Cloning and Functional Characterization of Two 4-Coumarate: CoA Ligase Genes from Selaginella moellendorffii

Abstract
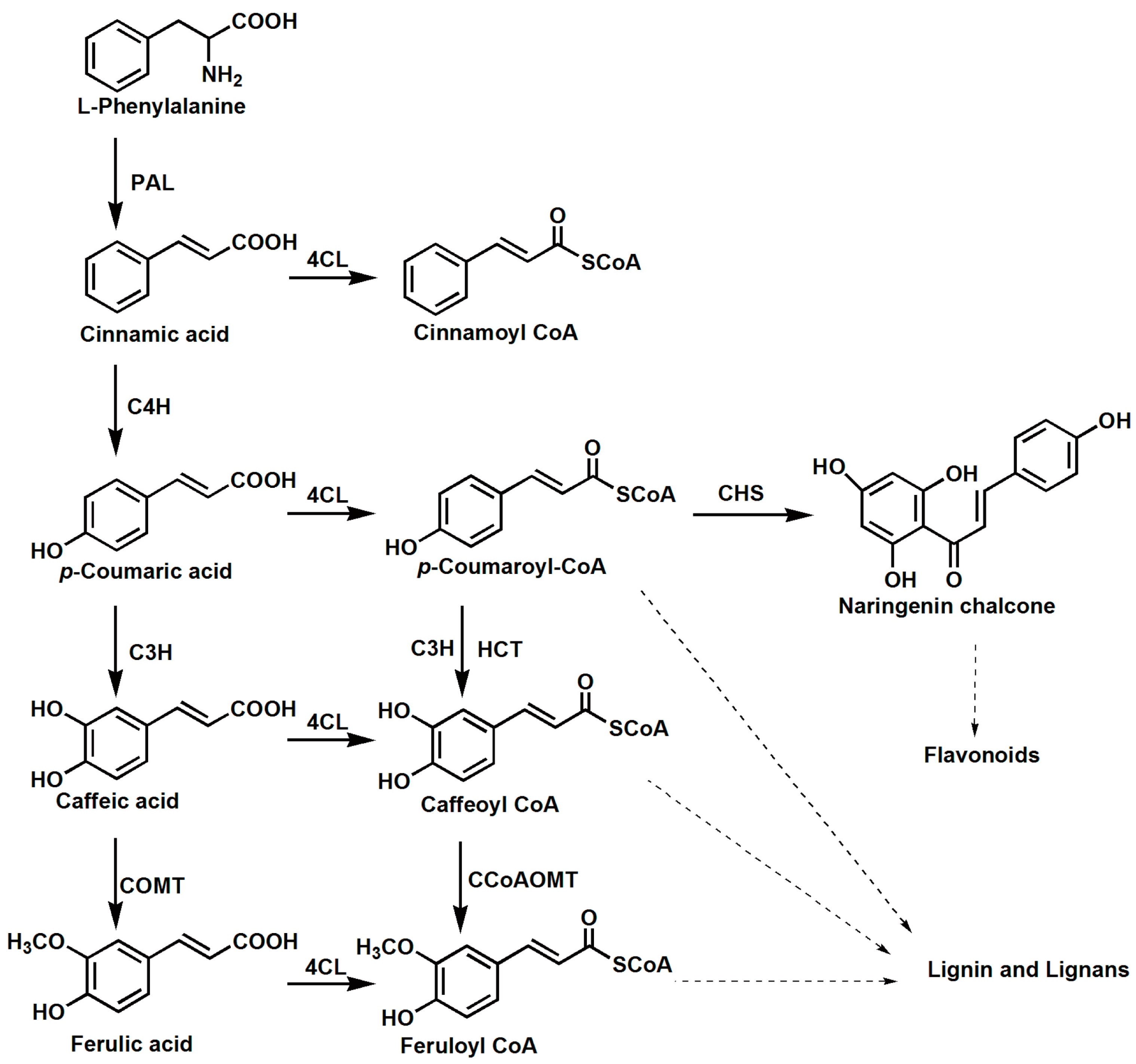
:1. Introduction
2. Results
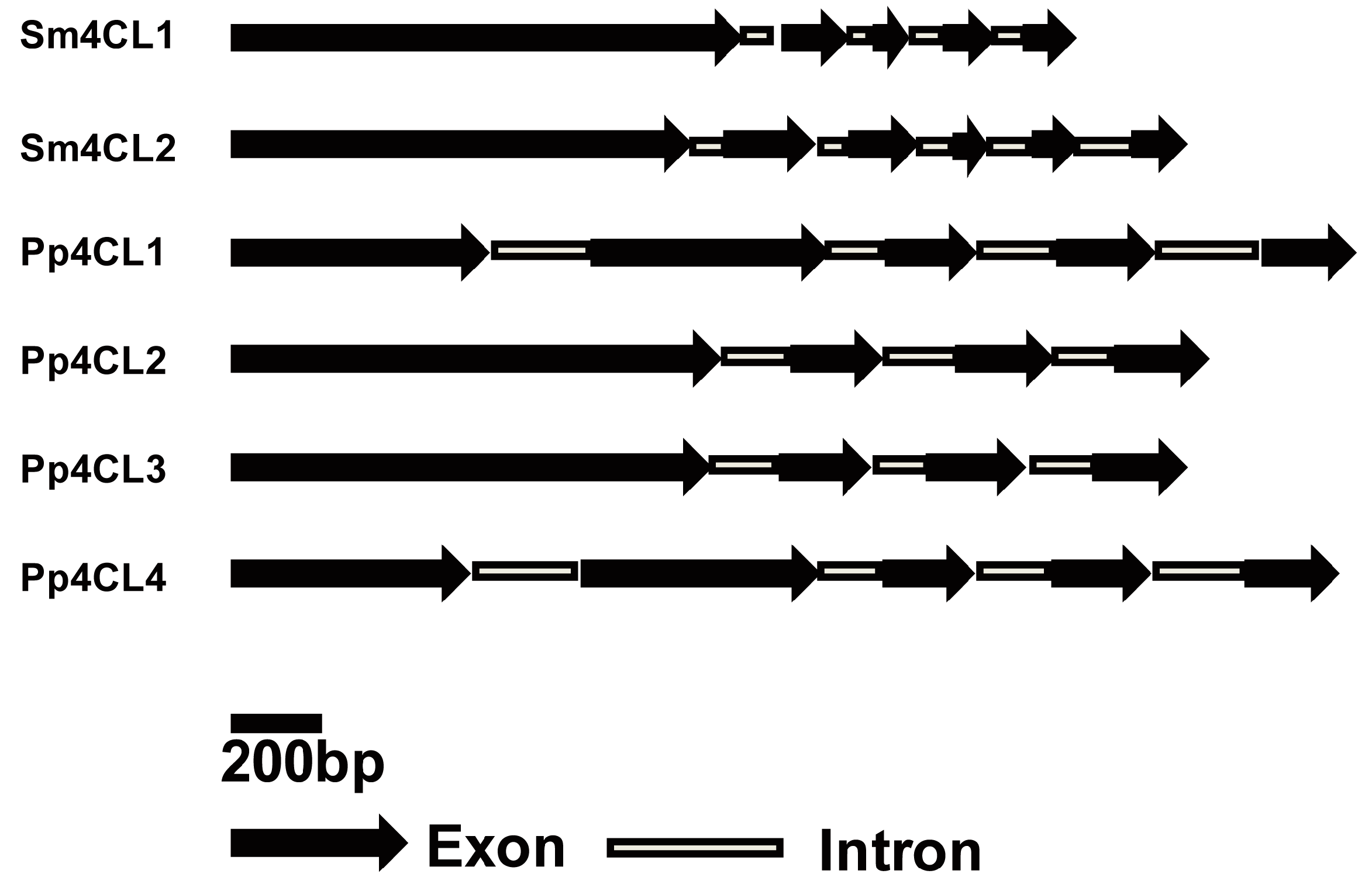
2.1. Identification of 4CL Genes from S. moellendorffii
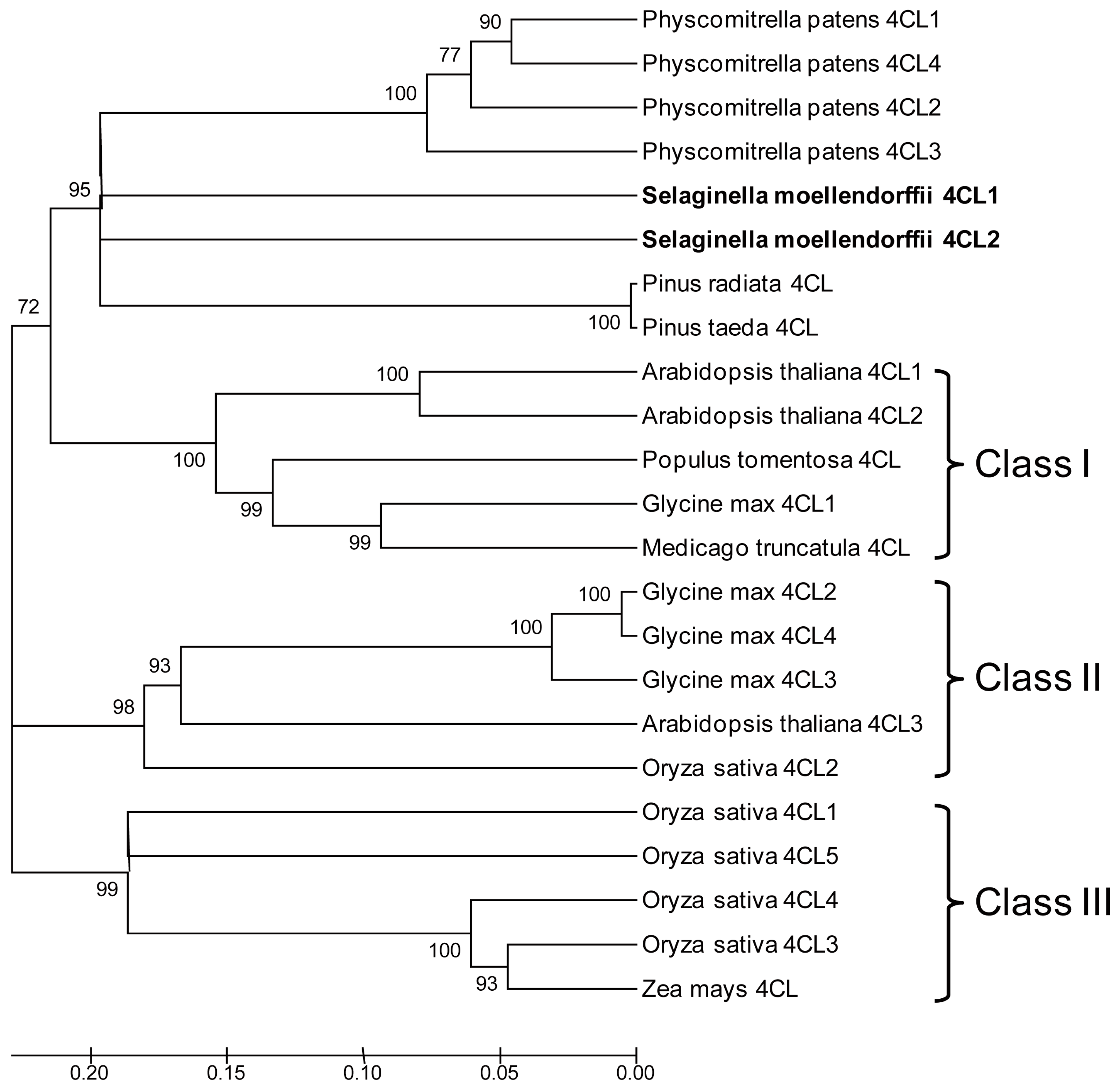
2.2. Protein Sequence and Phylogenetic Analysis of Sm4CLs
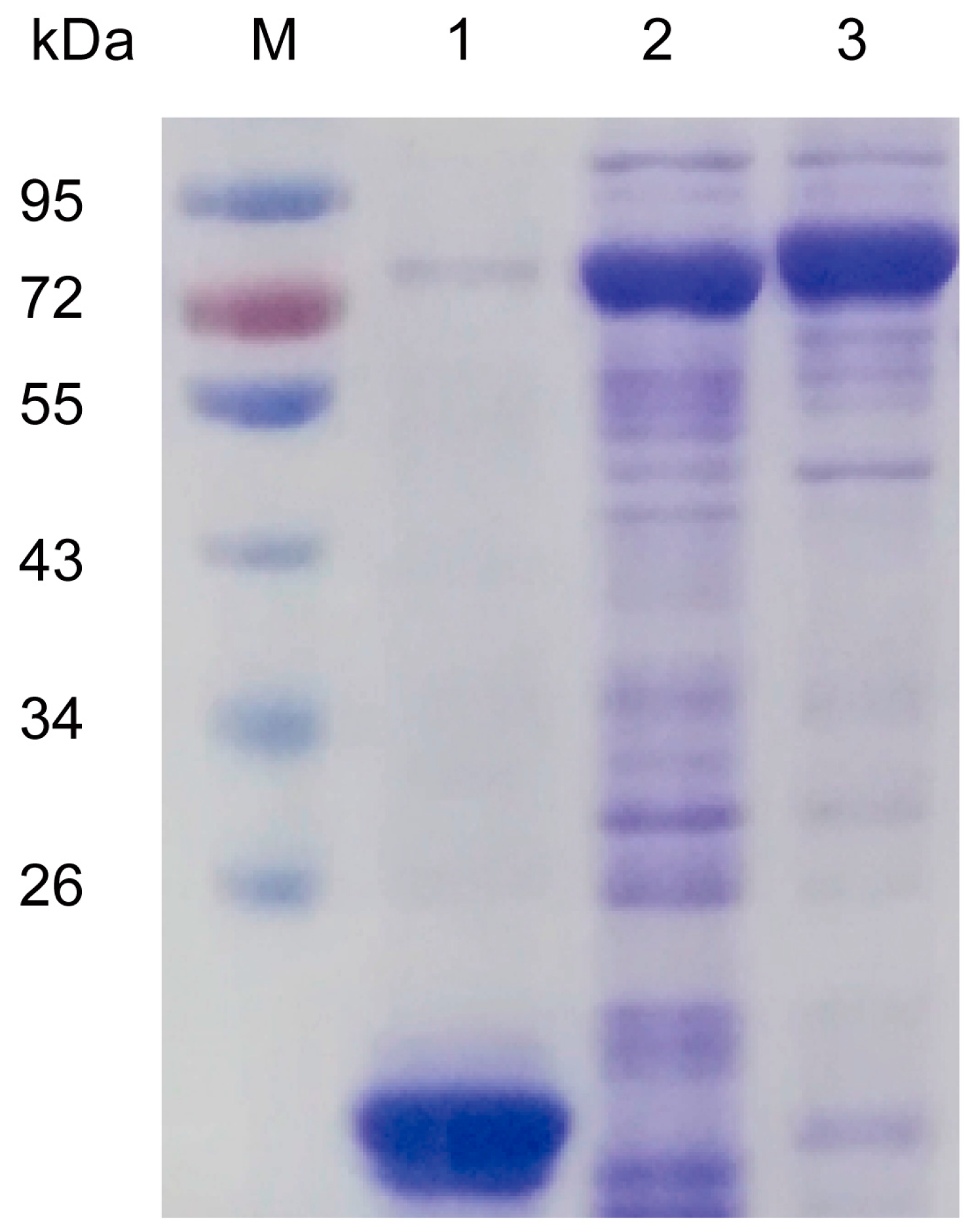
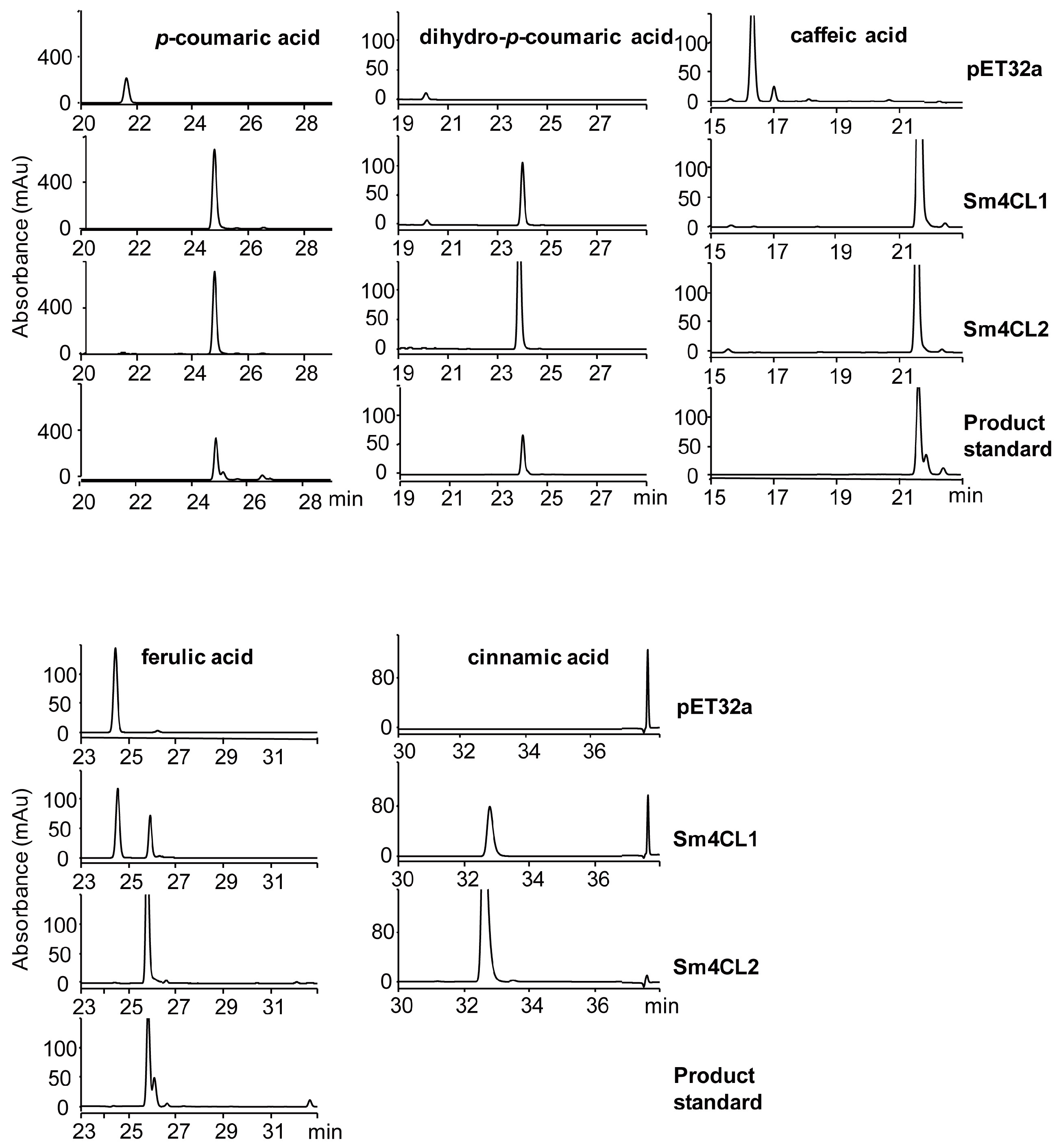
2.3. Biochemical Characterization of Recombinant Sm4CL In Vitro
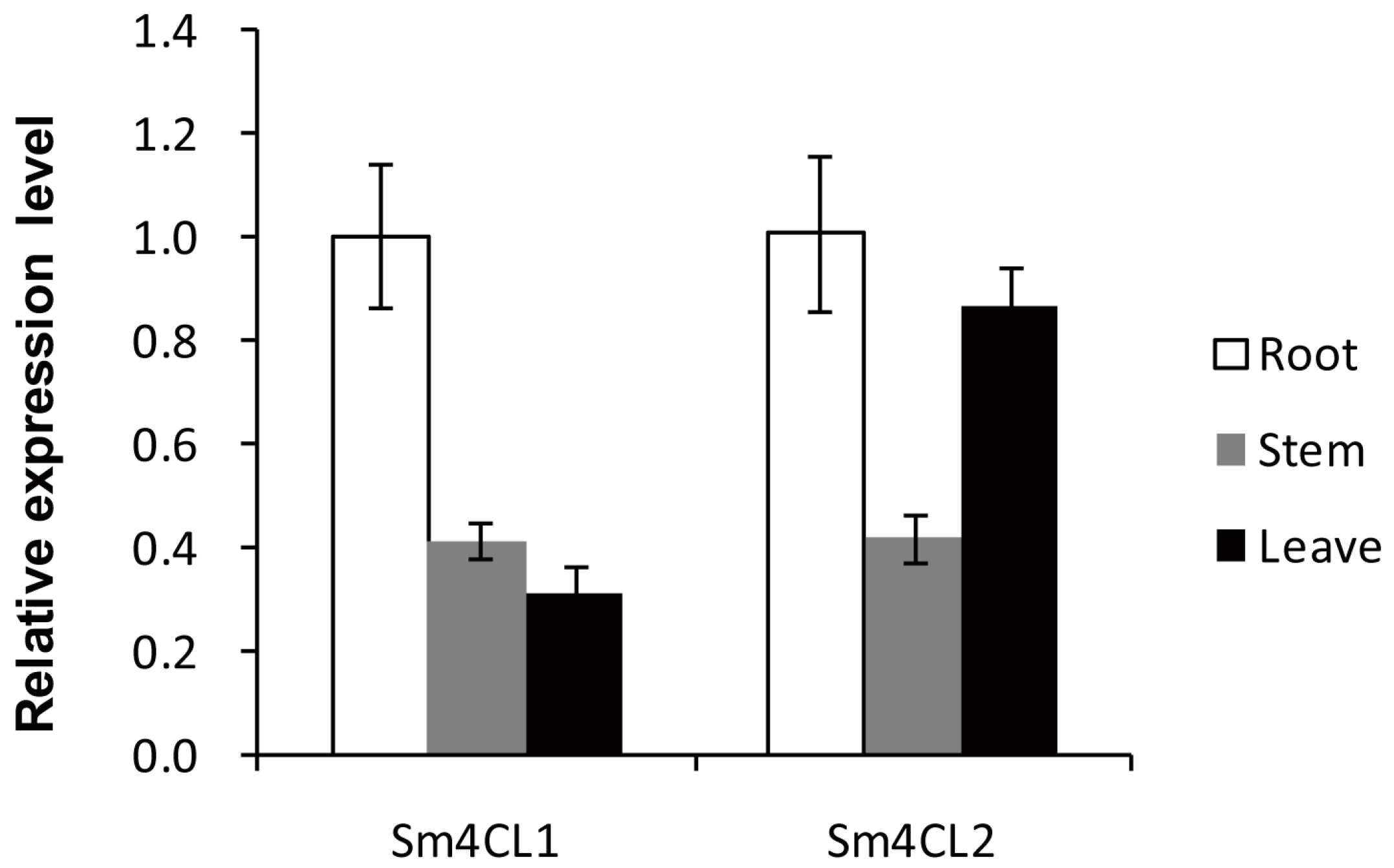
2.4. Determination of Gene Expression Patterns in Tissues
3. Discussion
4. Materials and Methods
4.1. Plant Material, Nucleic Acid Extraction, and Reagents
4.2. DNA Isolation and Sequence Analysis
4.3. Recombinant Protein Expression and Purification
4.4. Enzyme Assays
4.5. Expression Profile Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence-a genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Vassão, D.G.; Kim, K.-W.; Davin, L.B.; Lewis, N.G. Lignans (neolignans) and allyl/propenyl phenols: Biogenesis, structural biology, and biological/human health considerations. In Comprehensive Natural Products II Chemistry and Biology; Mander, L., Lui, H.-W., Eds.; Elsevier: Oxford, UK, 2010; Volume 1, pp. 815–928. [Google Scholar]
- Hamberger, B.; Hahlbrock, K. The 4-coumarate:CoA ligase gene family in Arabidopsis thaliana comprises one rare, sinapate-activating and three commonly occurring isoenzymes. Proc. Natl. Acad. Sci. USA 2004, 101, 2209–2214. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Shen, J.; Li, L. Functional characterization of evolutionarily divergent 4-coumarate:coenzyme A ligases in rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Silber, M.V.; Meimberg, H.; Ebel, J. Identification of a 4-coumarate:CoA ligase gene family in the moss, physcomitrella patens. Phytochemistry 2008, 69, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Becker-Andre, M.; Schulze-Lefert, P.; Hahlbrock, K. Structural comparison, modes of expression, and putative cis-acting elements of the two 4-coumarate:CoA ligase genes in potato. J. Biol. Chem. 1991, 266, 8551–8559. [Google Scholar] [PubMed]
- Lozoya, E.; Hoffmann, H.; Douglas, C.; Schulz, W.; Scheel, D.; Hahlbrock, K. Primary structures and catalytic properties of isoenzymes encoded by the two 4-coumarate:CoA ligase genes in parsley. Eur. J. Biochem. 1988, 176, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Kawaoka, A.; Tsai, C.J.; Lung, J.; Osakabe, K.; Ebinuma, H.; Chiang, V.L. Compartmentalized expression of two structurally and functionally distinct 4-coumarate:CoA ligase genes in aspen (Populus tremuloides). Proc. Natl. Acad. Sci. USA 1998, 95, 5407–5412. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.A. Selaginella and 400 million years of separation. Annu. Rev. Plant Biol. 2009, 60, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.K.; Noel, J.P. Chemodiversity in Selaginella: A reference system for parallel and convergent metabolic evolution in terrestrial plants. Front. Plant Sci. 2013, 4, 119. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tanurdzic, M.; Luo, M.; Sisneros, N.; Kim, H.R.; Weng, J.K.; Kudrna, D.; Mueller, C.; Arumuganathan, K.; Carlson, J.; et al. Construction of a bacterial artificial chromosome library from the spikemoss Selaginella moellendorffii: A new resource for plant comparative genomics. BMC Plant Biol. 2005, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, J.A.; Nishiyama, T.; Hasebe, M.; Bowman, J.L.; Gribskov, M.; dePamphilis, C.; Albert, V.A.; Aono, N.; Aoyama, T.; Ambrose, B.A.; et al. The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science 2011, 332, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Stuible, H.P.; Buttner, D.; Ehlting, J.; Hahlbrock, K.; Kombrink, E. Mutational analysis of 4-coumarate:CoA ligase identifies functionally important amino acids and verifies its close relationship to other adenylate-forming enzymes. FEBS Lett. 2000, 467, 117–122. [Google Scholar] [CrossRef]
- Kajita, S.; Hishiyama, S.; Tomimura, Y.; Katayama, Y.; Omori, S. Structural characterization of modified lignin in transgenic tobacco plants in which the activity of 4-coumarate:coenzyme A ligase is depressed. Plant Physiol. 1997, 114, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Donaldson, L.; Kim, H.; Phillips, L.; Flint, H.; Steward, D.; Torr, K.; Koch, G.; Schmitt, U.; Ralph, J. Suppression of 4-coumarate-CoA ligase in the coniferous gymnosperm Pinus radiata. Plant Physiol. 2009, 149, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Ehlting, J.; Buttner, D.; Wang, Q.; Douglas, C.J.; Somssich, I.E.; Kombrink, E. Three 4-coumarate:coenzyme A ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Meyer, K.; Chapple, C.; Douglas, C.J. Antisense suppression of 4-coumarate:coenzyme A ligase activity in Arabidopsis leads to altered lignin subunit composition. Plant Cell 1997, 9, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Koutaniemi, S.; Warinowski, T.; Karkonen, A.; Alatalo, E.; Fossdal, C.G.; Saranpaa, P.; Laakso, T.; Fagerstedt, K.V.; Simola, L.K.; Paulin, L.; et al. Expression profiling of the lignin biosynthetic pathway in norway spruce using EST sequencing and real-time RT-PCR. Plant Mol. Biol. 2007, 65, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ellis, B.E. 4-coumarate:CoA ligase gene family in Rubus idaeus: cDNA structures, evolution, and expression. Plant Mol. Biol. 2003, 51, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Yu, J.; Cai, Y.X.; Zhu, P.P.; Liu, C.Y.; Zhao, A.C.; Lu, R.H.; Li, M.J.; Xu, F.X.; Yu, M.D. Characterization and functional analysis of 4-coumarate:CoA ligase genes in Mulberry. PLoS ONE 2016, 11, e0155814. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Kumar, R.; Chanotiya, C.S.; Shanker, K.; Gupta, M.M.; Nagegowda, D.A.; Shasany, A.K. 4-Coumarate:CoA ligase partitions metabolites for eugenol biosynthesis. Plant Cell Physiol. 2013, 54, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Yao, R.L.; Zhao, Y.C.; Xu, S.; Huang, C.L.; Luo, J.; Kong, L.Y. Cloning, functional characterization and site-directed mutagenesis of 4-coumarate:coenzyme A ligase (4CL) involved in coumarin biosynthesis in Peucedanum praeruptorum dunn. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Chen, H.Y.; Babst, B.A.; Nyamdari, B.; Hu, H.; Sykes, R.; Davis, M.F.; Harding, S.A.; Tsai, C.J. Ectopic expression of a loblolly pine class II 4-coumarate:CoA ligase alters soluble phenylpropanoid metabolism but not lignin biosynthesis in populus. Plant Cell Physiol. 2014, 55, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.B.; Li, C.F.; Li, J.; Zhang, Y.S. Molecular cloning and functional characterization of two divergent 4-coumarate:coenzyme A ligases from Kudzu (pueraria lobata). Biol. Pharm. Bull. 2014, 37, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Logan, K.J.; Thomas, B.A. Distribution of lignin derivatives in plants. New Phytol. 1985, 99, 571–585. [Google Scholar] [CrossRef]
- Jin, Z.F.; Matsumoto, Y.; Tange, T.; Akiyama, T.; Higuchi, M.; Ishii, T.; Iiyama, K. Proof of the presence of guaiacyl-syringyl lignin in Selaginella tamariscina. J. Wood Sci. 2005, 51, 424–426. [Google Scholar] [CrossRef]
- Weng, J.K.; Li, X.; Stout, J.; Chapple, C. Independent origins of syringyl lignin in vascular plants. Proc. Natl. Acad. Sci. USA 2008, 105, 7887–7892. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Choi, H.; An, G. Roles of lignin biosynthesis and regulatory genes in plant development. J. Integr. Plant Biol. 2015, 57, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.A.; Campbell, M.M. The genetic control of lignin deposition during plant growth and development. New Phytol. 2004, 164, 17–30. [Google Scholar] [CrossRef]
- Weng, J.K.; Akiyama, T.; Bonawitz, N.D.; Li, X.; Ralph, J.; Chapple, C. Convergent evolution of syringyl lignin biosynthesis via distinct pathways in the lycophyte Selaginella and flowering plants. Plant Cell 2010, 22, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.K.; Akiyama, T.; Ralph, J.; Chapple, C. Independent recruitment of an o-methyltransferase for syringyl lignin biosynthesis in Selaginella moellendorffii. Plant Cell 2011, 23, 2708–2724. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Gambino, G.; Perrone, I.; Gribaudo, I. A rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. PCA 2008, 19, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Beuerle, T.; Pichersky, E. Enzymatic synthesis and purification of aromatic coenzyme A esters. Anal. Biochem. 2002, 302, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.B.; Campbell, W.H. Facile enzymic synthesis of caffeoyl CoA. Phytochemistry 1997, 44, 605–608. [Google Scholar] [CrossRef]
- Obel, N.; Scheller, H.V. Enzymatic synthesis and purification of caffeoyl-CoA, p-coumaroyl-CoA, and feruloyl-CoA. Anal. Biochem. 2000, 286, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Phytozome v12.1: Home. Available online: https://phytozome.jgi.doe.gov/pz/portal.html (accessed on 12 February 2018).
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. Mega4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Luderitz, T.; Schatz, G.; Grisebach, H. Enzymic synthesis of lignin precursors. Purification and properties of 4-coumarate:CoA ligase from cambial sap of spruce (Picea abies L.). Eur. J. Biochem. 1982, 123, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Stockigt, J.; Zenk, M.H. Chemical syntheses and properties of hydroxycinnamoyl-coenzyme a derivatives. Z. Naturforsch. C 1975, 30, 352–358. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds p-Coumaroyl CoA, dihydro-p-coumaroyl CoA, caffeoyl CoA, and feruloyl CoA are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Specific Activity (nmol mg−1 min−1) | |
|---|---|---|
| Sm4CL1 | Sm4CL2 | |
| p-coumaric acid | 71.77 ± 0.31 | 557.5 ± 15.58 |
| caffeic acid | 16.92 ± 0.2 | 239.71 ± 5.6 |
| cinnamic acid | 2.29 ± 0.08 | 283.08 ± 5.22 |
| ferulic acid | 1.14 ± 0.00 | 494.83 ± 3.61 |
| dihydro-p-coumaric acid | 16.71 ± 0.45 | 271.77 ± 29.57 |
| sinapic acid | ND | ND |
| Enzyme | Substrate | Km (μM) | Vmax (nmol mg−1 min−1) | kcat (min−1) | kenz (M−1 min−1) |
|---|---|---|---|---|---|
| Sm4CL1 | p-coumaric acid | 11.89 ± 1.56 | 71.37 ± 2.58 | 4.09 ± 0.15 | 343,986.54 |
| caffeic acid | 10.87 ± 1.6 | 25.40 ± 0.94 | 1.46 ± 0.05 | 134,314.63 | |
| di-p-coumaric acid | 442.4 ± 53.74 | 43.73 ± 1.34 | 2.51 ± 0.08 | 5673.6 | |
| Sm4CL2 | p-coumaric acid | 19.67 ± 2.92 | 697.1 ± 32.06 | 42.91 ± 1.97 | 2,181,494.66 |
| caffeic acid | 18.96 ± 2.61 | 268.1 ± 9.74 | 16.50 ± 0.60 | 870,253.16 | |
| cinnamic acid | 126.9 ± 11.57 | 382 ± 9.31 | 23.52 ± 0.57 | 185,310.64 | |
| ferulic acid | 95.99 ± 13.39 | 525.2 ± 21.18 | 32.33 ± 1.3 | 336,819.59 | |
| di-p-coumaric acid | 220.1 ± 36.86 | 673.1 ± 23.19 | 41.44 ± 1.43 | 188,278.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.-Y.; Wang, P.-P.; Wu, Y.-F.; Cheng, A.-X.; Lou, H.-X. Cloning and Functional Characterization of Two 4-Coumarate: CoA Ligase Genes from Selaginella moellendorffii. Molecules 2018, 23, 595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030595
Liu X-Y, Wang P-P, Wu Y-F, Cheng A-X, Lou H-X. Cloning and Functional Characterization of Two 4-Coumarate: CoA Ligase Genes from Selaginella moellendorffii. Molecules. 2018; 23(3):595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030595
Chicago/Turabian StyleLiu, Xin-Yan, Ping-Ping Wang, Yi-Feng Wu, Ai-Xia Cheng, and Hong-Xiang Lou. 2018. "Cloning and Functional Characterization of Two 4-Coumarate: CoA Ligase Genes from Selaginella moellendorffii" Molecules 23, no. 3: 595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030595