoptosis through modulation of PI3K/AKT and ERK and JNK signaling pathways in human lung cancer cell lines. Arch. Toxicol. 2013, 87, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Xu, Y.; Han, X.; Qi, Y.; Xu, L.; Xu, Y.; Yin, L.; Sun, H.; Liu, K.; Peng, J. Anti-cancer effects of dioscin on three kinds of human lung cancer cell lines through inducing DNA damage and activating mitochondrial signal pathway. Food Chem. Toxicol. 2013, 59, 118–128. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Qiao, A.; Wang, X.; Liu, B.; Jiang, M.; Su, L.; Yao, X. Structural identification of methyl protodioscin metabolites in rats’ urine and their antiproliferative activities against human tumor cell lines. Steroids 2006, 71, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cheng, Y.; Wang, N.; Wang, D.M.; Li, Y.W.; Han, F.; Shen, J.G.; Yang de, P.; Guan, X.Y.; Chen, J.P. Dioscin induces cancer cell apoptosis through elevated oxidative stress mediated by downregulation of peroxiredoxins. Cancer Biol. Ther. 2012, 13, 138–147. [Google Scholar] [PubMed]
- Hsieh, M.J.; Yang, S.F.; Hsieh, Y.S.; Chen, T.Y.; Chiou, H.L. Autophagy inhibition enhances apoptosis induced by dioscin in HUH7 cells. Evid.-Based Complement. Altern. Med. 2012, 2012, 134512. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Li, F.R.; Jiao, P.; Yang, M.F.; Zhou, X.J.; Si, Y.H.; Jiang, W.J.; Zheng, T.T. Paris chinensis dioscin induces G2/M cell cycle arrest and apoptosis in human gastric cancer SGC-7901 cells. World J. Gastroenterol. 2011, 17, 4389–4395. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Xu, L.; Yin, L.; Qi, Y.; Li, H.; Xu, Y.; Han, X.; Peng, J.; Wan, X. Cytotoxicity of dioscin in human gastric carcinoma cells through death receptor and mitochondrial pathways. J. Appl. Toxicol. 2013, 33, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xu, L.; Yin, L.; Xu, Y.; Han, X.; Qi, Y.; Zhao, Y.; Liu, K.; Peng, J. ITRAQ-based proteomic analysis of dioscin on human HCT-116 colon cancer cells. Proteomics 2014, 14, 51–73. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Liu, M.; Wang, Z.; Ju, Y. Apoptosis induced by dioscin in hela cells. Biol. Pharm. Bull. 2002, 25, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Li, F.R.; Jiao, P.; Yao, S.T.; Sang, H.; Si, Y.H. Apoptosis of human ovarian cancer cells induced by paris chinensis dioscin via a Ca2+-mediated mitochondrion pathway. Asian Pac. J. Cancer Prev. 2011, 12, 1361–1366. [Google Scholar] [PubMed]
- Chen, J.; Li, H.M.; Zhang, X.N.; Xiong, C.M.; Ruan, J.L. Dioscin-induced apoptosis of human lncap prostate carcinoma cells through activation of caspase-3 and modulation of Bcl-2 protein family. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.A.; Jang, J.H.; Lee, Y.H.; Sung, E.G.; Song, I.H.; Kim, J.Y.; Kim, S.; Sohn, H.Y.; Lee, T.J. Dioscin induces caspase-independent apoptosis through activation of apoptosis-inducing factor in breast cancer cells. Apoptosis 2014, 19, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Qing, Y.; Wu, Y.; Hu, X.; Jiang, L.; Wu, X. Dioscin inhibits colon tumor growth and tumor angiogenesis through regulating VEGFR2 and AKT/MAPK signaling pathways. Toxicol. Appl. Pharmacol. 2014, 281, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Shuli, M.; Wenyuan, G.; Yanjun, Z.; Chaoyi, M.; Liu, Y.; Yiwen, L. Paridis saponins inhibiting carcinoma growth and metastasis in vitro and in vivo. Arch. Pharm. Res. 2011, 34, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, S.; Zheng, L.; Yin, L.; Xu, L.; Peng, J. A 90-day subchronic toxicological assessment of dioscin, a natural steroid saponin, in sprague-dawley rats. Food Chem. Toxicol. 2012, 50, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Zheng, L.; Dong, D.; Xu, L.; Yin, L.; Xu, Y.; Qi, Y.; Han, X.; Peng, J. Dioscin, a natural steroid saponin, induces apoptosis and DNA damage through reactive oxygen species: A potential new drug for treatment of glioblastoma multiforme. Food Chem. Toxicol. 2013, 59, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, Q.Y.; Chiu, J.F. Dioscin induced activation of p38 MAPK and JNK via mitochondrial pathway in HL-60 cell line. Eur. J. Pharmacol. 2014, 735, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheung, Y.H.; Yang, Z.; Chiu, J.F.; Che, C.M.; He, Q.Y. Proteomic approach to study the cytotoxicity of dioscin (saponin). Proteomics 2006, 6, 2422–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, L.; Zheng, L.; Xu, L.; Yin, L.; Han, X.; Qi, Y.; Xu, Y.; Wang, C.; Peng, J. Dioscin suppresses human laryngeal cancer cells growth via induction of cell-cycle arrest and mapk-mediated mitochondrial-derived apoptosis and inhibition of tumor invasion. Eur. J. Pharmacol. 2016, 774, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, E.A.; Park, K.G.; Lee, S.J.; Kim, M.S.; Sohn, H.Y.; Lee, T.J. Dioscin sensitizes cells to trail-induced apoptosis through downregulation of c-FLIP and Bcl-2. Oncol. Rep. 2012, 28, 1910–1916. [Google Scholar] [PubMed]
- Zhao, X.; Tao, X.; Xu, L.; Yin, L.; Qi, Y.; Xu, Y.; Han, X.; Peng, J. Dioscin induces apoptosis in human cervical carcinoma hela and siha cells through ROS-mediated DNA damage and the mitochondrial signaling pathway. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.; Serly, J.; Dinchev, D.; Ocsovszki, I.; Kostova, I.; Molnar, J. Screening of some saponins and phenolic components of tribulus terrestris and smilax excelsa as mdr modulators. In Vivo 2009, 23, 545–550. [Google Scholar] [PubMed]
- Sun, B.T.; Zheng, L.H.; Bao, Y.L.; Yu, C.L.; Wu, Y.; Meng, X.Y.; Li, Y.X. Reversal effect of dioscin on multidrug resistance in human hepatoma HepG2/adriamycin cells. Eur. J. Pharmacol. 2011, 654, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Meng, Q.; Wang, C.; Liu, Q.; Peng, J.; Huo, X.; Sun, H.; Ma, X.; Liu, K. Dioscin restores the activity of the anticancer agent adriamycin in multidrug-resistant human leukemia k562/adriamycin cells by down-regulating MDR1 via a mechanism involving NF-κB signaling inhibition. J. Natl. Prod. 2013, 76, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Qiu, Z.; Wang, Y.; Zhang, Y.; Li, M.; Yu, J.; Zhang, L.; Zhu, Z.; Yu, B. Synthesis, cytotoxicity, and hemolytic activity of 6′-O-substituted dioscin derivatives. Carbohydr. Res. 2007, 342, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Aumsuwan, P.; Khan, S.I.; Khan, I.A.; Ali, Z.; Avula, B.; Walker, L.A.; Shariat-Madar, Z.; Helferich, W.G.; Katzenellenbogen, B.S.; Dasmahapatra, A.K. The anticancer potential of steroidal saponin, dioscin, isolated from wild yam (Dioscorea villosa) root extract in invasive human breast cancer cell line MDA-MB-231 in vitro. Arch. Biochem. Biophys. 2016, 591, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Cai, H.; Meng, X. Polyphyllin D induces apoptosis and differentiation in K562 human leukemia cells. Int. Immunopharmacol. 2016, 36, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, Q.; Liu, Y. Polyphyllin D induces apoptosis in K562/A02 cells through G2/M phase arrest. J. Pharm. Pharmacol. 2014, 66, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Yuet-Wa, J.C.; Kong, S.K.; Yu, B.; Eng-Choon, V.O.; Nai-Ching, H.W.; Chung-Wai, T.M.; Fung, K.P. Effects of polyphyllin D, a steroidal saponin in Paris polyphylla, in growth inhibition of human breast cancer cells and in xenograft. Cancer Biol. Ther. 2005, 4, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Siu, F.M.; Ma, D.L.; Cheung, Y.W.; Lok, C.N.; Yan, K.; Yang, Z.; Yang, M.; Xu, S.; Ko, B.C.; He, Q.Y.; et al. Proteomic and transcriptomic study on the action of a cytotoxic saponin (polyphyllin D): Induction of endoplasmic reticulum stress and mitochondria-mediated apoptotic pathways. Proteomics 2008, 8, 3105–3117. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.Y.; Ong, R.C.; Suen, Y.K.; Ooi, V.; Wong, H.N.; Mak, T.C.; Fung, K.P.; Yu, B.; Kong, S.K. Polyphyllin D is a potent apoptosis inducer in drug-resistant HepG2 cells. Cancer Lett. 2005, 217, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Li, Q.; Lu, P.; Chen, Q. Polyphyllin D induces apoptosis in U87 human glioma cells through the c-Jun NH2-terminal kinase pathway. J. Med. Food 2014, 17, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Darbour, N.; Bayet, C.; Doreau, A.; Raad, I.; Phung, B.H.; Dumontet, C.; Di Pietro, A.; Dijoux-Franca, M.G.; Guilet, D. Selective modulation of P-glycoprotein activity by steroidal saponines from Paris polyphylla. Fitoterapia 2009, 80, 39–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Sawah, E.; Marchion, D.C.; Xiong, Y.; Ramirez, I.J.; Abbasi, F.; Boac, B.M.; Bush, S.H.; Bou Zgheib, N.; McClung, E.C.; Khulpateea, B.R.; et al. The chinese herb polyphyllin D sensitizes ovarian cancer cells to cisplatin-induced growth arrest. J. Cancer Res. Clin. Oncol. 2015, 141, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ong, R.C.; Lei, J.; Lee, R.K.; Cheung, J.Y.; Fung, K.P.; Lin, C.; Ho, H.P.; Yu, B.; Li, M.; Kong, S.K. Polyphyllin D induces mitochondrial fragmentation and acts directly on the mitochondria to induce apoptosis in drug-resistant HepG2 cells. Cancer Lett. 2008, 261, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.D.; Lu, H.X.; Xu, L.S.; Xiao, W. Polyphyllin D exerts potent anti-tumour effects on lewis cancer cells under hypoxic conditions. J. Int. Med. Res. 2009, 37, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.Y.; Koon, J.C.; Liu, X.; Detmar, M.; Yu, B.; Kong, S.K.; Fung, K.P. Polyphyllin D, a steroidal saponin from Paris polyphylla, inhibits endothelial cell functions in vitro and angiogenesis in zebrafish embryos in vivo. J. Ethnopharmacol. 2011, 137, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Tao, L.; Chau, S.L.; Wu, R.; Zhang, H.; Yang, Y.; Yang, D.; Bian, Z.; Lu, A.; Han, Q.; et al. Folding fan mode counter-current chromatography offers fast blind screening for drug discovery. Case study: Finding anti-enterovirus 71 agents from Anemarrhena asphodeloides. J. Chromatogr. A 2014, 1368, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.P.; Zhang, J.; Cong, Y.; Li, B.; Xiong, C.Q.; Zhao, Y.; Tan, D.W.; Yu, H.S.; Yu, Z.Y.; Cong, Y.W.; et al. Steroidal glycosides from the rhizomes of Anemarrhena asphodeloides and their antiplatelet aggregation activity. Planta Medica 2012, 78, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.Q.; Qiu, Y.; Li, T.J.; Tao, X.; Sun, L.N.; Chen, W.S. Antiplatelet and antithrombotic activities of timosaponin B-II, an extract of Anemarrhena asphodeloides. Clin. Exp. Pharmacol. Physiol. 2011, 38, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Lin, L.C.; Chen, C.F.; Cheng, J.S.; Lo, Y.K.; Chou, K.J.; Lee, K.C.; Liu, C.P.; Wu, Y.Y.; Su, W.; et al. Effect of timosaponin A-III, from Anemarrhenae asphodeloides Bunge (Liliaceae), on calcium mobilization in vascular endothelial and smooth muscle cells and on vascular tension. Life Sci. 2002, 71, 1081–1090. [Google Scholar] [CrossRef]
- Chen, J.R.; Jia, X.H.; Wang, H.; Yi, Y.J.; Wang, J.Y.; Li, Y.J. Timosaponin A-III reverses multi-drug resistance in human chronic myelogenous leukemia K562/ADM cells via downregulation of MDR1 and MRP1 expression by inhibiting PI3K/AKT signaling pathway. Int. J. Oncol. 2016, 48, 2063–2070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, J.; Wang, S.; Ma, C. Timosaponin B-II inhibits lipopolysaccharide-induced acute lung toxicity via TLR/NF-κB pathway. Toxicol. Mech. Methods 2015, 25, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Li, T.J.; Qiu, Y.; Yang, P.Y.; Rui, Y.C.; Chen, W.S. Timosaponin B-II improves memory and learning dysfunction induced by cerebral ischemia in rats. Neurosci. Lett. 2007, 421, 147–151. [Google Scholar] [CrossRef] [PubMed]
- King, F.W.; Fong, S.; Griffin, C.; Shoemaker, M.; Staub, R.; Zhang, Y.L.; Cohen, I.; Shtivelman, E. Timosaponin A-III is preferentially cytotoxic to tumor cells through inhibition of mtor and induction of er stress. PLoS ONE 2009, 4, e7283. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Chung, H.J.; Nam, J.W.; Park, H.J.; Seo, E.K.; Kim, Y.S.; Lee, D.; Lee, S.K. Cytotoxic and antineoplastic activity of timosaponin A-III for human colon cancer cells. J. Natl. Prod. 2011, 74, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Zhao, X.C.; Wang, S.J.; Gao, P.Y.; Li, L.Z.; Ikejima, T.; Song, S.J. Synthesis and biological evaluation of novel sarsasapogenin derivatives as potential anti-tumor agents. Steroids 2015, 93, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lok, C.N.; Sy, L.K.; Liu, F.; Che, C.M. Activation of autophagy of aggregation-prone ubiquitinated proteins by timosaponin A-III. J. Biol. Chem. 2011, 286, 31684–31696. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Chiang, W.L.; Hsiao, P.C.; Chien, M.H.; Chen, H.Y.; Weng, W.C.; Hsieh, M.J.; Yang, S.F. Timosaponin aiii mediates caspase activation and induces apoptosis through JNK1/2 pathway in human promyelocytic leukemia cells. Tumour Biol. 2015, 36, 3489–3497. [Google Scholar] [CrossRef] [PubMed]
- Sy, L.K.; Yan, S.C.; Lok, C.N.; Man, R.Y.; Che, C.M. Timosaponin A-III induces autophagy preceding mitochondria-mediated apoptosis in hela cancer cells. Cancer Res. 2008, 68, 10229–10237. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Yang, C.W.; Wang, J.Y.; Tsai, Y.F.; Tseng, L.M.; King, K.L.; Chen, W.S.; Chiu, J.H.; Shyr, Y.M. Timosaponin AIII suppresses hepatocyte growth factor-induced invasive activity through sustained ERK activation in breast cancer MDA-MB-231 cells. Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Im, A.R.; Kim, S.H.; Hyun, J.W.; Chae, S. Timosaponin AIII inhibits melanoma cell migration by suppressing COX-2 and in vivo tumor metastasis. Cancer Sci. 2016, 107, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yu, Y.; Wang, L.; Wu, B.; Xia, L.; Feng, F.; Ling, Z.; Wang, S. Additive antiangiogenesis effect of ginsenoside Rg3 with low-dose metronomic temozolomide on rat glioma cells both in vivo and in vitro. J. Exp. Clin. Cancer Res. 2016, 35, 32. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.K.; Zhang, M.M.; Zhou, H.; Lam, K.Y.; Chan, P.L.; Law, C.K.; Yue, P.Y.; Liu, L. Saikosaponin-D enhances the anticancer potency of TNF-α via overcoming its undesirable response of activating NF-κB signalling in cancer cells. Evid.-Based Complement. Altern. Med. 2013, 2013, 745295. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Wang, J.; Wang, Y.; Wang, Y.; Cai, J.; Wang, M.; Chen, Q.; Song, J.; Yu, Z.; Huang, W.; et al. Inhibition of autophagy potentiates anticancer property of 20(S)-ginsenoside Rh2 by promoting mitochondria-dependent apoptosis in human acute lymphoblastic leukaemia cells. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Liu, F.; Qian, W.; Zhang, T.; Li, F. Combined effect of sodium selenite and ginsenoside Rh2 on HCT116 human colorectal carcinoma cells. Arch. Iran. Med. 2016, 19, 23–29. [Google Scholar] [PubMed]
- Chen, F.; Deng, Z.Y.; Zhang, B.; Xiong, Z.X.; Zheng, S.L.; Tan, C.L.; Hu, J.N. Esterification of ginsenoside Rh2 enhanced its cellular uptake and antitumor activity in human HepG2 cells. J. Agric. Food Chem. 2016, 64, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Nakata, H.; Kikuchi, Y.; Tode, T.; Hirata, J.; Kita, T.; Ishii, K.; Kudoh, K.; Nagata, I.; Shinomiya, N. Inhibitory effects of ginsenoside Rh2 on tumor growth in nude mice bearing human ovarian cancer cells. Jpn. J. Cancer Res. 1998, 89, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shimizu, K.; Yu, H.; Zhang, C.; Jin, F.; Kondo, R. Stereospecificity of hydroxyl group at c-20 in antiproliferative action of ginsenoside Rh2 on prostate cancer cells. Fitoterapia 2010, 81, 902–905. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, H.; Rayburn, E.R.; Zhao, Y.; Hill, D.L.; Zhang, R. 20(S)-25-methoxyl-dammarane-3β,12β, 20-triol, a novel natural product for prostate cancer therapy: Activity in vitro and in vivo and mechanisms of action. Br. J. Cancer 2008, 98, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, C.; Xue, R.; Jiang, W.; Wu, B.; Huang, C. Cytotoxic activities of chemical constituents from rhizomes of Anemarrhena asphodeloides and their analogues. Arch. Pharm. Res. 2015, 38, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, M.; Wu, X.; Hu, Y.; Cao, Y.; Wang, X.; Xiang, S.; Li, H.; Jiang, L.; Tan, Z.; et al. 20(S)-ginsenoside Rg3 promotes senescence and apoptosis in gallbladder cancer cells via the p53 pathway. Drug Des. Dev. Ther. 2015, 9, 3969–3987. [Google Scholar]
- Zhao, X.; Xu, L.; Zheng, L.; Yin, L.; Qi, Y.; Han, X.; Xu, Y.; Peng, J. Potent effects of dioscin against gastric cancer in vitro and in vivo. Phytomedicine 2016, 23, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zou, J.; Bui-Nguyen, T.M.; Bai, P.; Gao, L.; Liu, J.; Liu, S.; Xiao, J.; Chen, X.; Zhang, X.; et al. Paris saponin ii of rhizoma paridis—A novel inducer of apoptosis in human ovarian cancer cells. Biosci. Trends 2012, 6, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Liao, C.; Li, Y.; Fan, X.; Fan, L.; Xu, H.; Kang, Q.; Zeng, Y.; Wu, X.; Wu, H.; et al. Gypenoside l, isolated from Gynostemma pentaphyllum, induces cytoplasmic vacuolation death in hepatocellular carcinoma cells through reactive-oxygen-species-mediated unfolded protein response. J. Agric. Food Chem. 2016, 64, 1702–1711. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Zheng, K.; Li, Y.; Xu, H.; Kang, Q.; Fan, L.; Hu, X.; Jin, Z.; Zeng, Y.; Kong, X.; et al. Gypenoside l inhibits autophagic flux and induces cell death in human esophageal cancer cells through endoplasm reticulum stress-mediated Ca2+ release. Oncotarget 2016. [Google Scholar] [CrossRef]
- Chen, M.; Guo, Y.; Zhao, R.; Wang, X.; Jiang, M.; Fu, H.; Zhang, X. Ophiopogonin B induces apoptosis, mitotic catastrophe and autophagy in A549 cells. Int. J. Oncol. 2016, 49, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Wu, L.; Wang, J.L.; Yang, J.D.; Zhang, J.; Zhang, J.; Li, L.H.; Xia, Y.; Yao, L.B.; Qin, H.Z.; et al. Astragaloside iv prevents MPP+-induced SH-SY5Y cell death via the inhibition of bax-mediated pathways and ROS production. Mol. Cell. Biochem. 2012, 364, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, W.S.; Wu, G.S.; Chen, X.P.; Wang, Y.T.; Lu, J.J. Platycodin D induces apoptosis, and inhibits adhesion, migration and invasion in HepG2 hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, X.H.; Tang, Z.H.; Wang, Y.F.; Leung, C.H.; Ma, D.L.; Chen, X.P.; Wang, Y.T.; Chen, Y.; Lu, J.J. Platycodin D induces apoptosis and triggers ERK- and JNK-mediated autophagy in human hepatocellular carcinoma Bel-7402 cells. Acta Pharmacol. Sin. 2015, 36, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tang, Z.H.; Xu, W.S.; Wu, G.S.; Wang, Y.F.; Chang, L.L.; Zhu, H.; Chen, X.P.; Wang, Y.T.; Chen, Y.; et al. Platycodin D triggers autophagy through activation of extracellular signal-regulated kinase in hepatocellular carcinoma HepG2 cells. Eur. J. Pharmacol. 2015, 749, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.H.; Li, T.; Gao, H.W.; Sun, W.; Chen, X.P.; Wang, Y.T.; Lu, J.J. Platycodin D from Platycodonis radix enhances the anti-proliferative effects of doxorubicin on breast cancer MCF-7 and MDA-MB-231 cells. Chin. Med. 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.J.; Lu, D.Z.; Chen, Y.F.; Dong, Y.T.; Zhang, J.R.; Li, T.; Tang, Z.H.; Yang, Z. Proteomic analysis of hepatocellular carcinoma HepG2 cells treated with platycodin D. Chin. J. Natl. Med. 2015, 13, 673–679. [Google Scholar] [CrossRef]
- Lee, Y.J.; Son, Y.M.; Gu, M.J.; Song, K.D.; Park, S.M.; Song, H.J.; Kang, J.S.; Woo, J.S.; Jung, J.H.; Yang, D.C.; et al. Ginsenoside fractions regulate the action of monocytes and their differentiation into dendritic cells. J. Ginseng Res. 2015, 39, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Yuan, Q.; Cui, Q.; Liu, C.; Zhou, Z.; Zhao, H.; Dun, Y.; Wang, T.; Zhang, C. Vaccine adjuvant ginsenoside Rg1 enhances immune responses against hepatitis B surface antigen in mice. Can. J. Physiol. Pharmacol. 2016, 94, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wang, J.; Shi, M.; Liu, W.; Yang, Z.; Fu, Y. Saikosaponin a inhibits LPS-induced inflammatory response by inducing liver X receptor α activation in primary mouse macrophages. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tian, Y.H.; Liu, Y.; Wang, Z.; Tang, S.; Zhang, J.; Wang, Y.P. Platycodin D exerts anti-tumor efficacy in H22 tumor-bearing mice via improving immune function and inducing apoptosis. J. Toxicol. Sci. 2016, 41, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Wisloff, H.; Uhlig, S.; Scheie, E.; Loader, J.; Wilkins, A.; Flaoyen, A. Toxicity testing of saponin-containing Yucca schidigera Roetzl. juice in relation to hepato- and nephrotoxicity of Narthecium ossifragum (L.) Huds. Toxicon 2008, 51, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Chen, F.; Li, C. Absorption, disposition, and pharmacokinetics of saponins from Chinese medicinal herbs: What do we know and what do we need to know more? Curr. Drug Metab. 2012, 13, 577–598. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Basu, S.; Yang, Z.; Deb, A.; Hu, M. Bioavailability challenges associated with development of saponins as therapeutic and chemopreventive agents. Curr. Drug Targets 2012, 13, 1885–1899. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; He, Y.; Sun, Y.; Zheng, B.; Liu, Y.; Wang, X.; Zhang, Y.; Lee, R.J.; Teng, L.; Xie, J. Improvement of oral availability of ginseng fruit saponins by a proliposome delivery system containing sodium deoxycholate. Saudi J. Biol. Sci. 2016, 23, S113–S125. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Yang, H.; Yoon, I.S.; Kim, S.B.; Ko, S.H.; Shim, J.S.; Sung, S.H.; Cho, H.J.; Kim, D.D. Nanocomplexes based on amphiphilic hyaluronic acid derivative and polyethylene glycol-lipid for ginsenoside Rg3 delivery. J. Pharm. Sci. 2014, 103, 3254–3262. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compounds | Tissue Types | Cell Lines | Effects | Reference |
|---|---|---|---|---|
| Triterpenoid Saponins | ||||
| Ginsenoside Rg3 | Lung cancer | H460 | IC50 = 392 μM/24 h | [23] |
| Esophageal carcinoma | Eca-109 | IC50 > 127 μM/48 h | [24] | |
| Gastric cancer | AGS | IC50 = 31 μM/24 h | [25] | |
| Colon cancer | HCT116 | IC50 > 100 μM/24 h | [26] | |
| Colon cancer | HCT116 | IC50 = 100 μM/48 h | [27] | |
| Colon cancer | HT29 | IC50 = 100 μM/48 h | [28] | |
| Colon cancer | SW620 | IC50 = 100 μM/24 h | [26] | |
| Hepatocellular carcinoma | Hep1-6 | IC50 > 127 μM/24 h | [29] | |
| Hepatocellular carcinoma | HepG2 | IC50 < 64 μM/24 h | [30] | |
| Renal carcinoma | 786-0 | IC50 > 127 μM/48 h | [31] | |
| Bladder cancer | T24R2 | IC50 = 265 μM/48 h | [32] | |
| Breast cancer | MDA-MB-231 | IC50 < 38 μM/24 h | [33] | |
| Ovarian cancer | SKOV3 | IC50 = 187 μM/48 h | [34] | |
| Ovarian cancer | 3AO | IC50 = 309 μM/48 h | [34] | |
| Prostate cancer | PC-3 | IC50 = 85.8 μM/48 h | [35] | |
| Prostate cancer | PC-3 | IC50 = 91.3 μM/24 h | [35] | |
| Prostate cancer | PC-3 | EC50 = 14.1 μM/24 h | [36] | |
| Prostate cancer | DU145 | IC50 = 81.5 μM/24 h | [35] | |
| Prostate cancer | DU145 | IC50 = 73.1 μM/48 h | [35] | |
| Prostate cancer | LNCaP | EC50 = 8.4 μM/24 h | [36] | |
| Prostate cancer | LNCaP | IC50 = 74.7 μM/24 h | [35] | |
| Prostate cancer | LNCaP | IC50 = 62.2 μM/48 h | [35] | |
| Melanoma | B16 | IC50 = 92 μM/48 h | [38] | |
| Melanoma | C8161 | IC50 = 71 μM/24 h | [39] | |
| Melanoma | C8161 | IC50 = 64 μM/48 h | [39] | |
| Melanoma | C8161 | IC50 = 63 μM/72 h | [39] | |
| Melanoma | A375 | IC50 = 64 μM/24 h | [39] | |
| Melanoma | A375 | IC50 = 54 μM/48 h | [39] | |
| Melanoma | A375 | IC50 = 41 μM/72 h | [39] | |
| Ginsenoside Rh2 | Leukemia | THP-1 | IC50 = 24 μM/72 h | [70] |
| Leukemia | HL-60 | IC50 = 25.0 μM/48 h | [71] | |
| Leukemia | Reh | IC50 = 40 μM/24 h | [190] | |
| Leukemia | Jurkat | IC50 = 35 μM/24 h | [190] | |
| Colon cancer | HCT-116 | IC50 = 50 μM/24 h | [191] | |
| Colon cancer | HCT-116 | IC50 = 35 μM/48 h | [73] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 42.12 μM/24 h | [192] | |
| Hepatocellular carcinoma | HepG2 | IC50 < 16 μM/24 h | [30] | |
| Breast cancer | MCF-7 | IC50 = 50 μM/24 h | [74] | |
| Breast cancer | MCF-7 | IC50 > 20 μM/24 h | [75] | |
| Breast cancer | MDA-MB-231 | IC50 = 50 μM/24 h | [74] | |
| Breast cancer | MDA-MB-231 | IC50 > 40 μM/24 h | [75] | |
| Ovarian cancer | KF | IC50 = 40 μM/5 days | [193] | |
| Ovarian cancer | KFr | IC50 = 41 μM/5 days | [193] | |
| Ovarian cancer | HRA | IC50 = 30 μM/5 days | [193] | |
| Ovarian cancer | KK | IC50 = 45 μM/5 days | [193] | |
| Prostate cancer | PC-3 | EC50 = 5.5 μM/24 h | [36] | |
| Prostate cancer | PC-3 | IC50 = 35 μM/72 h | [194] | |
| Prostate cancer | PC-3 | IC50 = 35.7 μM/72 h | [195] | |
| Prostate cancer | LNCaP | EC50 = 4.4 μM/24 h | [36] | |
| Prostate cancer | LNCaP | IC50 = 46.7 μM/72 h | [195] | |
| Prostate cancer | LNCaP | IC50 = 17 μM/72 h | [194] | |
| Prostate cancer | DU145 | IC50 = 38 μM/72 h | [194] | |
| Saikosaponin A | Gastric cancer | AGS | IC50 = 34.6 μM/24 h | [112] |
| Colon cancer | HCT116 | IC50 < 20 μM/40 h | [117] | |
| Colon cancer | LoVo | IC50 < 20 μM/40 h | [117] | |
| Colon cancer | SW48 | IC50 < 20 μM/40 h | [117] | |
| Colon cancer | SW480 | IC50 about 20 μM/40 h | [117] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 13 μM/24 h | [113] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 23.4 μM/24 h | [112] | |
| Breast cancer | MDA-MB-231 | ED50 = 6.4 μM/48 h | [115] | |
| Breast cancer | MCF-7 | ED50 = 6.4 μM/48 h | [115] | |
| Breast cancer | MCF-7 | IC50 = 33.3 μM/24 h | [112] | |
| Saikosaponin D | Lung cancer | A549 | IC50 = 10.18 μM/48 h | [119] |
| Hepatocellular carcinoma | HepG2 | IC50 = 2.63 μM/48 h | [121] | |
| Hepatocellular carcinoma | Hep3B | IC50 = 4.26 μM/48 h | [121] | |
| Hepatocellular carcinoma | SMMC-7721 | IC50 > 15 μM/72 h | [127] | |
| Prostate cancer | DU145 | IC50 about 10 μM/24 h | [123] | |
| Thyroid cancer | ARO | IC50 about 20 μM/24 h | [124] | |
| Thyroid cancer | 8305C | IC50 about 15 μM/24 h | [124] | |
| Thyroid cancer | SW1736 | IC50 about 18 μM/24 h | [124] | |
| Steroid Saponins | ||||
| Dioscin | Leukemia | K562 | IC50 = 4.7 μM/48 h | [159] |
| Lung cancer | NCI-H460 | IC50 = 18.2 μM/72 h | [138] | |
| Lung cancer | NCI-H446 | IC50 about 12 μM/48 h | [137] | |
| Lung cancer | H1299 | IC50 about 2.5 μM/24 h | [136] | |
| Lung cancer | A549 | IC50 about 6 μM/48 h | [137] | |
| Lung cancer | A549 | IC50 < 2.5 μM/24 h | [136] | |
| Gastric carcinoma | SGC-7901 | IC50 = 16 μM/24 h | [141] | |
| Gastric carcinoma | SGC-7901 | IC50 = 10 μM/48 h | [141] | |
| Gastric carcinoma | SGC-7901 | IC50 = 4 μM/72 h | [141] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 8.3 μM/72 h | [138] | |
| Cervical cancer | HeLa | IC50 = 4.4 μM/48 h | [144] | |
| Cervical cancer | HeLa | IC50 = 40.2 μM/72 h | [138] | |
| Breast cancer | MCF-7 | IC50 = 50.6 μM/72 h | [138] | |
| Polyphyllin D | Leukemia | K562 | IC50 = 0.9 μM/24 h | [163] |
| Leukemia | K562/A02 | IC50 = 0.8 μM/24 h | [163] | |
| Lung cancer | NCI-H460 | IC50 = 3.0 μM/48 h | [165] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 7 μM/24 h | [166] | |
| Hepatocellular carcinoma | HepG2 | IC50 = 3.5 μM/24 h | [165] | |
| Hepatocellular carcinoma | R-HepG2 | IC50 = 5 μM/24 h | [166] | |
| Ovarian cancer | A2780CP | EC50 = 0.22 μM/72 h | [169] | |
| Ovarian cancer | TYKNU-CIS-R | EC50 = 0.25 μM/72 h | [169] | |
| Ovarian cancer | TYKNU | EC50 = 0.28 μM/72 h | [169] | |
| Ovarian cancer | TOV112D | EC50 = 0.30 μM/72 h | [169] | |
| Ovarian cancer | HEYA8 | EC50 = 0.36 μM/72 h | [169] | |
| Ovarian cancer | TOV21G | EC50 = 0.37 μM/72 h | [169] | |
| Ovarian cancer | A2780S | EC50 = 0.37 μM/72 h | [169] | |
| Ovarian cancer | IMCC5 | EC50 = 0.39 μM/72 h | [169] | |
| Ovarian cancer | OVCAR8 | EC50 = 0.39 μM/72 h | [169] | |
| Ovarian cancer | M41-R | EC50 = 0.40 μM/72 h | [169] | |
| Ovarian cancer | SKOV3 | EC50 = 0.46 μM/72 h | [169] | |
| Ovarian cancer | M41 | EC50 = 0.70 μM/72 h | [169] | |
| Ovarian cancer | PEO1 | EC50 = 0.81 μM/72 h | [169] | |
| Ovarian cancer | OVCAR2 | EC50 = 0.85 μM/72 h | [169] | |
| Ovarian cancer | OVCA433 | EC50 = 0.97 μM/72 h | [169] | |
| Ovarian cancer | IGROV1 | EC50 = 0.97 μM/72 h | [169] | |
| Ovarian cancer | OVCAR5 | EC50 = 1.10 μM/72 h | [169] | |
| Ovarian cancer | OVCA432 | EC50 = 1.10 μM/72 h | [169] | |
| Ovarian cancer | OVCA420 | EC50 = 1.27 μM/72 h | [169] | |
| Ovarian cancer | MCAS | EC50 = 1.44 μM/72 h | [169] | |
| Cervical cancer | HeLa | IC50 = 3.7 μM/48 h | [165] | |
| Breast cancer | MCF-7 | IC50 = 5 μM/48 h | [164] | |
| Breast cancer | MCF-7 | IC50 = 3.7 μM/48 h | [165] | |
| Breast cancer | MDA-MB-231 | IC50 = 2.5 μM/48 h | [164] | |
| Glioblastoma | SF-268 | IC50 = 3.0 μM/48 h | [165] | |
| Glioma | U87 | IC50 = 49.4 μM/24 h | [167] | |
| Timosaponin AIII | Colon cancer | HCT15 | IC50 = 6.1 μM/72 h | [181] |
| Colon cancer | HCT116 | IC50 = 5.5 μM/72 h | [181] | |
| Colon cancer | HT29 | IC50 = 10.3 μM/72 h | [181] | |
| Colon cancer | HT29 | IC50 = 2.2 μM/72 h | [196] | |
| Colon cancer | SW480 | IC50 = 13.1 μM/72 h | [181] | |
| Colon cancer | SW620 | IC50 = 11.1 μM/72 h | [181] | |
| Hepatocellular carcinoma | BEL-7402 | IC50 = 1.65 μM/72 h | [196] | |
| Cervical cancer | HeLa | IC50 = 9.63 μM/72 h | [196] | |
| Breast cancer | MDA-MB-468 | IC50 = 1.6 μM/72 h | [196] | |
| Compounds | Method | Effect | Reference |
|---|---|---|---|
| Triterpenoid Saponins | |||
| Ginsenoside Rg3 | Daily oral administration of 20 mg/kg ginsenoside Rg3 for 18 days in C57BL/6 mice bearing Lewis lung carcinoma cells | Enhanced about 50% of survival rate and delayed about 33.3% of tumor growth without side effects | [44] |
| 28-day oral treatment with 100 mg/kg ginsenoside Rg3 in nude mice bearing H460 cells | Remarkably suppressed the tumor growth by decreasing the tumor volume and weight by 30%–31% | [23] | |
| Daily oral administration of 20 mg/kg ginsenoside Rg3 for 21 days in nude mice bearing Huh-7 cells | Reduced tumor volume for 23%. | [48] | |
| H22-bearing mice were injected intraperitoneally with 20(S)-Rg3 and 20(R)-Rg3 (3 mg/kg body weight) once a day for 10 days | Inhibited the 23.6% and 40.9% of tumor growth, respectively. And enhanced cellular immunity with lymphocyte proliferation and IL-2 and IFN-γ production in serum and immune organs | [51] | |
| Daily intra-tumor injection of ginsenoside Rg3 (3.0 mg/kg) for ten days in C57BL/6 mice bearing Hep1-6 cells | Inhibited the tumor growth by more than 50% and prolonged survival time. | [29] | |
| Rg3 was administered at 20 mg/kg body weight to nude mice bearing HCT116 cells daily for 3 weeks via i.p. injection | Inhibited about 70% of the tumor growth by down-regulating Wnt/beta-catenin signaling pathway | [27] | |
| Rg3 was injected intraperitoneally at 20 or 40 mg/kg body weight every day for 3 weeks to gallbladder cancer NOZ-bearing BALB/c nude mice | Effectively reduced tumor growth for about 60% of tumor weight | [197] | |
| From day 1, 5 mg/kg of Rg3 was injected via tail-vein of SKOV3-bearing mice every other day till day 30 | Effectively reduced tumor growth for about 65% of tumor weight | [34] | |
| Daily intraperitoneal injection of 3 mg/kg ginsenoside Rg3 for 10 days in athymic mice bearing SKOV-3 cells | Prolonged 74.3% of survival time, decreased 41.9% of tumor weight, and improved life quality | [41] | |
| Rg3 was subcutaneously administered at 20 mg/kg body weight 3 weeks with time interval of 48 h to nude mice bearing melanoma A375 cells | Significantly inhibited the tumor volume by 52.50% | [53] | |
| Rg3 was administered at 20 mg/kg body weight 5 times per week for 3 weeks via i.p. injection to nude mice bearing A375 cells | Significantly reduced tumor volume by 55.65% | [39] | |
| Ginsenoside Rh2 | Daily oral administration of ginsenoside Rh2 at 20 mg/kg for 3 weeks in nude mice bearing K562 cells | Significantly inhibited the tumor volume by about 50% | [86] |
| Daily gavaged with ginsenoside Rh2(S) and (R) at 2–6 mg/kg for 10 days in H22 hepatoma-bearing mice | 4 mg/kg of ginsenoside Rh2(S) and (R) suppressed 42.2% and 46.8% of tumor growth without causing side effects | [87] | |
| Ginsenoside Rh2 was intravenously administrated at a concentration of 1 mg/kg body weight to the mice bearing reporter-carrying PC3-luc cells, twice per week for 4 weeks | The bioluminescence levels were 83.5% ± 10.5% lower than those in control group | [88] | |
| Daily oral gavage of 120 mg/kg ginsenoside Rh2 for 25 days in nude mice bearing PC-3 cells | Effectively delayed about 60% of tumor growth in terms of tumor volume without any overt toxicity | [77] | |
| Intravenous injection of 1 mg/kg ginsenoside Rh2 twice a week for 1 month in NOD/SCID mice bearing A-172 gliobalastoma cells | The bioluminescence levels were 76.8% ± 12.5% lower than those in control group | [89] | |
| Saikosaponin D | Saikosaponin D was intraperitoneally injected at a concentration of 0.75 mg/kg body weight to the BALB/c nude mice bearing SMMC-7721 xenograft tumor, thrice a week for two weeks | Saikosaponin D treatment reduced tumor volume by 11%, while the combination with radiation therapy reduced tumor volume by 66% | [125] |
| Saikosaponin D was daily intraperitoneally injected at a concentration of 2 mg/kg body weight for 17 weeks to the SD rats, starting 1 week before diethylinitrosamine induction | Saikosaponin D treatment reduced about 85% nodules at the surface of the liver without invasion to surrounding tissues | [126] | |
| Steroid Saponins | |||
| Dioscin | Orally administrated 30 mg/kg dioscin in SD rat allograft with C6 cells | The average survival time of rats in the model group was 31.5 days compared to 49.97 days in the dioscin-treated group | [151] |
| Dioscin was oral administrated at the doses of 40 and 80 mg/kg body weight for 30 days to the BALB/c nude mice bearing reporter-carrying MGC-803-luc cells | Inhibited about 43% and 59% of tumor weight, respectively | [198] | |
| Polyphyllin D | Daily administration of 2.73 mg/kg body weight through intravenous injection for ten days in nude mice bearing MCF-7 cells | Effectively reduced tumor growth for 50% in terms of tumor weight and size, given no significant toxicity in heart and liver to the host | [164] |
| One week after implantation, treatment groups received their first doses of polyphyllin D (15 or 25 mg/kg body weight) and intraperitoneal administrations were carried out on 4 consecutive days per week for 4 weeks in nude mice bearing SKOV3 cells | Administration of polyphyllin D led to a 40% (15 mg/kg) and 64% (25 mg/kg) tumor growth inhibition, respectively | [199] | |
| Timosaponin AIII | Treatment with timosaponin AIII (2 or 5 mg/kg body weight, three times/week, i.p. administration) for 4 weeks in nude mice bearing HCT-15 cells | It suppressed tumor growth without any overt toxicity. The inhibition rates of tumor size compared with control volume were 8.3% (2 mg/kg) and 37.3% (5 mg/kg) | [181] |
| C57/BL mice injected with B16-F10 melanoma cells were treated with single dose of timosaponin AIII (25 mg/kg body weight) and anatomized fourteen days later | It reduced about 50% of metastasis of melanoma cells to lung in mice, and inhibited the transcription of COX-2 and NF-κB | [187] | |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.-H.; Li, T.; Fong, C.M.V.; Chen, X.; Chen, X.-J.; Wang, Y.-T.; Huang, M.-Q.; Lu, J.-J. Saponins from Chinese Medicines as Anticancer Agents. Molecules 2016, 21, 1326. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101326
Xu X-H, Li T, Fong CMV, Chen X, Chen X-J, Wang Y-T, Huang M-Q, Lu J-J. Saponins from Chinese Medicines as Anticancer Agents. Molecules. 2016; 21(10):1326. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101326
Chicago/Turabian StyleXu, Xiao-Huang, Ting Li, Chi Man Vivienne Fong, Xiuping Chen, Xiao-Jia Chen, Yi-Tao Wang, Ming-Qing Huang, and Jin-Jian Lu. 2016. "Saponins from Chinese Medicines as Anticancer Agents" Molecules 21, no. 10: 1326. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101326