1. Introduction
Insomnia, depression and Alzheimer’s disease (AD) are severe diseases of the central nervous system (CNS). Neurotransmitters (NTs) are widely recognized to make the closest contact with CNS diseases. Abnormal alteration of NTs has been found to be nearly concerned to several neurological diseases including insomnia, depression, AD, Huntington’s disease and Parkinsonism etc. [
1]. Serotonin (5-HT), a tryptophan-derived biogenic amine, eventually endures oxidative deamination to form 5-hydroxyindolacetic acid (5-HIAA), has been found decreasing significantly in brains of insomnia [
2], depression [
3] and AD [
4] rats. Dopamine (DA) and norepinephrine (NE), the metabolites of tyrosine (Tyr), are involved in sleep-wake cycles, learning, memory and emotion. A number of studies [
2,
3,
4] have shown that the levels of DA and NE decreased significantly in the brains of depression and AD rats, counter to insomnia. Acetylcholine (ACh) is recognized participant in rapid eye movement (REM) sleep [
5]. The increasing release of ACh in the pontine reticular formation promoted REM sleep in murine [
6,
7]. Studies suggested that cholinergic insufficiency contributed to several of the most significant neuropsychiatric appearance of AD [
8] and the decreasing breakdown of ACh induced symptoms of anxiety and depression [
9]. γ-aminobutyric acid (GABA) and glutamic acid (Glu) are reported as major inhibitory and excitatory NTs, in CNS, respectively. The lack of transformation from Glu to GABA leading strong excitement and weak inhibition is one of causes of some neurological diseases such as insomnia [
2], AD [
10] and depression [
11,
12]. It was found that most studies have only investigated if NTs increasing or decreasing in insomnia, depression and AD, respectively, rather than revealing differences and similarities of NTs changes from an overall perspective. So, monitoring neurotransmitter and their metabolite levels in rat brain tissues is an essential tool for examining the similarities and differences in the pathophysiology of these three diseases and also a good method to study the differences in the nature of these interrelated neuropsychiatric disorders.
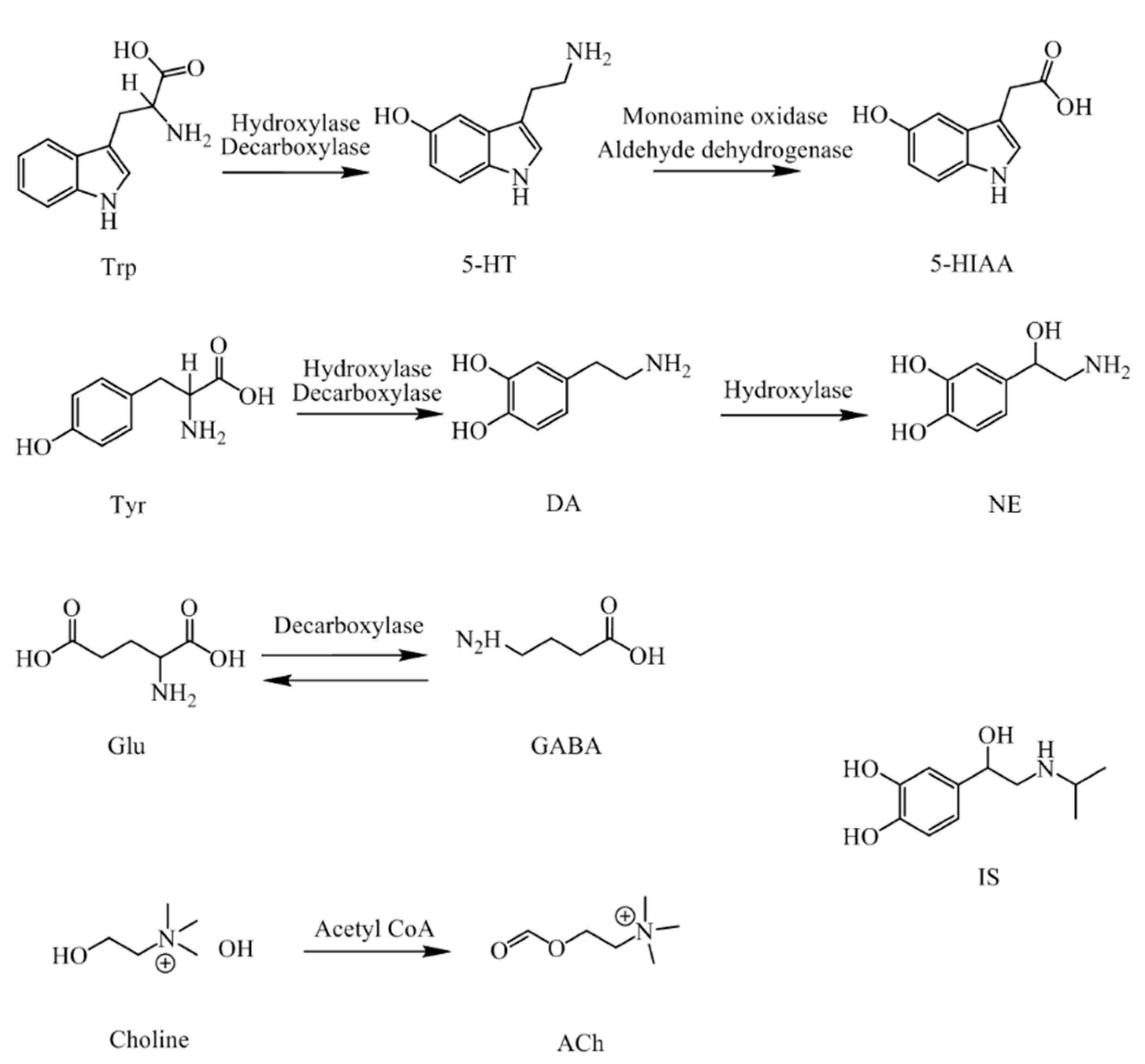
NTs and their metabolites are exist extensively in CNS and the peripheral biofluids of mammals [
13,
14], including neurotransmitters of amino acid, such as Trp, Tyr, ACh, Glu and GABA, and monoamine neurotransmitters, such as 5-HT, DA, and NE, and also their acidic metabolite 5-HIAA. Their molecular structures and pathways are presented in
Figure 1.
Numerous analytical methods, including high performance liquid chromatography (HPLC) or capillary electrophoresis (CE) involving a series of techniques, such as ultraviolet detection (UV), electrochemical detection (ECD), fluorescence detection (FLD), laser induced fluorescence detection (LIFD) and mass spectrometry (MS), have been engaged in the analysis of NTs in biological samples [
15,
16,
17,
18]. Whereas, except the narrow scope of application, all of the analytical procedures have several different drawbacks, such as, low sensitivity and selectivity for UV, poor repeatability and difficulty in simultaneous separation of NTs which have similar electrophoretic behavior, such as Glu and GABA for ECD, and interference after derivatization for FLD. Conventional methods of LC-MS/MS have been used to determine the concentration of NTs with more complex sample preparation, a longer assay time and fewer analytes [
19,
20]. Therefore, it is necessary to set up a procedure that is simpler, faster and can be used to quantify more analytes simultaneously.
Extracting important features from data is a vital procedure in the issue of pattern recognition, not just can it decrease the computational complexity, but result in better performance through eliminating data redundancy and noise, while overcoming dimension problem for a statistical classifier [
21]. As to large category pattern classification, linear supervised feature extraction approach is engaged widely. Bayes linear discriminant method is widely accepted as a good way for the analysis of reliability data, which calculates the between scatter matrix and the within scatter matrix only, then figures out the problem of eigenvalue decomposition easily with efficient computation. To compare different changes of NTs in rat models of insomnia, AD and depression, in our study, a rapid and reliable LC-MS/MS method was established to determinate nine NTs and their metabolites simultaneously in rat brains with a higher sensitivity and shorter chromatographic separation time. The parameters of linearity, precision, recovery and matrix effect were completely validated and successfully applied to analyze the NTs in rat models of insomnia, AD and depression. A Bayes linear discriminant function was established to distinguish these three kinds of nervous system diseases by DA, Tyr and ACh for their significant differences among control and three model groups. The analytical method in this study can also be applied to not only brain issues but also plasma and urine samples, moreover the conclusions of this paper can be used to distinguish insomnia, depression and AD models, which can assist research of neurological diseases. Finally, the marked changes in biomarkers might be applied to clinical diagnosis.
3. Materials and Methods
3.1. Materials and Reagents
5-HT, 5-HIAA, Glu, GABA, DA, NE, ACh, Trp and Tyr were purchased from Sigma (St. Louis, MO, USA). The internal standard isoproterenol (IS) was from the Institute for Food and Drug Control (Liaoning, China). 4-Chloro-D,L-phenylalanine (PCPA), D-galactopyranose (D-gal) and Amyloid β-Peptide Fragment 25–35 (Aβ25–35) were also supplied by Sigma-Aldrich (St. Louis, MO, USA). Distilled water was used throughout the experiments. HPLC grade methanol and acetonitrile were from Fisher Scientific (Fair Lawn, NJ, USA). HPLC grade reagents such as acetic acid, formic acid, sodium hydroxide and ammonium hydroxide were provided by Shandong Yuwang Industrial Co., Ltd. (Yucheng, China). Other reagents were all analytical grade.
3.2. Animals and Treatment
Forty-eight male wistar rats (200–220 g) were kindly provided by the Experimental Animal Center of Shenyang Pharmaceutical University and bred with unlimited access to food and water in an air-conditioned animal center at a temperature of 22 ± 2 °C and a relative humidity of 50 ± 10%, with a natural light-dark cycle. The animal study was carried out in accordance with the Guideline for Animal Experimentation of Shenyang Pharmaceutical University and the protocol was approved by the Animal Ethics Committee of the institution (Ethic approval document NO. SYPU-IACUC-C2017-1-31-203).
The animals were allowed to acclimatize to the environment for one week before the experiment. Forty-eight rats were randomly divided into six groups, with eight animals each: the insomnia control and model group, the AD control and model group, the depression control and model group. The methods of treating animals used hereinafter are recognized as scientific and effective, widely used [
22,
30,
31,
32,
33].
Animal model of insomnia was induced by the chemical reagents according to precious report [
34]. Briefly, PCPA was suspended in physiological saline. The animal of insomnia model group was administrated intraperitoneal injection of PCPA (350 mg kg
−1), while the control group was given the same volume of saline solution once a day and last for four days, respectively. After the injection of PCPA, the model group lost their weight and circadian rhythm, and were sleepless all day which proved the success of the model.
The AD model in rats was induced by D-gal and Aβ
25–35, as described previously [
35]. In brief, the rats in AD model and control groups were respectively given intraperitoneal injection of D-gal (50 mg kg
−1 day
−1) and the same volume of saline for 6 weeks; then the AD rats were injected with 5 μL Aβ
25–35 (10 μg μL
−1) into each bilateral hippocampus at the co-ordinates antero-posterior, −3.5, medio lateral, +2.0, dorso-ventral, 2.8 mm according to the stereotaxic atlas on the fourth week by brain stereotaxic apparatus, individually, while the control rats were administrated the same volume of saline in the same way. After 6 weeks, all rats were trained to find a visible platform in Morris water maze testing. The result demonstrated that the rats in AD model group had a longer time to get to the platform than the control rats at the last trial (
p < 0.05), which proved the success of the AD model.
The depression model was induced by chronic unpredictable mild stress (CUMS). The model rats were kept separately. Control animals were housed in a separate room and had no contact with the stressed animals. The CUMS procedure was referenced from the precious study [
36]. Stressors consisted of (1) 4 h restraint; (2) 24 h wet litter; (3) 24 h food deprivation; (4) 60 s tail pinch; (5) 24 h water deprivation; (6) 5 min cage shake; (7) 12 h cage tilt (cages were tilted to 45° from the horizontal). In this respect, stressors were administrated in a semi-random manner at any time of day, and the stress sequence was changed every week in order to make the stress procedure unpredictable. After 3 weeks the autonomous behavior of rats was evaluated by the Open-field test (OFT). The result demonstrated that the vertical and horizontal bouts of rats in depression model group were lower than the control group (
p < 0.05), which proved the success of the CUMS model.
3.3. Instruments and LC–MS/MS Conditions
The LC-ESI-MS/MS system was performed using an LC-20 A Prominence™ UFLC XR system consisted of a binary pump, a degasser, an autosampler and a thermostatted column compartment (Shimadzu, Kyoto, Japan); a 4000 QTRAP™ triple quadrupole-linear ion trap mass spectrometry system equipped with a turbo ion spray source (Sciex, Foster City, CA, USA). All the operations, the acquiring and analysis of data were controlled by Analyst software (version 1.6, Sciex).
Separations were accomplished on an Inertsil ODS-EP column (4.6 mm × 150 mm, 5.0 μm) (GL Sciences, Tokyo, Japan) protected by a high-pressure column pre-filter (2 mm) at 35 °C. The mobile phase consisting of 0.01% formic acid in water (A) and methanol (B) was delivered at a flow rate of 1.2 mL min
−1 with one third of the eluent splitted into the inlet of mass spectrometer. The linear gradient elution program was as follows: 0–0.5 min, 15% B; 0.5–5 min, 15–85% B; 5–6 min, 85% B; 6–8 min, 15% B. The injection volume was 4 μL and the total time taken for the chromatographic run was 8.0 min. The analytes and IS were ionized by the ESI source in positive ion mode and the ion spray voltage was set at 5500 V. The curtain gas, gas 1 and gas 2 (nitrogen) were set at 20 psi, 50 psi and 50 psi with a source temperature of 500 °C. Quantitative MRM ion pairs parameters are listed in
Table 6.
3.4. Standard Solution and Quality Control Samples
Standard stock solutions of 5-HT, 5-HIAA, Glu, GABA, DA, NE, ACh, Trp, Tyr and IS were separately prepared in methanol-water (20:80, v/v). The stock solutions of the analytes were further diluted with methanol-water (20:80, v/v) to make a series of mixed working standard solutions at the desired concentrations. The brain tissue samples were homogenized in a twenty-fold volume of methanol with 1% formic acid. The brain tissue standards of analytes for each of the nine analytes were prepared as follows, GABA and Glu at concentrations of 1, 2, 4, 8, 16 and 32 μg mL−1, 5-HT and 5-HIAA at concentrations of 2, 4, 8, 16, 32 and 64 ng mL−1, NE and Tyr at concentrations of 100, 200, 400, 800, 1600 and 3200 ng mL−1, Trp, DA and ACh at concentrations of 20, 40, 80, 160, 320 and 640 ng mL−1. These standards were prepared by adding appropriate amount of standard working solutions to blank brain tissue homogenates. Quality control samples were prepared using the same method (4, 16 and 50 ng mL−1 for 5-HT and 5-HIAA; 2, 8 and 25μg mL−1 for Glu and GABA; 200, 800 and 2500 ng mL−1 for NE and Tyr; 40, 160 and 500 ng mL−1 for Trp, DA and ACh. A working solution for the IS (10 μg mL−1) was also prepared.
3.5. Sample Preparation
All rats were sacrificed by decapitation without anesthesia. The brains were rapidly removed above ice bath, frozen and stored at −80 °C until extraction. All frozen samples, calibration standards and QC samples were thawed and allowed to equilibrate at room temperature prior to analysis. The frozen brain tissue samples were dissected and homogenized in a twenty-fold volume of methanol with 1% formic acid above ice-water bath. The homogenates were centrifuged at 12,000 × g for 20 min at 4 °C. Then, 100 μL of the supernatant was transferred to a 1.5 mL centrifuge tube, spiked with 10 μL of the IS solution and 10 μL methanol, followed by vortexing for 30 s. Then 100 μL methanol was added into the supernatant, followed by vortexing for 1 min. Next, these samples were centrifuged at 12,000× g for 5 min at 4 °C. Finally, a 4 μL aliquot was injected into the LC-MS/MS system for analysis.
3.6. Method Validation
The analytical method validation procedure was in accordance with the US Food and Drug Administration Bioanalytical Method Validation Guidance for Industry [
37]. Validation parameters included lower limit of quantification (LLOQ), linearity, precision, accuracy, recovery, matrix effect and stability.
Neurotransmitters are endogenous substances which already existed inherently in biosamples, especially brain homogenates. Therefore, blank value of each analyte should be subtracted from each calibration point. The calibration curve was constructed for each analyte by plotting the increased analyte-to-IS peak area ratio (y) between the brain tissue homogenates that spiked standard solutions and the mean ratio of blank samples versus nominal concentration (x) by 1/x2 weighted least square linear regression. LLOQ was defined as the lowest concentration of the calibration curve with acceptable accuracy and precision, which provided an intensity of signal-to-noise ratio above 10.
The validation of accuracy and precision and were performed with six replicates at three (low, medium and high) QC concentration levels on the same day and on three consecutive days. Precision was presented as the relative standard deviation (RSD %), while accuracy was presented as relative error (RE%).
The extraction recovery for each analyte was calculated by comparing the difference of the responses between the spiked sample before and after extraction at three QC levels, while the mean value of blank samples was removed.
The matrix effect was evaluated as follows: the analyte-to-IS peak area ratio for each analyte in the spiked sample after extraction subtracted the mean ratio of blank samples at three concentration levels of QC samples, and then the increased peak area ratio was divided by the mean ratio measured in corresponding standard solutions.
Stability tests were conducted on triplicate at low and high QC levels under the storage conditions as follows: 8 h at room temperature, 4 °C in the autosampler for 8 h after preparation, three freeze-thaw cycles and stored at −80 °C for three months.
3.7. Statistical Data Analysis
All data are illustrated as the mean ± SD. The acquired data were analyzed by ANOVA and Bayes discriminant analysis using the SPSS 20 statistical software (IBM, Armonk, NY, USA). A p value less than 0.05 was considered statistically significant for all the tests.
4. Conclusions
In this paper, a simple, fast and sensitive LC-MS/MS method using an ODS-EP column, without derivatization or ion-pairing reagents for simultaneous determination of nine NTs in rat brain was established and fully validated. Following optimization of the chromatographic conditions, the compounds were found to have symmetric peak shapes and could be assayed in 8.0 min, which was very suitable for high throughput bioanalysis. A simple sample preparation involving extraction was applied and the recoveries of all the analytes above 85.0%. In addition, we discovered changes in several NTs and their metabolites in rats with insomnia, depression and AD, and established a Bayes linear discriminant function to differentiate these three kinds of neurological diseases. Results indicated that there were significant differences for 5-HT, DA, NE, Trp, Tyr and ACh between model and control group in all three models, what is more, DA, Tyr and ACh showed significant differences among control and three model groups and were used to distinguish these three kinds of nervous system diseases successfully with a cross-validation accuracy rate 86.0%.The data obtained in this study make a promising understanding to the diagnosis and treatment of insomnia, depression and AD.

 ,
, 


{kind=link}
{kind=link}