
Production of Laccase by a New Myrothecium verrucaria MD-R-16 Isolated from Pigeon Pea [Cajanus cajan (L.) Millsp.] and its Application on Dye Decolorization

Abstract
:1. Introduction
2. Results and Discussion
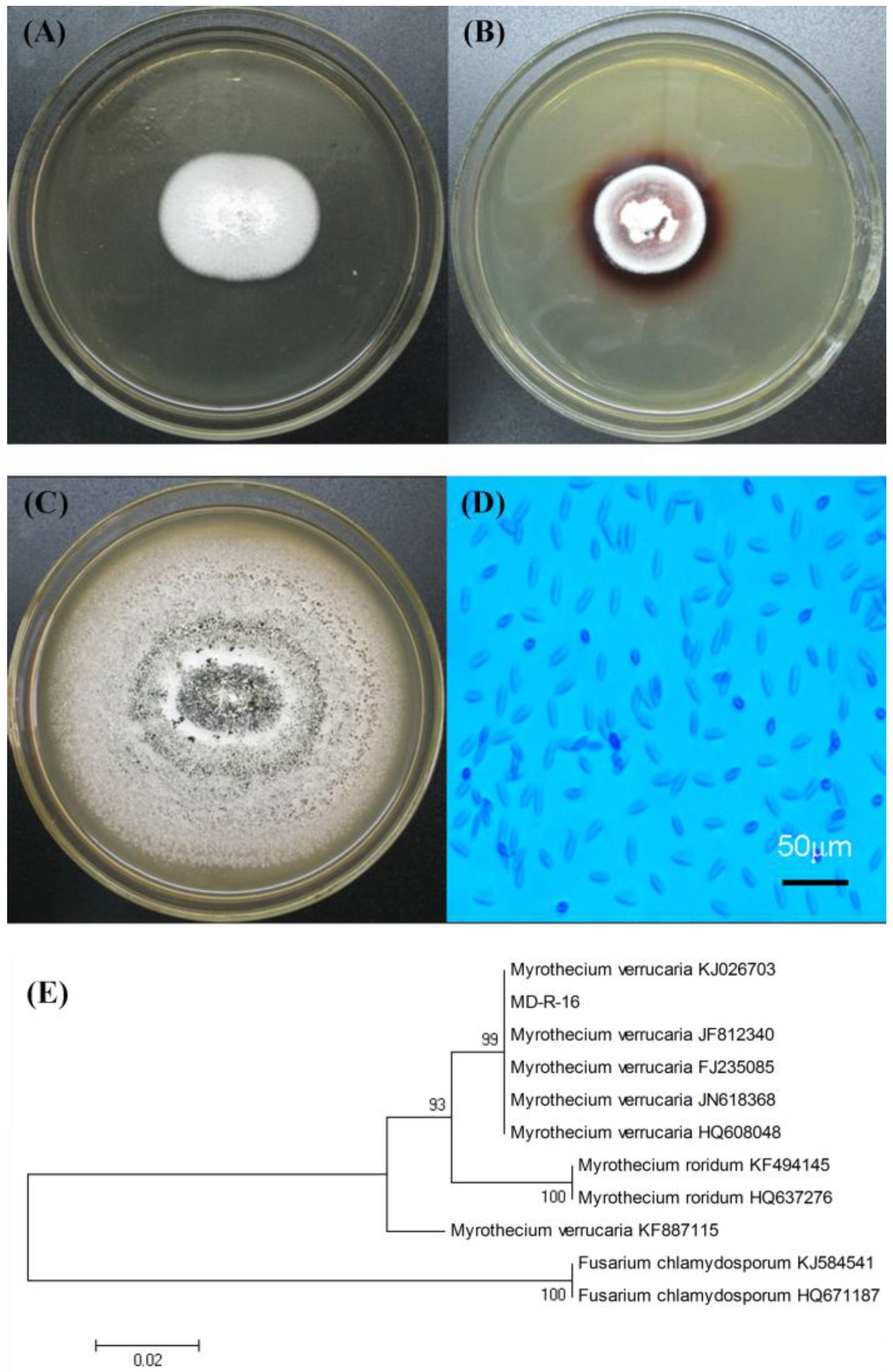
2.1. Isolation and Identification of Laccase-Producing Fungal Endophyte
2.2. Determination of Laccase Activity
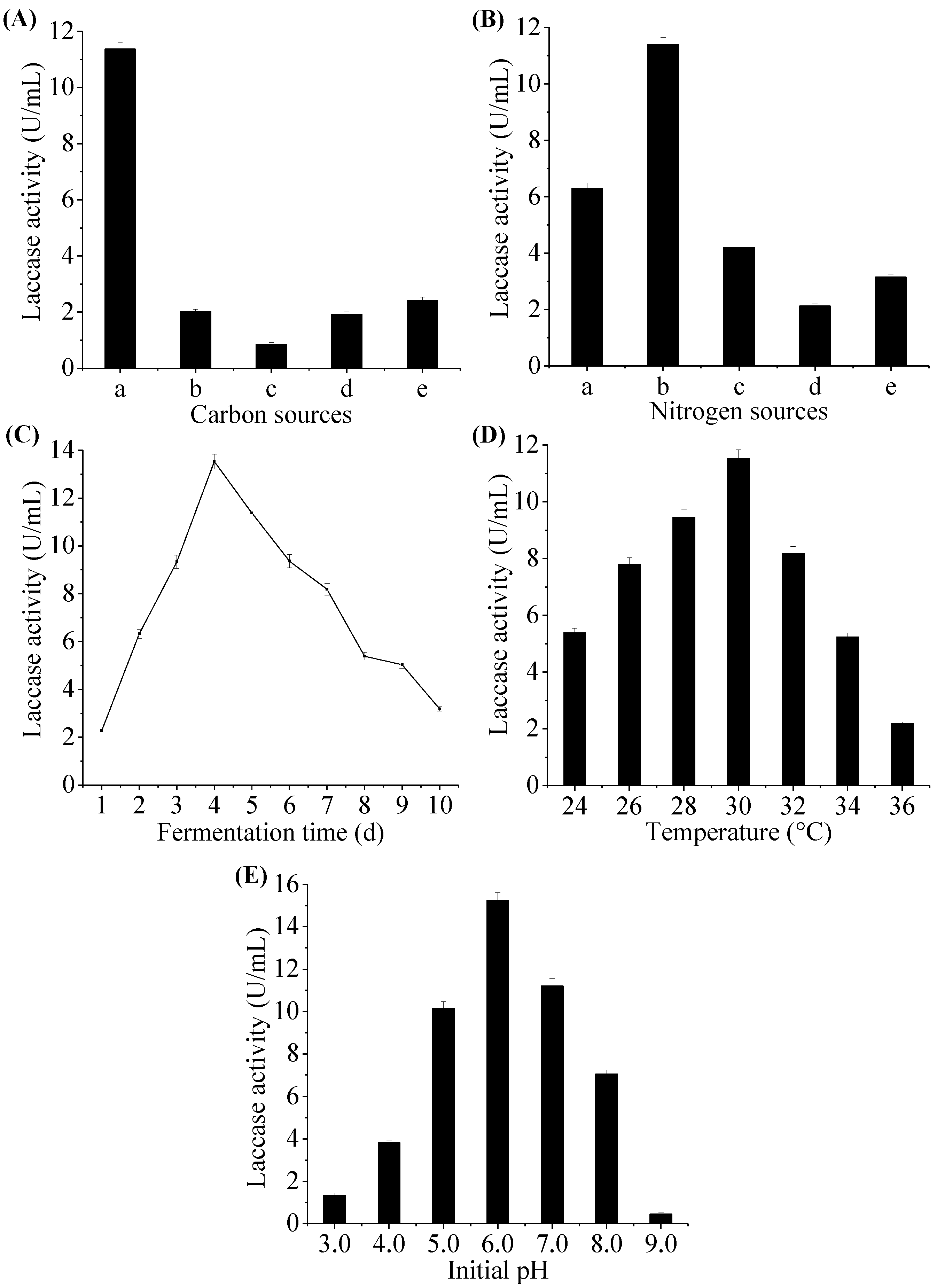
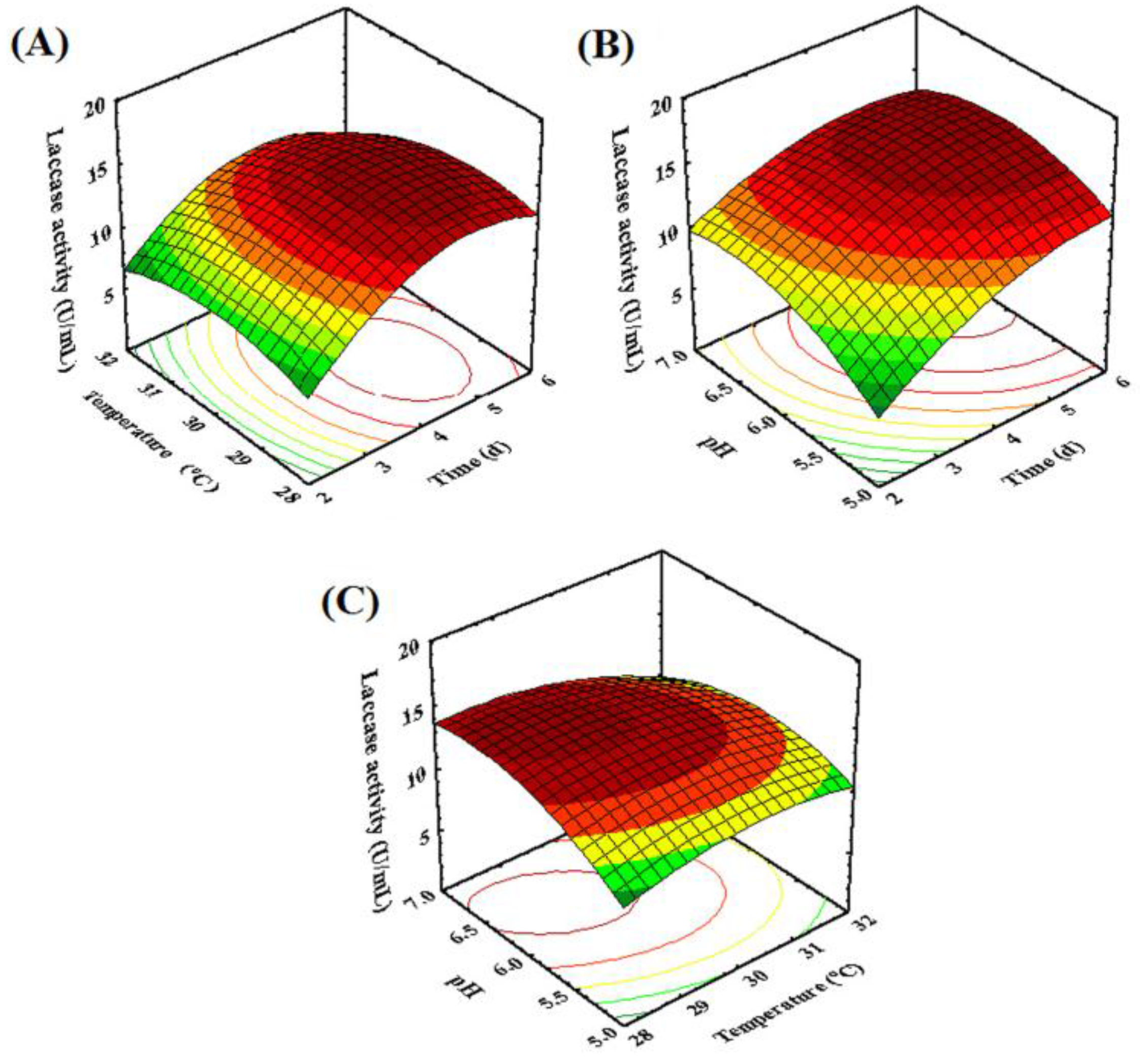
2.3. Optimization of Fermentation Conditions for Laccase Production by Fungal Endophyte MD-R-16
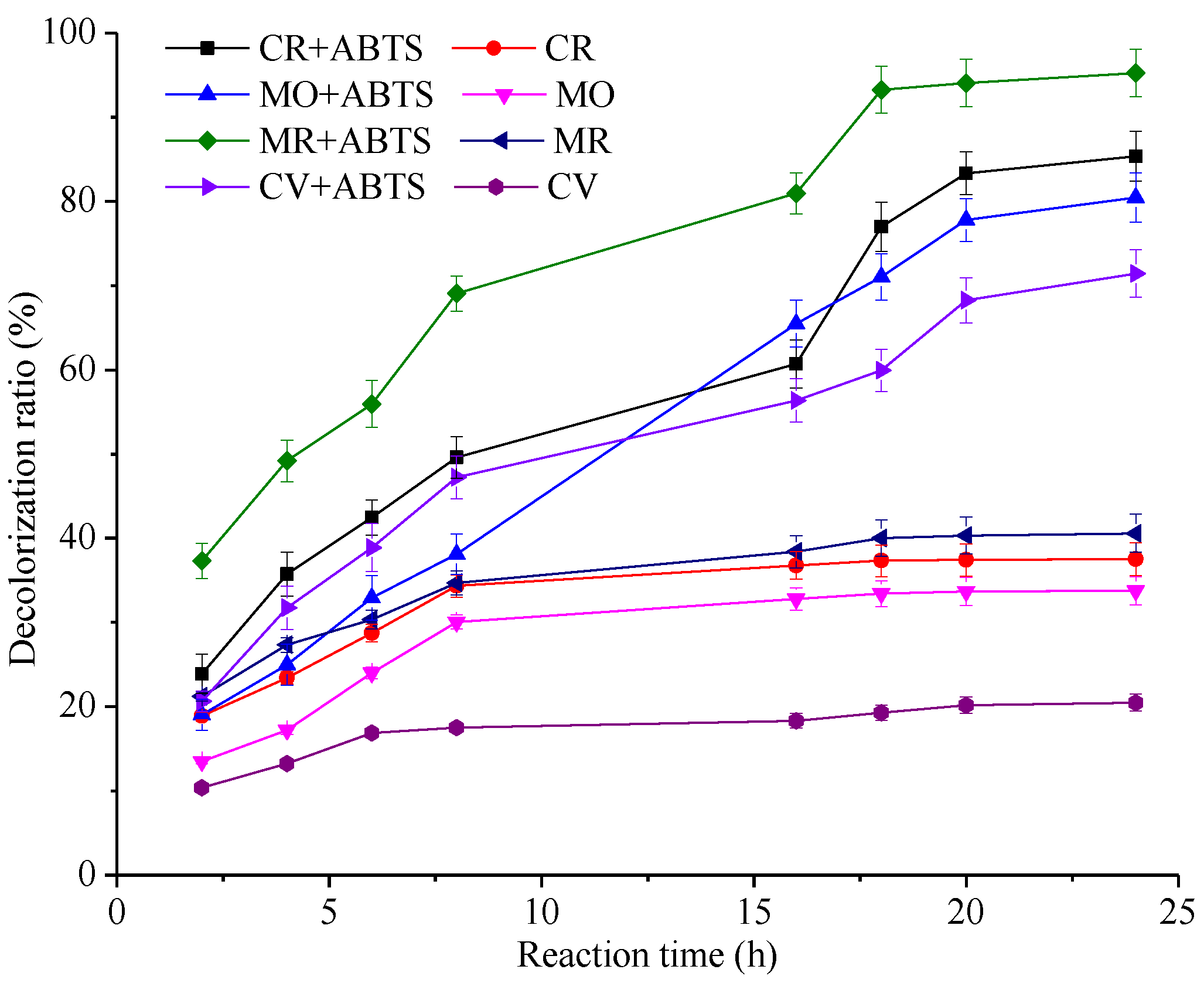
2.4. Dye Decolorization Test
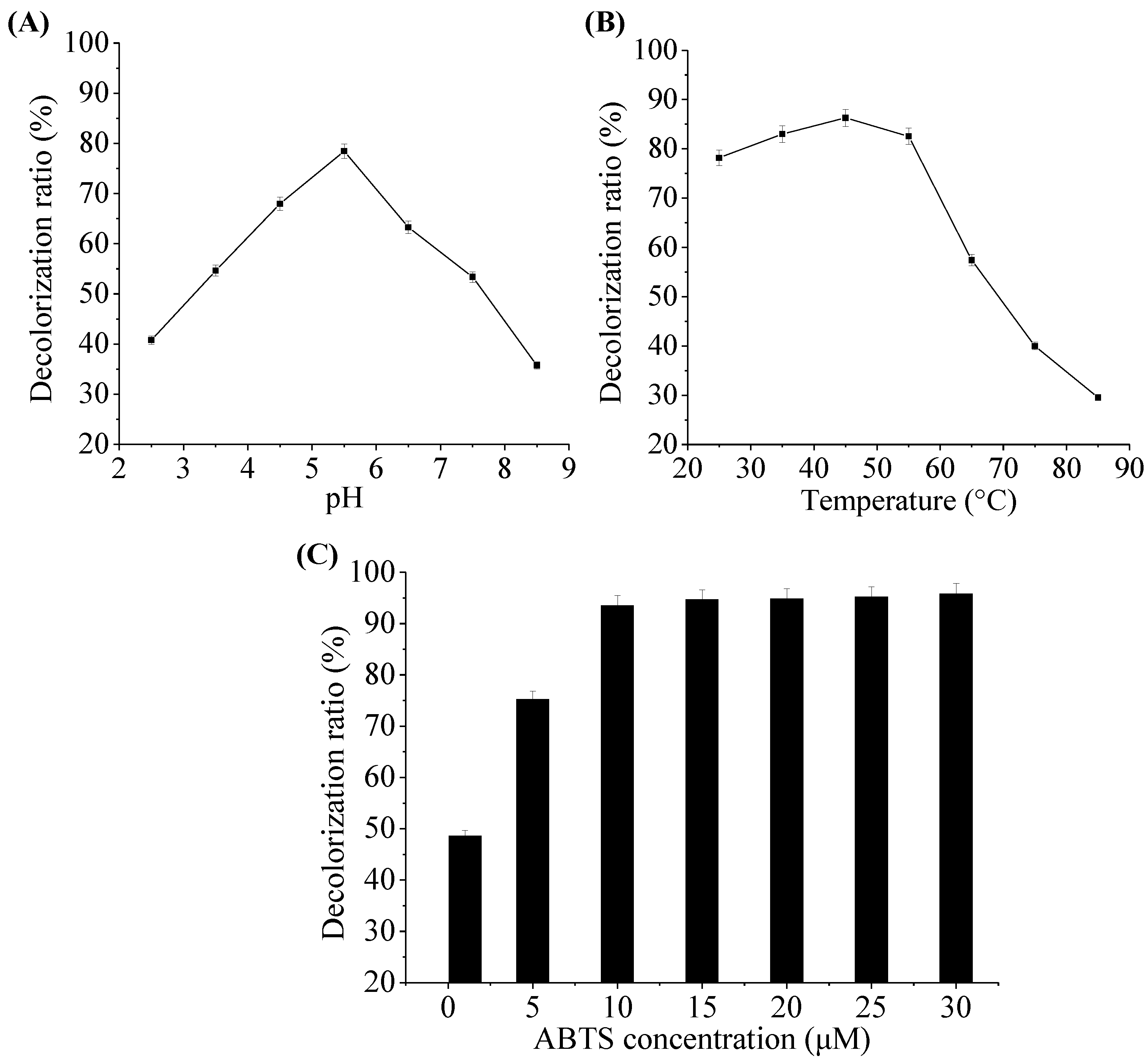
2.5. Effect of pH, Temperature and ABTS Concentration on Decolorization Ability of Laccase
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Isolation and Culture of the Fungal Endophyte
3.3. Screening of Laccase-Producing Fungal Endophyte
3.4. Identification of the Selected Fungal Endophyte by Morphological Properties and rDNA Sequence Analysis
3.5. Laccase Production and Enzyme Preparation
3.6. Determination of the Enzyme Activity
3.7. Experimental Design
3.8. Dye Decolorization
3.9. Statistical Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roriz, M.S.; Osma, J.F.; Teixeira, J.A.; Couto, S.R. Application of response surface methodological approach to optimise Reactive Black 5 decoloration by crude laccase from Trametes pubescens. J. Hazard. Mater. 2009, 169, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, S.; Ulaganathan, P.; Sivasubramanian, S.; Esakkimuthu, S.; Krishnaswamy, S.; Subramaniam, S. Trichoderma asperellum laccase mediated crystal violet degradation–optimization of experimental conditions and characterization. J. Environ. Chem. Eng. 2017, 5, 222–231. [Google Scholar] [CrossRef]
- Zeng, X.K.; Cai, Y.J.; Liao, X.R.; Zeng, X.L.; Li, W.X.; Zhang, D.B. Decolorization of synthetic dyes by crude laccase from a newly isolated Trametes trogii strain cultivated on solid agro-industrial residue. J. Hazard. Mater. 2011, 187, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Govindwar, S.P.; Kurade, M.B.; Tamboli, D.P.; Kabra, A.N.; Kim, P.J.; Waghmode, T.R. Decolorization and degradation of xenobiotic azo dye Reactive Yellow-84A and textile effluent by Galactomyces geotrichum. Chemosphere 2014, 109, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, S.; Tahmasbi, H.; Mogharabi, M.; Firuzyar, S.; Ameri, A.; Khoshayand, M.R.; Faramarzi, M.A. Efficient decolorization and detoxification of reactive orange 7 using laccase isolated from Paraconiothyrium variabile, kinetics and energetics. J. Taiwan Inst. Chem. Eng. 2015, 56, 113–121. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B.K. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, R.; Yuan, P.; Yang, Y.; Zhang, S.; Ma, F.; Zhang, X. Induction of laccase by metal ions and aromatic compounds in Pleurotus ostreatus HAUCC 162 and decolorization of different synthetic dyes by the extracellular laccase. Biochem. Eng. J. 2017, 117, 62–72. [Google Scholar] [CrossRef]
- Heidary, M.; Khoobi, M.; Ghasemi, S.; Habibi, Z.; Faramarzi, M.A. Synthesis of quinazolinones from alcohols via laccase-mediated tandem oxidation. Adv. Synth. Catal. 2014, 356, 1789–1794. [Google Scholar] [CrossRef]
- Nyanhongo, G.S.; Gomes, J.; Gübitz, G.; Zvauya, R.; Read, J.S.; Steiner, W. Production of laccase by a newly isolated strain of Trametes modesta. Bioresour. Technol. 2002, 84, 259–263. [Google Scholar] [CrossRef]
- Campos, P.A.; Levin, L.N.; Wirth, S.A. Heterologous production, characterization and dye decolorization ability of a novel thermostable laccase isoenzyme from Trametes trogii BAFC 463. Process Biochem. 2016, 51, 895–903. [Google Scholar] [CrossRef]
- Eldridge, H.C.; Milliken, A.; Farmer, C.; Hampton, A.; Wendland, N.; Coward, L.; Gregory, D.J.; Johnson, C.M. Efficient remediation of 17α-ethinylestradiol by Lentinula edodes (shiitake) laccase. Biocatal. Agric. Biotechnol. 2017, 10, 64–68. [Google Scholar] [CrossRef]
- Skoronski, E.; Souza, D.H.; Ely, C.; Broilo, F.; Fernandes, M.; Fúrigo, A.; Ghislandi, M.G. Immobilization of laccase from Aspergillus oryzae on graphene nanosheets. Int. J. Biol. Macromol. 2017, 99, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Basak, B.; Dutta, S.; Bhunia, B.; Dey, A. Decolorization and biodegradation of congo red dye by a novel white rot fungus Alternaria alternata CMERI F6. Bioresour. Technol. 2013, 147, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Pandey, V.P.; Dwivedi, U.N. Purification of a thermostable laccase from Leucaena leucocephala using a copper alginate entrapment approach and the application of the laccase in dye decolorization. Process Biochem. 2014, 49, 1196–1204. [Google Scholar] [CrossRef]
- Pan, K.; Zhao, N.N.; Yin, Q.; Zhang, T.W.; Xu, X.L.; Fang, W.; Hong, Y.Z.; Fang, Z.M.; Xiao, Y.Z. Induction of a laccase Lcc9 from Coprinopsis cinerea by fungal coculture and its application on indigo dye decolorization. Bioresour. Technol. 2014, 162, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Saroj, S.; Kumar, K.; Pareek, N.; Prasad, R.; Singh, R.P. Biodegradation of azo dyes Acid Red 183, Direct Blue 15 and Direct Red 75 by the isolate Penicillium oxalicum SAR-3. Chemosphere 2014, 107, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Ting, A.S.Y. Biodecolorization and biodegradation potential of recalcitrant triphenylmethane dyes by Coriolopsis sp. isolated from compost. J. Environ. Manag. 2015, 150, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Bhattacharya, S.; Panchanan, G.; Navya, B.S.; Nambiar, P. Production, characterization and Congo red dye decolorizing efficiency of a laccase from Pleurotus ostreatus MTCC 142 cultivated on co-substrates of paddy straw and corn husk. J. Genet. Eng. Biotechnol. 2016, 14, 281–288. [Google Scholar] [CrossRef]
- Riva, S. Laccases: Blue enzymes for green chemistry. Trends Biotechnol. 2006, 24, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhang, X.; Cui, D.Z.; Zhao, M. Characterisation of a novel white laccase from the deuteromycete fungus Myrothecium verrucaria NF-05 and its decolourisation of dyes. PLoS ONE 2012, 7, e38817. [Google Scholar] [CrossRef] [PubMed]
- EI Monssef, R.A.; Hassan, E.A.; Ramadan, E.M. Production of laccase enzyme for their potential application to decolorize fungal pigments on aging paper and parchment. Ann. Agric. Sci. 2016, 61, 145–154. [Google Scholar]
- Wu, Y.R.; Nian, D.L. Production optimization and molecular structure characterization of a newly isolated novel laccase from Fusarium solani MAS2, an anthracene-degrading fungus. Int. Biodeterior. Biodegrad. 2014, 86, 382–389. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Zhang, D.X.; Hua, Z.Z.; Li, J.H.; Du, G.C.; Chen, J. A newly isolated Paecilomyces sp. WSH-L07 for laccase production: Isolation, identification, and production enhancement by complex inducement. J. Ind. Microbiol. Biotechnol. 2009, 36, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Forootanfar, H.; Faramarzi, M.A.; Shahverdi, A.R.; Yazdi, M.T. Purification and biochemical characterization of extracellular laccase from the ascomycete Paraconiothyrium variabile. Bioresour. Technol. 2011, 102, 1808–1814. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tang, B.; Liu, Y.; Chen, A.; Tang, W.; Wei, S. High-level production and characterization of laccase from a newly isolated fungus Trametes sp. LS-10C. Biocatal. Agric. Biotechnol. 2016, 8, 278–285. [Google Scholar] [CrossRef]
- Adak, A.; Tiwari, R.; Singh, S.; Sharma, S.; Nain, L. Laccase production by a novel white-rot fungus Pseudolagarobasidium acaciicola LA 1 through solid-state fermentation of Parthenium biomass and its application in dyes decolorization. Waste Biomass Valor. 2016, 7, 1427–1435. [Google Scholar] [CrossRef]
- Wu, W.P. Studies on hyphomycetes in Hebei ΙΙΙ. species of the genus Myrothecium Tode: Fr. on plants. J. Hebei Acad. Sci. 1991, 8, 69–74. [Google Scholar]
- Kuhls, K.; Lieckfeldt, E.; Samuels, G.J.; Meyer, W.; Kubicek, C.P.; Börner, T. Revision of Trichoderma sect. longibrachiatum including related teleomorphs based on analysis of ribosomal DNA internal transcribed spacer sequences. Mycologia 1997, 89, 442–460. [Google Scholar]
- Murao, S.; Tanaka, N. Isolation and identification of a microorganism producing bilirubin oxidase. Agric. Biol. Chem. 1982, 46, 2031–2034. [Google Scholar]
- Sulistyaningdyah, W.T.; Ogawa, J.; Tanaka, H.; Maeda, C.; Shimizu, S. Characterization of alkaliphilic laccase activity in the culture supernatant of Myrothecium verrucaria 24G-4 in comparison with bilirubin oxidase. FEMS Microbiol. Lett. 2004, 230, 209–214. [Google Scholar] [CrossRef]
- Gao, D.; Fan, X.; Zhao, D. Isolation of a deuteromycete fungus Myrothecium verrucaria NF-08 and its laccase production. Sci. Silv. Sin. 2015, 51, 80–87. [Google Scholar]
- Hao, J.; Song, F.; Huang, F.; Yang, C.; Zhang, Z.; Zheng, Y.; Tian, X. Production of laccase by a newly isolated deuteromycete fungus Pestalotiopsis sp. and its decolorization of azo dye. J. Ind. Microbiol. Biotechnol. 2007, 34, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Goel, G.; Sud, A.; Chauhan, R.S. A novel laccase from newly isolated Cotylidia pannosa and its application in decolorization of synthetic dyes. Biocatal. Agric. Biotechnol. 2015, 4, 661–666. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Savitha, S.; Swaminathan, K.; Lin, F. Production, purification and characterization of mid-redox potential laccase from a newly isolated Trichoderma harzianum WL1. Process Biochem. 2008, 43, 736–742. [Google Scholar] [CrossRef]
- Bakkiyaraj, S.; Aravindan, R.; Arrivukkarasan, S.; Viruthagiri, T. Enhanced laccase production by Trametes hirusta using wheat bran under submerged fermentation. Int. J. Chem. Technol. Res. 2013, 5, 1224–1238. [Google Scholar]
- Afreen, S.; Anwer, R.; Singh, R.K.; Fatma, T. Extracellular laccase production and its optimization from Arthrospira maxima catalyzed decolorization of synthetic dyes. Saudi J. Biol. Sci 2016, in press. [Google Scholar] [CrossRef]
- Pointing, S.B.; Jones, E.B.G.; Vrijmoed, L.L.P. Optimization of laccase production by Pycnoporus sanguineus in submerged liquid culture. Mycologia 2000, 92, 139–144. [Google Scholar] [CrossRef]
- Mansor, A.F.; Mohidem, N.A.; Wan Mohd Zawawi, W.N.I.; Othman, N.S.; Endud, S.; Mat, H. The optimization of synthesis conditions for laccase entrapment in mesoporous silica microparticles by response surface methodology. Microporous Mesoporous Mater. 2016, 220, 308–314. [Google Scholar] [CrossRef]
- Kluczek-Turpeinen, B.; Tuomela, M.; Hatakka, A.; Hofrichter, M. Lignin degradation in a compost environment by the deuteromycete Paecilomyces inflatus. Appl. Microbiol. Biotechnol. 2003, 61, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Tavares, A.P.M.; Coelho, M.A.Z.; Agapito, M.S.M.; Coutinho, J.A.P. Optimization and modeling of laccase production by Trametes versicolor, in a bioreactor using statistical experimental design. Appl. Biochem. Biotechnol. 2006, 134, 233–248. [Google Scholar] [CrossRef]
- Vikineswary, S.; Abdullah, N.; Renuvathani, M.; Sekaran, M.; Pandey, A.; Jones, E.B.G. Productivity of laccase in solid substrate fermentation of selected agro-residues by Pycnoporus sanguineus. Bioresour. Technol. 2006, 97, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, R.; Rajendran, R.; Balakumar, C.; Tamilvendan, M. Isolation, screening and optimization of production medium for thermostable laccase production from Ganoderma sp. Int. J. Eng. Sci. Technol. 2010, 2, 7133–7141. [Google Scholar]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal laccases and their applications in bioremediation. Enzyme Res. 2014, 2014, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cañas, A.I.; Camarero, S. Laccases and their natural mediators: Biotechnological tools for sustainable eco-friendly processes. Biotechnol. Adv. 2010, 28, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.; Yu, J. Laccase-catalyzed decolorization of synthetic dyes. Appl. Biochem. Biotechnol. 1999, 33, 3512–3520. [Google Scholar] [CrossRef]
- Forootanfar, H.; Movahednia, M.M.; Yaghmaei, S.; Tabatabaei-Sameni, M.; Rastegar, H.; Sadighi, A.; Faramarzi, M.A. Removal of chlorophenolic derivatives by soil isolated ascomycete of Paraconiothyrium variabile and studying the role of its extracellular laccase. J. Hazard. Mater. 2012, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Kim, G.Y.; Yeon, K.H.; Cho, S.H.; Woo, J.J.; Moon, S.H. Degradation of azo dye by an electroenzymatic method using horseradish peroxidase immobilized on porous support. Korean J. Chem. Eng. 2007, 24, 72–78. [Google Scholar] [CrossRef]
- Zhao, J.; Li, C.; Wang, W.; Zhao, C.; Luo, M.; Mu, F.; Fu, Y.; Zu, Y.; Yao, M. Hypocrea lixii, novel endophytic fungi producing anticancer agent cajanol, isolated from pigeon pea (Cajanus cajan [L] Millsp.). J. Appl. Microbiol. 2013, 115, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Kiiskinen, L.L.; Rättö, M.; Kruus, K. Screening for novel laccase-producing microbes. J. Appl. Microbiol. 2004, 97, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Z.; Wang, G.Y. Diversity of fungal isolates from three Hawaiian marine sponges. Microbiol. Res. 2009, 164, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhao, M.; Zhang, B.B.; Yu, S.Y.; Bian, X.J.; Wang, W.; Wang, Y. Purification and characterization of laccase from Pycnoporus sanguineus and decolorization of an anthraquinone dye by the enzyme. Appl. Microbiol. Biotechnol. 2007, 74, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Runs | Factors | Laccase Activity (U/mL) | |||
|---|---|---|---|---|---|
| X1 (Time, d) | X2 (Temperature, °C) | X3 (pH) | Exp.a | Pred.b | |
| 1 | −1 (2) | −1 (28) | −1 (5) | 5.63 | 5.46 |
| 2 | 1 (6) | −1 (28) | −1 (5) | 11.95 | 11.88 |
| 3 | −1 (2) | 1 (32) | −1 (5) | 4.01 | 3.84 |
| 4 | 1 (6) | 1 (32) | −1 (5) | 8.03 | 7.86 |
| 5 | −1 (2) | −1 (28) | 1 (7) | 6.44 | 6.30 |
| 6 | 1 (6) | −1 (28) | 1 (7) | 11.51 | 11.36 |
| 7 | −1 (2) | 1 (32) | 1 (7) | 7.59 | 7.32 |
| 8 | 1 (6) | 1 (32) | 1 (7) | 10.12 | 9.98 |
| 9 | −1.68 (0.64) | 0 (30) | 0 (6) | 1.82 | 2.12 |
| 10 | 1.68 (7.36) | 0 (30) | 0 (6) | 9.55 | 9.75 |
| 11 | 0 (4) | −1.68 (26.64) | 0 (6) | 11.68 | 11.88 |
| 12 | 0 (4) | 1.68 (33.16) | 0 (6) | 9.05 | 9.36 |
| 13 | 0 (4) | 0 (30) | −1.68 (4.32) | 7.43 | 7.66 |
| 14 | 0 (4) | 0 (30) | 1.68 (7.68) | 9.84 | 10.14 |
| 15 | 0 (4) | 0 (30) | 0 (6) | 16.28 | 16.24 |
| 16 | 0 (4) | 0 (30) | 0 (6) | 16.53 | 16.24 |
| 17 | 0 (4) | 0 (30) | 0 (6) | 16.49 | 16.24 |
| 18 | 0 (4) | 0 (30) | 0 (6) | 15.82 | 16.24 |
| 19 | 0 (4) | 0 (30) | 0 (6) | 15.97 | 16.24 |
| 20 | 0 (4) | 0 (30) | 0 (6) | 16.41 | 16.24 |
| Source | Sum of Squares | DF | Mean Square | F Value | p-value Prob > F | Significance |
|---|---|---|---|---|---|---|
| Model | 387.31 | 9 | 43.03 | 422.29 | <0.0001 | Significant |
| X1 | 70.10 | 1 | 70.10 | 688.17 | <0.0001 | |
| X2 | 7.62 | 1 | 7.62 | 74.84 | <0.0001 | |
| X3 | 7.46 | 1 | 7.46 | 73.23 | <0.0001 | |
| X1X2 | 2.93 | 1 | 2.93 | 28.75 | 0.0003 | |
| X1X3 | 0.94 | 1 | 0.94 | 9.21 | 0.0126 | |
| X2X3 | 3.51 | 1 | 3.51 | 34.47 | 0.0002 | |
| X12 | 191.68 | 1 | 191.68 | 1881.84 | <0.0001 | |
| X22 | 57.21 | 1 | 57.21 | 561.64 | <0.0001 | |
| X32 | 97.72 | 1 | 97.72 | 959.40 | <0.0001 | |
| Residual | 1.02 | 10 | 0.10 | |||
| Lack of Fit | 0.59 | 5 | 0.12 | 1.39 | 0.3726 | Not significant |
| R2 | 0.9974 | |||||
| Adjusted R2 | 0.9950 |
| Dye | Classification | Molecular Structure | Molecular Weight | λmax (nm) |
|---|---|---|---|---|
| Crystal Violet | Triphenylmethane |  | 407.98 | 577 |
| Congo Red | Azo |  | 696.66 | 497 |
| Methyl Orange | Azo |  | 327.33 | 463 |
| Methyl Red | Azo |  | 269.30 | 410 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Guo, N.; Niu, L.-L.; Wang, Q.-F.; Zang, Y.-P.; Zu, Y.-G.; Fu, Y.-J. Production of Laccase by a New Myrothecium verrucaria MD-R-16 Isolated from Pigeon Pea [Cajanus cajan (L.) Millsp.] and its Application on Dye Decolorization. Molecules 2017, 22, 673. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040673
Sun J, Guo N, Niu L-L, Wang Q-F, Zang Y-P, Zu Y-G, Fu Y-J. Production of Laccase by a New Myrothecium verrucaria MD-R-16 Isolated from Pigeon Pea [Cajanus cajan (L.) Millsp.] and its Application on Dye Decolorization. Molecules. 2017; 22(4):673. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040673
Chicago/Turabian StyleSun, Jiao, Na Guo, Li-Li Niu, Qing-Fang Wang, Yu-Ping Zang, Yuan-Gang Zu, and Yu-Jie Fu. 2017. "Production of Laccase by a New Myrothecium verrucaria MD-R-16 Isolated from Pigeon Pea [Cajanus cajan (L.) Millsp.] and its Application on Dye Decolorization" Molecules 22, no. 4: 673. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040673