1. Introduction
Docosahexaenoic acid (DHA) plays a significant role as a nutrient required by both invertebrates and vertebrates. Since DHA is indispensable for prenatal and postnatal development of visual and other perceptive functions, this compound is added to many infant dietary formulas [
1,
2]. Additionally, it has been observed that DHA is correlated with enhanced quality of life and reduced risk of cardiovascular and neurological diseases [
3].
Mammals lack the enzymes required to produce DHA de novo, so this substance must be obtained through the diet. A variety of marine fish species are currently the primary commercial sources of DHA. However, fish-derived DHA is associated with many shortcomings, including unstable quality, a fishy odor, environmental pollution, high processing costs and a tendency to oxidize [
4,
5]. Accordingly, efforts have been made to divert commercial DHA production towards the exploitation of oleaginous microorganisms that biosynthesize DHA, such as thraustochytrids and marine diatoms [
6].
Thraustochytrium, unicellular eukaryotic marine protists, have been isolated mainly from coastal marine environments [
7].
Thraustochytrium strains have attracted attention from researchers worldwide, since these can build dense biomass in culture and produce polyunsaturated fatty acids (PUFAs), especially those from the omega−3 series, such as DHA [
8]. Therefore, some
Thraustochytrium strains are considered to be prospective alternatives to marine fish oils as new sources of DHA for use in aquaculture products and food additives.
To optimize DHA production, several chemical parameters, including carbon and nitrogen sources at different concentrations, as well as physical parameters, including initial pH and inoculum age, must be considered [
9]. It was observed that thraustochytrids isolated from different environments also have variant DHA production levels [
10,
11]. The fermentation conditions must, furthermore, have a decisive role in regulating the DHA production of thraustochytrids [
12]. With the aim of high DHA productivity, a temperature shift regime resulted in a DHA content of 52% of total fatty acids. A decline in temperature from 30 °C to 20 °C yielded high levels of DHA, making 30 °C the optimal culture temperature for the highest biomass growth [
13]. Glucose and glycerol are used as a mixed carbon source for
Aurantiochytrium limacinum SR21, and DHA productivity was 15.24% higher than that obtained using glucose as a single carbon source [
14]. The continuous culture method was carried out to feed
A. limacinum SR21, whereby the yield of biomass was 11.78 g/L and DHA concentration reached 1.74 g/L [
15]. Response surface methodology (RSM) is known to be effective for the optimization of multiple factors for fermentation yields [
16]. The empirical model developed by RSM was adequate to ascertain the possible interactions between the studied parameters and the response of DHA production. For example, RSM was previously used in optimizing significant fermentation variables (glucose concentration, yeast extract addition, NaCl content, pH, and incubation time), leading to a maximum DHA concentration of 516 mg/L by
Schizochytrium sp. S31 [
17].
This work attempts to optimize the fermentation parameters in the cultivation of Thraustochytrium sp. so that it may enhance the DHA yield. The influence of fermentation parameters on the production of DHA were investigated through the Box-Behnken design of experiments.
3. Discussion
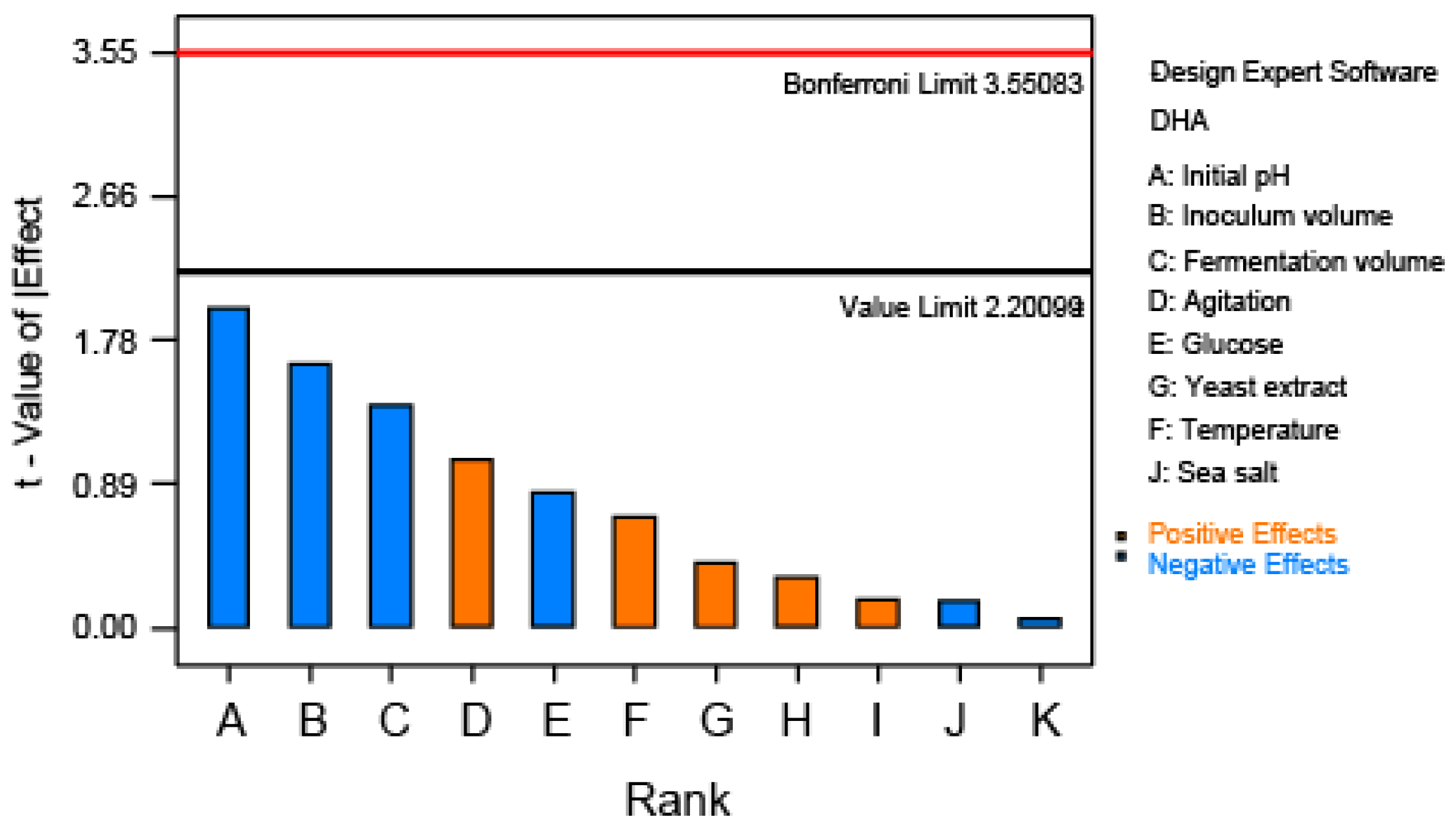
Response surface methodology is suitable for processes optimization for many industrial products, including paints and coatings, foods and beverages, and pharmaceuticals. Compared with one-factor-at-a-time experiments, statistically designed experiments are able to describe the effect of the interactions among the factors in linear and quadratic terms. In the present study, the optimization of DHA production by Thraustochytrium sp. ATCC 26185 was divided into two phases: a screening of main effects of the selected variables and a response optimization. Reduction in the initial number of variables was carried out through Plackett-Burman design and, among eight tested variables, the initial pH, inoculum volume, and fermentation volume were taken for BBD of RSM to assess their effects on DHA production by Thraustochytrium sp. ATCC 26185.
A large model
F-value indicates that most of the variation in the response can be explained by a given regression equation. The ANOVA result for DHA production by
Thraustochytrium sp. ATCC 26185 in this study showed an
F-value of 9.93, which indicated that the terms in the model had a significant effect on the response. The probability
p-value of 0.0031 for DHA production reinforced that the model terms are statistically significant at the 95% confidence level. The results of ANOVA also provide a term for residual error, which measures the amount of variation in the response data that was left unexplained by the model. In the present study, the
R2 and Adj
R2 values for DHA production were 0.9210 and 0.8559, respectively. It can, thus, be understood that at least 7% of the experimental results were not explained by the proposed model. The predicted
R2 is the measure of the variation in data explained by the model. The “Pred
R2” of 0.7290 was in reasonable agreement with the “Adj
R2” of 0.8559. The adequate precision value is the signal to noise ratio, and a value >4 is desirable in support of the fitness of the model [
18]. The ratio determined of 9.047 indicated an adequate signal, suggesting that these models can be used to navigate the design space (
Figure 3).
The ratio of triacylglycerols to phospholipids, as well as the fatty acid profile of
Thraustochytrium is known to depend on culture conditions [
19]. The optimization of culture conditions (pH, dissolved oxygen, and temperature) led to a significant increase in the amount of eicosapentaenoic acid (EPA) produced by marine bacteria, where the amount of EPA increased from 33 mg/L to 350 mg/L [
20]. Production of EPA comprises biomass producing and lipid generating stages. During the first phase, the biomass is increased by supplying carbon and nitrogen. In the second phase, carbon alone is supplied to enhance lipid levels. Accumulation of DHA in the form of triacylglycerols or neutral lipids is important for these protists. It was observed that neutral and polar lipids were produced in equal amounts in the early growth phase in
Schizochytrium sp. strain SR21 and microalgae, but on further cell growth only the neutral lipids increased [
21]. The DHA content among total fatty acids was only 8.7% in mature cells of
S.
limacinum SR21 compared to 62% in young ones, suggesting the transfer of the fatty acids of polar lipids to neutral lipids [
22]. The biological significance of DHA in thraustochytrid cells was studied to show that DHA is conserved in the form of fatty acid energy reserves and utilized during starvation [
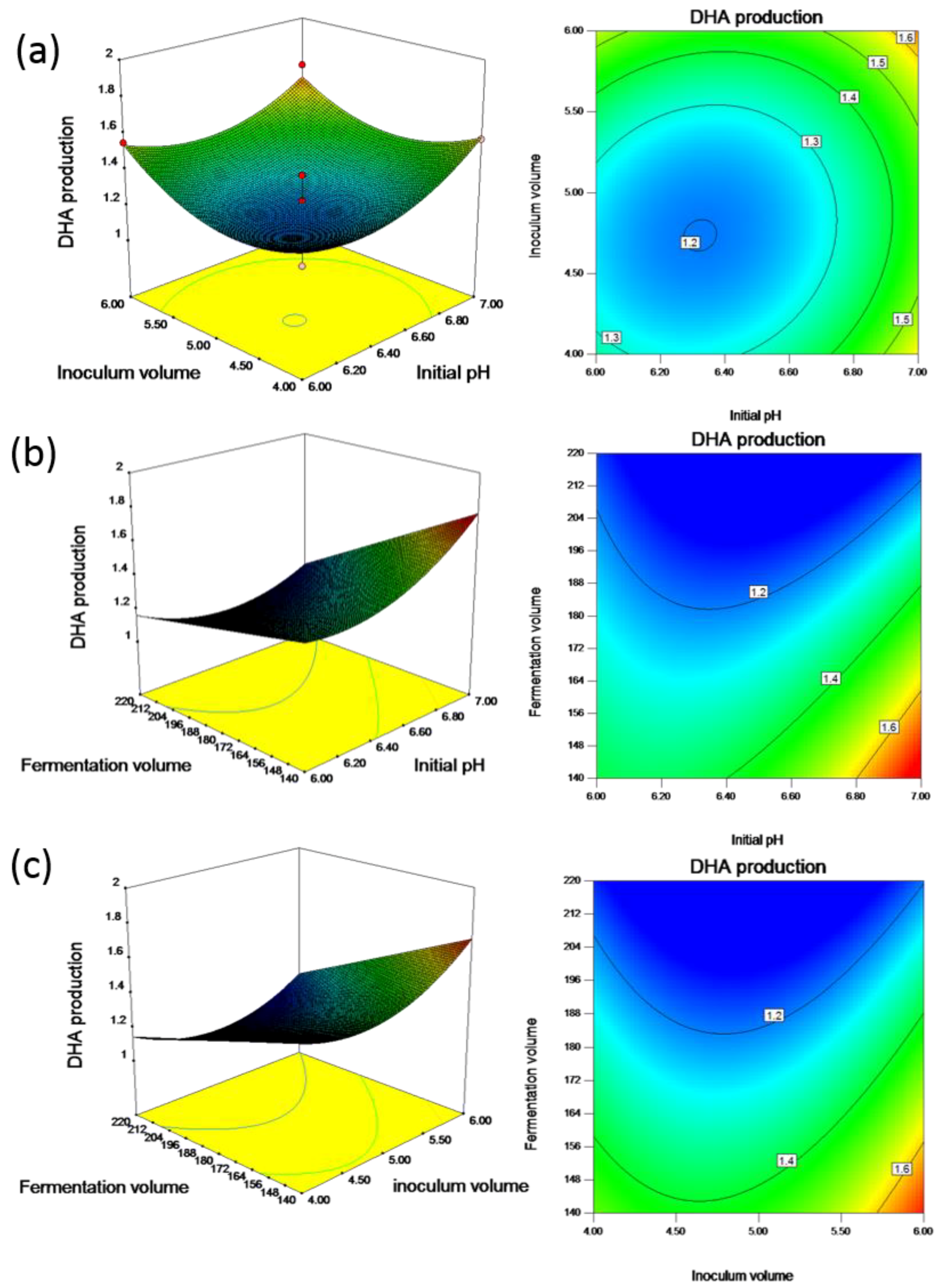
23]. A decline in total lipids was observed with the extension in the starvation phase. Therefore, obtaining the maximum possible biomass with a high amount of total lipids in the shortest possible time is recommended for the strategy in development for DHA production. In this study, the inoculum volume was manipulated for biomass growth and lipid accumulation to reach the stationary phase. It was found that inoculum volume and initial pH interacted negatively, with a coincident decrease in both of these variables resulting in an increase in DHA production. As shown in
Figure 2a, the pattern of the effect of inoculum volume on DHA can be visualized using a three-dimensional response surface plot and corresponding contour plot, and the production of DHA increased as the inoculum volume increased. The size of the inoculum obviously plays a significant role in the growth phase. This likely occurred because the increase in inoculum sizes manipulated microbial growth and lipid biosynthesis, whereas excessive inoculation led to the shortening of the fermentation cycle. The results also showed that an appropriate inoculum volume and initial pH enhanced the production of DHA.
The
p-value of each variable shown in
Table 5 show that the fermentation volume in flasks was the most significant growth factor affecting the growth and DHA production. The increase in the amount of liquid volume in the shake flask could cause the dissolved oxygen to decrease, which inhibits cell growth. It has been reported that the transformation of saturated fatty acids into unsaturated fatty acids occurred when oil-producing strains grow in oxygen-abundant environments [
24]. The volume of the liquid medium was also noted that DO was affected, thus resulting in a lower DHA yield [
25]. A large volume of liquid medium (more than 40 mL in a 250 mL flask) can also lead to a low DHA yield [
16]. The effect of fermentation volume on DHA production may be partially due to the fact that the solubility of oxygen significantly increases at lower liquid volume, which may lead to a relatively higher production of DHA. The higher level of DHA observed at higher DO may be due to the enhanced biosynthesis of DHA as a response to a high DO concentration in the surrounding environment. It has been reported that a stepwise aeration control strategy was adopted for
Schizochytrium sp., whereby biomass reached 71 g/L and total fatty acids reached 35.75 g/L [
26]. It was also observed that a DHA concentration, DHA productivity, and conversion yield of 28.93 g/L, 301 mg/L/h, and 0.44 ± 0.02 g/g, respectively, by
Schizochytrium sp. S31 under constantly high oxygen transfer conditions [
27]. Thus, obtaining high amounts of DHA requires a higher level of DO.
Table 5 indicates that the interaction between the fermentation volume and initial pH has a significant effect on DHA production, whereas the quadratic effect of pH had a significant impact on the amount of biomass and DHA production.
Figure 3b shows the three-dimensional response surface plot for this interaction. From this figure, it can be deduced that providing high levels of fermentation volume and low levels of pH has a negative effect on DHA. Thus, to obtain high amounts of DHA, either one of these factors must be kept at a low level while maintaining the other factor at a high level. The effect alters permeation of ions, acids and bases into the cell, and affects the biochemical metabolism. It was previously found that DHA formation was slightly affected by medium pH with the highest DHA proportion of total fatty acids at pH 7.2 in
Crypthecodinium cohnii, whereas the saturated fatty acids proportion of total fatty acids at this pH value were the lowest [
28]. The production of lipid and DHA by
Schizochytrium sp. S31 incubated at initial pH levels of 5.0 and 6.0 was low. An initial pH of 8.0 for cultivation was too basic for growth and there was a high level of residual glucose, 16 g/L, in the medium after four days of cultivation [
17].
To date, numerous studies have been conducted to enhance DHA production of
Thraustochytrium using various modes of fermentation and strategic techniques. In an attempt to enhance DHA production by
Thraustochytrium sp. ATCC 26185, Furlan et al. performed the conventional one-factor-at-a-time optimization, and the experiment showed the highest DHA yield (1.16 g/L) using different glucose and nitrogen concentrations [
29,
30]. In comparison, this isolate produced 1.68 g/L in the current study using the more sophisticated statistical design of experiments in shake-flask, which was 45% higher than prior to optimization.
4. Materials and Methods
4.1. Microorganisms and Culture Conditions
Thraustochytrium sp. ATCC 26185 was purchased from the American Type Culture Collection (Manassas, VA, USA). An extract produced from
Thraustochytrium sp. ATCC 26185 was previously shown to contain high amounts of lipids, up to 32% of the biomass, and DHA comprised as much as 25% of the total fatty acid content [
31]. This organism was maintained at room temperature on GYP agar plates containing 4 g/L glucose, 2 g/L yeast extract, 2 g/L peptone, 30 g/L artificial seawater salt, and 15 g/L agar. The single colonies from a plate culture were inoculated into 50 mL seeding broth (in 250 mL flask) containing 40 g/L glucose, 5 g/L yeast extract, 5 g/L peptone, and 30 g/L sea salt, then were grown at 28 °C on a shaker incubator with 200 rpm agitation.
4.2. Experimental Design: Plackett-Burman Design
Plackett-Burman design is applied to evaluate the relative importance of several fermentation parameters that influence the desired response, and enables one to screen N variables in at least N + 1 experiments [
32]. To increase the response, each factor selected was tested in two levels, high (+1) and low (−1). A Plackett-Burman design of the experiments was formulated for eight factors using the Design-Expert version 8.0 software (Stat-Ease, Inc., Minneapolis, MN, USA). For the present study, the selected factors included the levels of glucose, yeast extract and sea salt, fermentation volume, inoculum volume (mid-exponential phase), temperature, initial pH, and agitation rate. The levels of each factor were determined based on prior experience. The effect of each variable was estimated from by Equation (2), as follows:
where
and
are the sum of the responses for which variable
is in the high level (
= +1) and the low level (
= −1), respectively. In this equation, N is the total number of trials conducted.
Each trial run was performed in triplicate, and the average of DHA production from Thraustochytrium sp. ATCC 26185 was taken as the response. Pareto chart analysis was used to evaluate the variables, which presents the impact of each variable on DHA production, and the most significant variables were further investigated by Box-Behnken design of response surface methodology.
4.3. Experimental Design: Box-Behnken Design
It is possible to derive an expression for an overall performance measure based on the response values obtained from experiments at particular combinations of input variables [
33]. Box-Behnken design (BBD) was performed to ascertain the main effects of the parameters studied, the interactions among them and the quadratic effect for the responses. A three-level, three-factorial BBD system was used to investigate and validate the fermentation parameters affecting production of DHA by
Thraustochytrium sp. ATCC 26185. To evaluate the influence of significant factors, three independent variables were chosen after Pareto analysis of the Plackett-Burman experimental results: fermentation volume, inoculum volume and initial pH. The ranges of variables were assigned based on path of steepest ascent [
34]. A total of 17 experiments were conducted based on the output of Design Expert software.
The numbers of experiments were calculated as N in Equation (3), as follows:
In Equation (3),
is the variable number and
is the number of the center point. Quadratic and interaction effects of the independent variables were evaluated for RSM. The sequential
F-test, lack-of-fit test and other adequacy measures were used in selecting the best model [
35]. The relationship between the response and the input process parameters were performed by using Equation (4):
In Equation (4), ƒ is the real response function its format being unknown, and
is the residual error which describes the differentiation that can be included by the function ƒ. In consideration of the linear terms, square terms and linear interaction items, the second order equation response was modeled by Equation (5) to correlate the dependent and independent variables, as follows:
In Equation (5), is the predicted response, is the model constant, is the regression coefficient for the input factor , is the regression coefficient for interactions, and is the regression coefficient for square effects between the input factor and . The optimum values of the selected variables were determined by solving the regression equation and analyzing the three-dimensional surface plots. A fermentation experiment was employed to evaluate the DHA production of Thraustochytrium sp. ATCC 26185 using the predicted optimum parameter values.
4.4. Fatty Acid Methyl Ester Extraction, Preparation, and Analysis
Dried cells (20 mg) were suspended in 1.5 mL methanol/acetyl chloride (10:1, v/v), 1 mL of hexane and 15 μg nonadecanoic acid were added and heated at 70 °C for 2 h in sealed tubes. After cooling, 2.5 mL 6% K2CO3 and 1 mL hexane were then added to the extracts, the mixture shaken, centrifuged, and the upper layer separated.
The profiling of extracted FAMEs was performed by GC-MS with a SPB-50 fused silica capillary column, 30 m × 0.25 mm × 0.25 μm (Supelo, Bellefonte, PA, USA) installed in a QP2010 GC equipped with an AOC-20 autosampler (Shimadzu, Kyoto, Japan) and mass spectrometer (Shimadzu, Kyoto, Japan). The temperature of the injector was set to 250 °C. High-purity helium was used as the carrier gas, with a column flow rate of 0.81 mL min−1 and a pre-column pressure of 73.0 kPa. After injection, the oven temperature was kept at 150 °C for 3.5 min, increased at 20 °C/min to 200 °C, held for 5 min, then increased at 5 °C/min to a final temperature of 280 °C, where it was held for 30 min. The sample injection volume was 1 μL with a split ratio of 50:1. The MS was operated in electron impact (EI) mode with an electron energy of 70 eV. Furthermore, the source temperature was set at 200 °C, and the transfer line temperature was 250 °C. The mass spectrometer was set to scan from m/z 50 to 600. There was a 3.5 min solvent delay time. Nonadecanoic acid (C19:0) (Sigma Aldrich, Dorset, UK) was used as an internal standard and its peak area was used to determine the amount of each fatty acid based on the areas of all peaks and the known concentration of the standard added.






{kind=link}
{kind=link}
{kind=link}