
The Emerging Role of Polyphenols in the Management of Type 2 Diabetes


1
Department of Human Nutrition, Foods and Exercise, College of Agricultural and Life Sciences, Virginia Tech, Blacksburg, VA 24060, USA
2
Department of Nutrition and Food Technology, Jordan University of Science and Technology, Irbid 22110, Jordan
*
Author to whom correspondence should be addressed.
†
Current affiliation: Diabetes Center, University of California San Francisco, San Francisco, CA 94143, USA.
Molecules 2021, 26(3), 703; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030703
Submission received: 13 November 2020
/
Revised: 25 January 2021
/
Accepted: 26 January 2021
/
Published: 29 January 2021
(This article belongs to the Special Issue Bioactive Compounds on Health and Disease)
Abstract
:Type 2 diabetes (T2D) is a fast-increasing health problem globally, and it results from insulin resistance and pancreatic β-cell dysfunction. The gastrointestinal (GI) tract is recognized as one of the major regulatory organs of glucose homeostasis that involves multiple gut hormones and microbiota. Notably, the incretin hormone glucagon-like peptide-1 (GLP-1) secreted from enteroendocrine L-cells plays a pivotal role in maintaining glucose homeostasis via eliciting pleiotropic effects, which are largely mediated via its receptor. Thus, targeting the GLP-1 signaling system is a highly attractive therapeutic strategy to treatment T2D. Polyphenols, the secondary metabolites from plants, have drawn considerable attention because of their numerous health benefits, including potential anti-diabetic effects. Although the major targets and locations for the polyphenolic compounds to exert the anti-diabetic action are still unclear, the first organ that is exposed to these compounds is the GI tract in which polyphenols could modulate enzymes and hormones. Indeed, emerging evidence has shown that polyphenols can stimulate GLP-1 secretion, indicating that these natural compounds might exert metabolic action at least partially mediated by GLP-1. This review provides an overview of nutritional regulation of GLP-1 secretion and summarizes recent studies on the roles of polyphenols in GLP-1 secretion and degradation as it relates to metabolic homeostasis. In addition, the effects of polyphenols on microbiota and microbial metabolites that could indirectly modulate GLP-1 secretion are also discussed.
1. Introduction
Type 2 diabetes (T2D) is becoming a global health problem [1], presently affecting 34.2 million (or 10.5%) people in the U.S. alone [2], with 90% of them being overweight or obese [3]. It is well-established that T2D results from insulin resistance in insulin sensitive tissues and subsequent pancreatic beta-cell dysfunction [4,5]. Therefore, most of the therapeutic interventions for T2D focus on lowering blood glucose levels by stimulating insulin secretion, decreasing hepatic glucose production, or enhancing insulin action at target tissues [6,7]. Gastrointestinal (GI) tract plays a primary role in regulating glucose homeostasis, in particular, postprandial blood glucose levels. In addition to digesting and absorbing nutrients, the intestine secretes hormones and generates neuronal signals in response to the ingested nutrients that coordinately exert profound metabolic effects [8,9,10,11]. Of these, the incretins, which are a group of metabolic hormones that have an insulinotropic action, have drawn considerable attention for developing novel therapeutic strategy for treating T2D [12]. Incretin hormones accounts for over 50% of postprandial insulin secretion in healthy individuals [13]. Notably, glucagon-like peptide-1 (GLP-1), an incretin secreted from enteroendocrine L-cells after food ingestion [14], plays a pivotal role in glucose homeostasis via potentiating glucose stimulated insulin secretion (GSIS) and promoting β-cell proliferation and survival [15,16,17]. In addition, GLP-1 delays gastric emptying, induces satiety, and reduces body weight in animal models of obesity and obese humans [18,19]. In addition, the gut microbiota, a large and diverse microbial community in the colon, is involved in metabolizing dietary components, primarily indigestible carbohydrates and phenolic compounds [20], thereby, affecting host’s biological activity at the level of the intestine and the whole body. Indeed, increasing evidence shows that the gut microbiota play an important regulatory role in metabolic homeostasis of the host, as alterations in the gut microbiota, termed dysbiosis, are associated with obesity and the pathogenesis of T2D [21].
A mounting body of evidence from preclinical studies indicates that dietary polyphenols, present in many fruits, vegetables, and medicinal herbs, may exert anti-diabetic effects [22,23,24,25]. A recent epidemiological study showed that intake of polyphenol-rich diet was associated with a lower risk of diabetes [26]. Likewise, clinical trials showed beneficial effects of polyphenols on glucose metabolism. For instance, daily intake of apple (Malus pumila cv. Fuji) polyphenols for 12 weeks significantly improved glucose tolerance in humans [27]. While various potential mechanisms for the anti-diabetic effects of polyphenolic compounds have been proposed, they are largely based on the in vitro studies with the use of pharmacological doses that often yield confounding or inconsistent results. For example, naringenin, a predominant flavanone from citrus fruit, was found to increase glucose uptake in cultured skeletal muscle cells [28], but suppress intestinal glucose absorption [29]. Similarly, the flavonol quercetin increased glucose uptake of muscle cells but inhibited the uptake in adipocytes and intestinal cells [30,31,32]. In these studies, the lowest effective concentration of these polyphenolic compounds for achieving the observed results was at least 10 µM. However, the circulating concentrations of most polyphenols, including their metabolites following dietary intake, are typically less than 5 µM [33], suggesting that many in vitro findings from using pharmacological doses of polyphenols might be physiologically irrelevant. Thus, the exact anti-diabetic mode and the major target tissues of polyphenols in vivo have not been fully elucidated.
Given the low bioavailability but relatively high concentrations of polyphenols in the GI tract following dietary ingestion, it is likely that the intestine might be a primary site for polyphenols to exert the observed metabolic actions [34]. In that regard, polyphenols may influence glucose homeostasis partially through modulating glucose and fatty acid digestion and absorption [35]. Interestingly, emerging evidence suggests that phenolic compounds also affect gut hormone release and microbiota composition, which are then directly or indirectly involved in regulating metabolic homeostasis. For example, it was shown that intake of coffee polyphenol increases postprandial GLP-1 concentration [36], which was associated with the lower risk of T2D in the individuals consuming 6 or more cups of coffee daily [37]. Epigallocatechin gallate (EGCG), a main polyphenol from green tea (Camellia sinensis), suppresses the secretion of orexigenic peptide hormone, ghrelin [38], thereby reducing food intake in rodents [39]. Polyphenol-enriched beverage that made from turmeric increases the postprandial concentration of anorexigenic hormone, peptide tyrosine tyrosine (PYY) in the healthy individuals [40], suggesting a potential anti-obesity action of turmeric polyphenols [41]. In this review, we first provide an overview about GLP-1 and the regulatory mechanisms for its secretion, and summarize recent developments in the effects of polyphenolic compounds on GLP-1, with emphasis on the signaling pathways activated by polyphenolic compounds in the regulation of GLP-1 secretion. In addition, the effect of polyphenols on microbiota and subsequent microbial metabolites is discussed as they relate to GLP-1 secretion. Lastly, emerging literature exploring the potential of polyphenols as dipeptidyl-peptidase-IV (DPP-IV) inhibitors is also highlighted.
2. Regulation of GLP-1 Secretion
2.1. GLP-1 Physiology
The gut is the largest endocrine organ in the body and releases an array of hormones, which play many key roles in physiological and metabolic regulation [42]. Specifically, GI hormones secreted after a meal plays a critical role in regulating appetite, food intake, and glucose homeostasis [11]. GLP-1 is a well-studied incretin hormone secreted by L-cells in the gut [43], the second largest population of enteroendocrine cells [16,44]. GLP-1 is produced from post-translational processing of the proglucagon gene in the L-cells [45], leading to the generation of two major forms of GLP-1, GLP-1-(7–37) and GLP-1-(7–36)NH2, which are then released into the bloodstream [46]. GLP-1 has a very short half-time (2–3 min) in the blood because it is rapidly degraded by DPP-IV that cleaves GLP-1 from its N-terminal into GLP-1-(9-37) and GLP-1-(9-36) NH2 [43].
The L-cells are distributed throughout the intestine with much higher density in the ileum and the colon. The polarized morphology of L-cells allows its apical surface to face the lumen of the intestine thus facilitating sensing of nutritional, neural, and hormonal signaling by the cells [18,47]. The role of GLP-1 in maintaining glucose homeostasis is mediated through its receptor (GLP-1R). GLP-1 binding to its receptor triggers cyclic adenosine monophosphate (cAMP) signaling that augments GSIS from pancreatic β-cells while inhibiting glucagon release from α-cells [48]. In addition, the activation of GLP-1 signaling promotes β-cell proliferation and survival, therefore preserving islet mass [49]. Furthermore, GLP-1 slows the gastric emptying and gut mobility, and probably also targets its receptor in the hypothalamus to promote satiety, thereby reducing food intake [50,51]. Impaired GLP-1 secretion in response to intake of meals or glucose has been reported in obese and T2D individuals [52,53,54,55]. However, a systematic analysis of GLP-1 secretory data from human studies concluded that patients with T2D, particularly those with relatively low HbA1c did not show reduced GLP-1 secretion in response to glucose or a mixed meal [56]. In addition to ingested nutrients, GLP-1 secretion could also be influenced by other factors such as sex [55], obesity, insulin resistance, β-cell function, and diabetes state [57]. Therefore, it remains to be determined whether the defect of nutrient-induced GLP-1 secretion occurs early and is responsible for the pathogenesis of T2D. Regardless, activation of the GLP-1 signaling system is effective for the treatment of T2D, while also exerting other beneficial effects, such as promoting weight loss and β-cell function, and improving cardiovascular parameters [58]. Several GLP-1R agonists have been approved by the FDA for treating T2D. The first GLP-1R agonist, exenatide, also known as Byetta, shared 55% homology to human GLP-1, and when administered with a dose of 5 µg, it significantly improved hyperglycemia in T2D patients [59]. Consistently, another recently developed GLP-1R agonist, liraglutide, which has 96% homology with human GLP-1, showed more efficacious glycemic control as compared with that of exenatide [60].
2.2. Neuronal and Hormonal Regulation of GLP-1 Secretion
GLP-1 secretion is primarily stimulated by ingested nutrients. It has been observed that GLP-1 is secreted into the circulation in a biphasic pattern, with an initial rapid rise in circulating GLP-1 levels occurring 15 to 30 min after food ingestion, followed by a prolonged second phase about 60 min later [61]. Neuroendocrine regulators rather than the direct sensing of L-cells to the nutrients, should be primarily responsible for the early rise of GLP-1 levels because the digested nutrients are unable to reach the ileum or colon within 30 min where L-cells are primarily located [62]. Indeed, it was found that nutrients in the proximal intestine are able to trigger GLP-1 secretion from the distal gut via activation of vagus nerves [63,64]. In a study performed in rats, it was found that left cervical vagotomy reduced the basal GLP-1 concentration [63], suggesting that vagus nerve plays a role in maintaining basal GLP-1 release. In addition, infusion of nutrients into the proximal duodenum failed to increase GLP-1 secretion in rats with subdiaphragmatic vagotomy [63], confirming that vagus nerve is responsible for the first phase of postprandial GLP-1 secretion. Acetylcholine, a well-known neurotransmitter, is involved in the vagal nerve activation-induced GLP-1 secretion [65]. In that regard, acetylcholine released from vagal efferent nerves stimulates GLP-1 secretion via its receptors, muscarinic receptors, which are expressed on the L-cells [66]. There are reportedly two types of muscarinic receptors (M1 and M2) on the surface of L-cells. M1 muscarinic receptor is coupled to Gαq/11 and its activation triggers phospholipase C (PLC)/inositol trisphosphate (IP3)/Ca2+ signaling leading to GLP-1 release, while the M2 muscarinic receptor is coupled to Gαi, and its activation inhibits adenylyl cyclase and attenuates the cAMP signaling pathway [67], suggesting that M1 receptor plays a major role in mediating acetylcholine effect on GLP-1 secretion, as activation of M2 muscarinic receptor could potentially inhibit GLP-1 secretion via suppressing cAMP signaling [67].
Other gut hormones may affect GLP-1 secretion. Glucose-dependent insulinotropic peptide (GIP), which is secreted from the small intestinal K-cells in response to nutrient intake [43], may also be involved in the regulation of GLP-1 secretion. GIP-induced GLP-1 secretion might, in part, depend on the activation of the vagus nerve. In rodents, the administration of GIP at a supraphysiological dose increased GLP-1 release, however, this effect was abrogated by hepatic branch vagotomy [63]. In addition, the GIP may directly stimulate GLP-1 release via its receptor on L-cells, as demonstrated in cultured fetal rat intestinal cells and GluTag L-cells [68]. In addition to GIP, ghrelin, an orexigenic gut hormone mainly released from the stomach, was reported to enhance postprandial GLP-1 secretion. The plasma concentration of ghrelin increases during fasting and decreases after refeeding [69]. In clonal L-cells, ghrelin triggered GLP-1 secretion via activation of the ghrelin receptor and extracellular signal-related kinase 1/2 (ERK1/2)-dependent pathway [70]. In vivo, the intraperitoneal administration of ghrelin enhanced glucose-stimulated GLP-1 secretion (GSGS) and improved glucose tolerance in mice, while pharmacological inhibition of ghrelin receptor blocked ghrelin-induced GLP-1 secretion [71]. Consistently, ghrelin-induced improvement in glucose tolerance was diminished in mice administered with a GLP-1R antagonist and in GLP-1R-deficient mice [71]. These outcomes suggest that the effect of ghrelin on glucose tolerance is mediated through the GLP-1/GLP-1R pathway. Such an interaction between ghrelin and GLP-1 secretion in response to fasting and refeeding demonstrates a precise and reciprocal regulation of food intake in the body. In that regard, the increase in ghrelin levels during fasting stimulates appetite and thus increases food intake. Subsequently, the ingestion of food stimulates GLP-1 secretion, which in turn reduces appetite and decreases ghrelin secretion [72].
2.3. Nutritional Regulation of GLP-1 Secretion
As discussed above, GLP-1 is secreted in a biphasic pattern in response to the ingested meal. The second phase of GLP-1 secretion is largely stimulated by the digested macronutrients via their respective receptors on the plasma membrane of L-cells [73,74] (Figure 1). Sweet taste receptors 1 (Tas1Rs) have been reported to mediate monosaccharide-induced GLP-1 secretion [75]. Tas1Rs are present in the GI tract of both rodents and humans [75,76]. The activation of these receptors by glucose on the L-cells triggers GLP-1 secretion via the PLC/IP3/Ca2+-sensitive transient receptor potential channel M5 (TRPM5) pathway [77]. Consistently, blockage of Tas1R2/Tas1R3 reduced the increase in circulating GLP-1 concentrations in response to glucose administration in humans [78]. In addition, glucose could induce GLP-1 secretion in a Tas1R-independent pathway, where the glucose is transported into the cells via sodium-dependent glucose transporter 1 (SGLT1) and undergoes glycolysis and oxidation, resulting in increased ATP production and subsequent closure of KATP channels, which sequentially causes depolarization of the plasma membrane, opening of voltage-gated L-type Ca2+ channels, Ca2+ influx, and ultimate GLP-1 secretion [79,80]. Thus, inhibition of glucose uptake could prevent the cascade of these events and ablate GSGS [81].
Fatty acids (FAs)-stimulation of GLP-1 secretion is largely mediated via their respective receptors, which belong to G-protein coupled receptors (GPCRs). Specifically, four FA receptors (FFARs), FFAR1 (GPR40), FFAR2 (GPR43), FFAR3 (GPR41), and FFAR4 (GPR120), which are coupled to G-proteins, are involved in FA-induced GLP-1 secretion [73,74]. The binding of FFAR1 or FFAR4 by their ligands, mainly medium to long chain FAs, like linoleic acid [82,83] and omega-3 polyunsaturated FAs (PUFAs) [84], activates Gαq, which stimulates PLC to cleave membrane-associated phosphatidylinositol 4,5-bisphosphate (PIP2) into IP3 and diacylglycerol (DAG). The released IP3 binds to the IP3 receptor on the ER, triggering Ca2+ release from the ER and subsequent GLP-1 secretion [85]. In addition, DAG can also induce GLP-1 secretion through activation of the protein kinase C (PKC)/protein kinase D (PKD) pathway [86]. FFAR2 and FFAR3 are activated by short-chain FAs (SCFAs), in particular, acetate, propionate, and butyrate [74,87]. These SCFAs are generated from microbial fermentation of non-digestible dietary nutrients, such as fiber and other indigestible carbohydrates [88]. FFAR2 is coupled to both Gαi and Gαq, whereas FFAR3 is only coupled to Gαi and activation of FFAR3 in CHO cells overexpressing FFAR3 resulted in Gαi-mediated suppression of intracellular production of cAMP [89]. It was shown that mice deficient of FFAR2 had a blunted GLP-1 secretion in vivo [74]. Similarly, FFAR3-deficient mice also displayed impaired GLP-1 secretion in response to the administration of SCFAs [74], suggesting that both FFAR2 and FFAR3 play a role in SCFA-induced GLP-1 secretion. Although the mechanism is still unclear, it is possible that the impaired GLP-1 secretion in FFAR3-deficient mice could be due to the reduced FFAR2 expression [74]. However, more recent work using the perfused rat colon demonstrated that specific activation or blockage of FFAR2 or FFAR3 had no effect on colonic GLP-1 release, suggesting that SCFAs may trigger GLP-1 secretion independent of the FFAR2 and FFAR3 [90]. Therefore, it remains to be determined whether L-cell-specific FFAR2/FFAR3 are involved in mediating SCFA-stimulated GLP-1 secretion.
The ingestion of high-protein diets also was shown to increase GLP-1 secretion in rodents and humans [91,92]. Protein-induced GLP-1 secretion is likely executed via its enzymatic products, peptides and amino acids (AAs), which might act through peptide transporter-1 (PEPT1) and calcium-sensing receptor (CaSR) to induce GLP-1 secretion [93]. PEPT1 has a rather broad substrate specificity, as it has been demonstrated to transport di and tripeptides that have L-AA, an acidic, or hydrophobic group at C-terminus, a weak basic group in α-position at N-terminus, a ketomethylene or acid amide bond, or a trans conformation of peptide bonds [94,95]. In addition to PEPT1, CaSR is involved in the peptide-induced GLP-1 secretion. CaSR is a G-protein-coupled receptor widely expressed in human tissues. In the intestine, CaSR is involved in nutrient sensing [96] and serves as a broad-spectrum of L-aromatic AA sensor [97]. Studies showed that L-phenylalanine and L-tryptophan induce GLP-1 secretion through the CaSR-mediated mechanism [98,99]. The activation of CaSR triggers depolarization of the cell membrane that leads to the opening of the voltage-gated Ca2+ channel and subsequent influx of Ca2+ [100]. However, not all AAs elicit Ca2+-mediated GLP-1 secretion via a CaSR-independent mechanism. L-glutamine was shown to induce [Ca2+]i increase and GLP-1 secretion via a cAMP signaling-mediated but PLC-independent pathway [80], indicating that this AA-induced GLP-1 secretion is not associated with the activation of CaSR [101]. Interestingly, it was reported that L-cells also uptake L-glutamine through one of the Na+-dependent AA transporters, SLC38A2 [102]. Intracellular glutamine then enhances glutamate dehydrogenase and ATP production via promoting the citric acid cycle [103], which results in the depolarization of the cell membrane, triggering GLP-1 secretion. L-ornithine, a non-proteinogenic AA, can trigger GLP-1 secretion via activating the GPRC6A/Gαq/PLC/Ca2+ pathway [101].
3. Polyphenols
Polyphenolic compounds are a group of secondary metabolites that are ubiquitously present in many plants and foods. The biosynthesis of most polyphenols in the plant originates from the phenylalanine through a shikimate/phenylpropanoid pathway. However, secoiridoid polyphenols, typically derived from Oleaceae family, are synthesized through a mevalonic acid pathway [104]. Based on their diverse chemical structures, polyphenols are classified into phenolic acids, flavonoids, stilbenes, and lignans [105], their represent chemical structures are shown in Figure 2 The major food or plant sources of various polyphenols are beyond the scope of this review, but the interested readers can refer to previous review articles on this topic [106,107]. Phenolic acids are a group of compounds with a phenolic ring and carboxylic acid. Hydroxycinnamic acids are the most common phenolic acids in fruits [108], which primarily consist of caffeic acid, chlorogenic acid, and ferulic acid. Flavonoids are the largest and widely distributed group of polyphenols. The main structure of flavonoids is diphenylpropanes, consisting of two aromatic rings (A and B) linked by a heterocyclic ring C [109]. Based on the differences in the structures of the heterocyclic ring, flavonoids are further divided into six groups: flavones, flavonols, flavanones, isoflavones, flavanols, and anthocyanins. Stilbenes are a relatively small group of plant metabolites synthesized through the phenylpropanoid pathway, and they contain a 1,2-diphenylethylene as the basic structure. Stilbenes are primarily found in grapes and some berries, and one of the well-known stilbenes in human diet is resveratrol, which is present in red wine, grape skin, and blueberry. Lignans are derived from phenylalanine via the dimerization of substituted cinnamic alcohols. Flaxseed and sesame seed are very rich sources of lignans with matairesinols and secoisolariresinols being the dominant forms [110,111].
Most dietary polyphenols exist naturally in the form of esters, glycosides, or polymers, many of which are poorly absorbed by the intestine. One of the most extensively studied categories of polyphenols, are flavonoids. A majority of flavonoids are non-covalently bound to glucose in plants, and some are present as flavonoid oligomers such as epicatechin in cocoa. The hydrolysis of the glycoside form of flavonoids begins in the mouth where flavonoids are partially released from the food matrix by saliva [112]. The low pH in the stomach facilitates the degradation of flavonoid oligomers into small units [113], but most flavonoids are hydrolyzed and further metabolized in the small intestine [114,115], where β-glucosidases cleave the glucose moiety from flavonoids and generate the aglycone form [116]. The hydrophobic flavonoids pass the apical membrane via passive diffusion, while other flavonoids in the glycosylated form can be transported into the enterocytes via SGLT1 [117]. Afterward, the absorbed flavonoids are conjugated with glucuronic acid by uridine-diphosphate-glucuronosyltransferase in the enterocytes. The degree of conjugation depends on the structure of the absorbed flavonoids. For instance, flavonoids containing a hydroxyl group on the B-ring are less predisposed to glucuronidation, but those containing catechol are more prone to glucuronidation [118]. Flavonoids that are absorbed from the small intestine are transported to the liver where they can be further conjugated with sulfate and/or methyl groups and then partially excreted through bile [119,120,121]. A large proportion of ingested flavonoids do not undergo hydrolysis or conjugation in the small intestine [122] and are not absorbed [121]. Instead, these unabsorbed flavonoids reach the colon where they are metabolized by colonic bacteria into smaller molecules such as phenolic acids, which can be partially absorbed into the circulation [123,124]. Specifically, some intestinal bacteria strains have the enzymatic activity of glucuronidase and glucosidase that can deconjugate the glucuronidated or glycosylated forms of flavonoids, and yield simple deconjugated products that are partially absorbed and metabolized by colonic enterocytes.
3.1. Polyphenols and GLP-1 Secretion
Emerging evidence shows that some polyphenols can induce GLP-1 secretion from L-cells via various mechanisms Specifically, polyphenols can function as ligands of GPCRs, or regulate intracellular signaling molecules to modulate GLP-1 secretion. Additionally, some polyphenols might indirectly cause GLP-1 secretion mediated by gut microbiota. The review below summarizes the studies in the area of polyphenols and modulation of GLP-1 as well as the potential underlying mechanisms for this effect.
3.1.1. Polyphenols as Activators of GPCRs
L-cells apparently could respond to a variety of structurally unrelated polyphenols via various mechanisms. Several polyphenolic compounds have been found to regulate GLP-1 secretion via GPCRs or cAMP signaling pathway, as summarized in Table 1. Specifically, GPCRs on the L-cells might be activated by polyphenols to regulate GLP-1 secretion. Curcumin, a polyphenolic turmeric, has been found to induce GLP-1 secretion in cultured L-cells with EC50 of 4.5 μM [125,126]. Oral intake of 1.5 mg/kg curcumin increased GSGS concomitant with improved glucose tolerance in rats [127]. It was further found that curcumin-induced GLP-1 secretion can be abrogated by the Ca2+ channel antagonist or IP3-receptor inhibitor, suggesting that both Ca2+ entry and release from intracellular store play a role in mediating curcumin-stimulated GLP-1 secretion, the latter of which may be due to the activation of PLC [127]. Interestingly, preincubation of GluTag L-cells with an antagonist (GW1100) of GPR40/120, both of which are coupled to Gαq to activate the IP3/Ca2+ signaling pathway, blunted curcumin-induced GLP-1 secretion [127], demonstrating that curcumin-induced GLP-1 secretion could be mediated via the activation of GPR40/120. Consistently, administration of the antagonist of GPR40/120 abolished curcumin-induced GLP-1 secretion in rats [127]. While it is still unclear which receptor plays the predominant role in curcumin on GLP-1 secretion, the reported results suggest that curcumin could be a ligand of GPR40/120. Delphinidin 3-rutinoside (D3R), one of the anthocyanin derivatives from berry, increased GLP-1 secretion from L-cells with the lowest effective concentration of 10 μM, via a GPR40/120-mediated mechanism [128]. Similarly, the oral intake of 5 mg/kg of anthocyanin-rich blackcurrant (Ribes nigrum L.) extract, which contains 19.3% D3R, 4.6% delphinnidin 3-glucoside, 19.4% cyanidin 3-rutinoside, and 2% cyanidin 3-glucoside, augmented GSGS and improved glucose tolerance in rats [129]. Further, diabetic mice fed a diet containing 11 g/kg anthocyanins-rich extract from blackcurrant (R. nigrum) for 7 weeks showed an improved plasma GLP-1 concentration, accompanied with reduced hyperglycemia [130]. In contrast to the findings in animal studies, intake of anthocyanins-rich blackcurrant juice, which contains 599 mg anthocyanins (260 mg D3R, 209 mg cyanidin-3-rutinoside, 76 mg delphinidin-3-glucoside and 33 mg Cyanidin-3-glucoside), failed to elevate postprandial plasma GLP-1 levels in healthy individuals but led to a slightly decrease on GLP-1 level at 90 min after intake of anthocyanins-rich blackcurrant juice [131]. While the reasons for these inconsistent results observed in rodents and human studies are unclear, the dose of anthocyanins that was used in rats and in diabetic mice is equivalent to 57 mg/day and 12 mg/day, respectively, which are much lower than the dose used in above human study, implying that the supraphysiological dose might alter the stimulatory effect of anthocyanins on GLP-1 secretion. Other than flavonoids, a natural phenolic acid, grifolic acid, activated GPR120 that led to GLP-1 secretion from cultured L-cell at the concentration of 30 μM [132]. In addition, the elevated [Ca2+]i was detected in grifolic acid-treated cells, suggesting that this compound induced GLP-1 secretion was mediated via GPR120/Ca2+ signaling. However, it is presently unknown whether grifolic acid stimulates GLP-1 secretion in vivo.
As GPR120/GPR40 are FA receptors, it is unclear how above discussed polyphenols are able to activate GPR120/GPR40. Interestingly, Cho et al. tested the effect of five different polyphenols (chrysin, pinocembrin, galangin, pinobanksin, and caffeic acid phenethyl ester) on GPR120 and found that flavanones (pinocembrin and pinobanksin) are more potent in activating GPR120 than that of tested flavones (chrysin and galangin) [133]. The only structure difference between these two subcategories of flavonoids is that flavones have a double bond between the C2 and C3 positions, whereas flavanones is saturated in C-ring, suggesting that the double bond between C2 and C3 might decrease the affinity of polyphenols for GPR120 136]. Indeed, curcumin and D3R, which activate GPR120 as aforementioned, also lack this double bond.
In addition to FA receptors, it was found that the superfamily of bitter taste receptors (Tas2Rs), which consist of 25 members in humans [134] and 35 members in rats and mice [135], are expressed in the intestinal enteroendocrine cells [136,137], as well as clonal L-cell lines [138]. Tas2Rs are GPCRs that detect a diversity of bitter compounds and transduce the signal through G-protein-coupled signaling. Like other GPCRs, activation of Tas2Rs increases GLP-1 secretion via the Gα-gustducin/Gβγ-triggered pathway, which can initiate IP3/Ca2+ signaling, subsequently leading to the opening of transient receptor potential (TRP) ion channels [139], and triggering Ca2+ influx and exocytosis of GLP-1 vesicle. Given that many dietary polyphenolic compounds are known to have a bitter taste, and they can interact with various taste receptors, in particular, Tas2Rs [140], it is possible that Tas2Rs mediate polyphenols-induced GLP-1 secretion. Tas2Rs can be activated by a wide variety of polyphenols, but it appears that different polyphenolic compounds interact with different or the same forms of TasRs depending on their chemical structures. For instance, Tas2R14, one of the human TasRs with a broad agonist spectrum [141], is activated by ester form of phenolic acids, such as ferulic acid ethyl ester [142], whereas anthocyanins bind to the human taste receptor subtypes Tas2R5 and Tas2R7, isoflavones and epigallocatechins have better affinity to Tas2R39 [143]. While polyphenols have been found to activate several Tas2Rs, studies on whether polyphenols stimulate GLP-1 secretion via Tas2Rs are limited. EGCG is one of the most effective Tas2R39 activator among polyphenolic compounds with an EC50 value of 8.5μM [142]. In addition, EGCG activated transient receptor potential ion channel A1 (TRPA1) in the mouse clonal L-cell line (STC-1) [144], which was opened after Tas2Rs activation to initiate the Ca2+ influx. EGCG has been reported to induce GLP-1 secretion in the mouse ileum segment [145], and oral intake of EGCG stimulated GLP-1 secretion in mice and diabetic patients [146,147], suggesting that EGCG-stimulated GLP-1 might be mediated via Tas2Rs. However, as EGCG activates multiple subtypes of Tas2Rs, such as Tas2R43 [142], which expresses on L-cells in both mice and humans [148], more studies are needed to determine which Tas2Rs are involved in the EGCG-induced GLP-1 secretion. Grape seed-derived proanthocyanidins (GSPE), which is composed of monomers or flavan-3-ols (21.3%), dimers (17.4%), trimers (16.3%), tetramers (13.3%) and oligomers (31.7%) of procyanidins, were found to increase GLP-1 secretion from clonal L-cells as well as mouse ileum and colon segments [149]. Consistent with this ex vivo finding, a single oral administration of 1 g/kg GSPE induced GLP-1 secretion in rats [149]. The stimulatory action of GSPE on GLP-1 secretion might result from a rise in intracellular Na+ influx via cation channel, TRPM5, which leads to the depolarization and the opening of voltage-gated calcium channel, resulting in an increase in Ca2+ influx [150], an essential step for exocytosis of GLP-1 granules. It is noted that TRPM5 is the common downstream molecule on the taste receptor signaling pathway, suggesting that the GSPE-induced GLP-1 release might depend on the taste receptor. As condensed tannins are ligands of Tas2Rs [142], proanthocyanidins might also activate taste receptors to regulate GLP-1 secretion. However, the degree of polymerization of proanthocyanidins might affect their affinity for taste receptors [143], for example, procyanidin trimer C2 activates Tas2R5, but procyanidin dimer B2 fails to activate any Tas2Rs. Thus, studies on specific proanthocyanidin from GSPE should be performed to verify the activity of certain proanthocyanidin on Tas2Rs.
GPCRs play a critical role in neuronal, hormonal, and nutritional regulation of GLP-1 secretion [151]. While polyphenols have been reported to activate multiple types of GPCRs [152], the most abundant GPCRs that expressed on enteroendocrine L-cells are FA receptors and taste receptors [153]. Therefore, efforts identifying the GPCRs that mediate polyphenols-induced GLP-1 secretion are primarily focused on FA receptors and taste receptors. As one polyphenol might be able to activate multiple receptors, in particular Tas2Rs, and most Tas2Rs share the same intracellular signaling to regulate GLP-1 secretion, the results from pharmaceutical inhibition of specific Tas2R could be misleading and thus should be carefully interpreted. In addition, most of these findings were obtained using cultured cells and tissues, and their physiological relevance as well as the most effective doses needed to be confirmed in vivo, given that ingested polyphenols can be metabolized and their actions can be affected by interacting with proteins in the GI tract.
3.1.2. Polyphenols as Regulators of cAMP Signaling
It is well established that the elevation of intracellular cAMP levels induces both transcription and secretion of GLP-1 via protein kinase A (PKA) and Epac, a cAMP-activated effector protein [154,155]. Some polyphenols may promote GLP-1 secretion via activating cAMP-mediated intracellular signaling. Genistein, an isoflavone present in legumes and Chinese herbal medicines Genista tinctoria Linn and Sophora subprostrala Chun (Fabaceae family), is a popularly used supplement for various presumably beneficial effects. Indeed, it was shown that dietary intake of genistein or soy products lowered plasma glucose in diabetic animals [156,157] and in humans [158,159]. At physiologically achievable concentrations, genistein can activate cAMP signaling to induce insulin secretion from pancreatic β-cells [160]. In diabetic mouse models, dietary provision of genistein increased plasma insulin levels, and mitigated diabetes [161,162,163,164]. Interestingly, a recent study indicated that genistein enhanced GLP-1 secretion in alloxan-induced insulin deficient diabetic rats [165]. While this is a notable finding with respect of genistein effect, it is actually uncertain whether increased circulating GLP-1 levels as observed in genistein-treated diabetic animals were due to directly enhanced GLP-1 secretion, and whether cAMP signaling is involved in this genistein action.
Recently, it was found that hispidulin, a flavone found in Petasites, Artemisia, and Salvia, dose-dependently stimulates GLP-1 secretion in GluTag L-cells and primary mouse ileum crypts, with as low as 1 µM eliciting a significant effect [166]. In addition, oral administration of hispidulin significantly exerted antidiabetic effect concomitant with improved non-fasting insulin concentrations and β-cell survival in diabetic mice, consistent with well-established metabolic effects of GLP-1. It was further shown that hispidulin-induced GLP-1 secretion was mediated via increased intracellular cAMP content, likely as a result of the inhibition of phosphodiesterase (PDE), the enzyme that degrades cAMP into AMP [166]. However, it is unclear how hispidulin inhibits PDE in L-cells. Interestingly, this study further found that apigenin and luteolin, two other flavones, also increased GLP-1 secretion, whereas no other subclasses of tested flavonoids were able to significantly induce GLP-1 secretion from L-cells, suggesting that flavones, but not other subtypes of flavonoids, may be GLP-1 agonists. While the specific chemical structure on hispidulin or other flavones responsible for its stimulatory action on GLP-1 secretion is presently unclear, flavones have a double bond between C2 and C3 of the C-ring in the flavonoid skeleton, and an oxidized group at the C4 position of its B-ring, which may be crucial for this unique effect of flavone on GLP-1 secretion. Nevertheless, this research may have revealed a new mechanism underlying the antidiabetic effects of some flavones as observed in animal models [167,168].
Silymarin, a flavonolignan isolated from milk thistle (Silybum marianum), can stimulate GLP-1 secretion in obese rats [169]. While the mechanism underlying silymarin-induced GLP-1 secretion is unknown, it was found that this compound stimulates insulin secretion by inhibiting PDE activity in insulin secreting cell line [170], indicating that silymarin might have the similar effect in L-cells.
Coffee-derived polyphenols were also shown to stimulate GLP-1 secretion from human clonal L-cell in a dose-dependent manner, which was mediated via elevated intracellular cAMP signaling [36]. In line with this in vitro result, oral administration of coffee polyphenols to mice induced moderate but significant increase in GSGS [36]. Notably, a single intake of coffee-extract polyphenols, which mainly contain caffeoylquinic acids and chlorogenic acids, increased postprandial GLP-1 levels in healthy male adults [171]. Consistently, a large population cohort study showed that high coffee consumption was associated with low risks of T2D [172]. Caffeoylquinic acid and caffeic acid, two main polyphenols from coffee extract, have been found to inhibit PDE activity with IC50 of 0.49 and 0.48 mM, respectively [173], which might ascribe to the increased GLP-1 secretion following administration of coffee polyphenols.
3.2. The Potential Role of Microbiota in Polyphenol-Stimulated GLP-1 Secretion
In addition to directly targeting L-cells to stimulate GLP-1 secretion, polyphenols might also modulate GLP-1 secretion indirectly involving intestinal microbiota. The human gut hosts a vast diversity of microorganisms that have been the subject of extensive research in recent years, as they may play important roles in human physiology, metabolism, and disease. Before weaning, the composition of intestinal microbiota is simple, relatively unstable, and undergoes drastic changes [174], whereas the composition of gut microbiota in healthy adults is substantially stable over time [175,176]. However, dietary composition can profoundly influence the gut microbiota composition in the host [177,178,179]. As to nutrient metabolism, gut microbiota metabolizes non-digestible nutrients into SCFAs and gas. It was found that dietary intake of resistant starch, which escapes digestion in the small intestine, increased the proportion of butyrate-producing bacterium Eubacterium hallii [177], and sequentially increased butyrate production [180], suggesting that the shift in dietary pattern can modulate the abundance of SCFA-producing bacteria.
A large portion of the ingested polyphenols enters the large intestine where non-absorbed polyphenols are fermented by various colonic microbiota to generate a wide range of metabolites because of the large variability of their chemical structures [181]. For example, the flavonol, myricetin and quercetin, are structurally similar, and the only difference between them is that myricetin has three hydroxyl groups on the C ring while there are two for quercetin. Metabolism of quercetin by microbiota produces 2-(3,4-dihydroxyphenyl)-acetic acid and protocatechuic acid, but breakdown of myricetin generates 2-(3,5-dihydroxyphenyl)-acetic acid and gallic acid [182,183]. Resveratrol, a stilbenoid with two benzene rings, can be fermented by Slackia equolifaciens and Adlercreutzia equolifaciens to produce lunularin [184]. On the other hand, polyphenols can modulate the relative abundance of the gut microbiota. Some polyphenols have antibiotic activity by suppressing the growth of pathological bacteria. For instance, when intestinal bacteria were cultured with polyphenolic compounds from tea extracts, the growth of pathological bacteria, Clostridium perfringens, Clostridium difficile, and Bacteroides spp was inhibited, but the abundance of the probiotic bacteria was not affected [185]. In addition, some polyphenols can promote the growth of beneficial bacteria while inhibiting the growth of pathogens, therefore exhibiting a prebiotic-like effect. The polyphenols extracted from concord grapes and fermented green tea were shown to reduce high-fat-diet (HFD)-induced increase in the ratio of Firmicutes/Bacteroidetes and protect the intestinal barrier function in rodents [186,187]. Grape polyphenols, which primarily include anthocyanins, flavanols, flavonols, stilbenes and phenolic acids [188], enhanced the growth of Akkermansia (A) muciniphila in HFD-fed mice, which is an important species for degrading mucin [186].
As polyphenols modulate microbial abundance, they might also alter the production of microbial metabolites. As aforementioned, SCFAs are the main microbiota metabolites of non-digestible food components, and SCFAs, primarily acetate, propionate, and butyrate, can trigger GLP-1 secretion in vitro and in vivo via their GPCRs [74,87]. Clostridial clusters IV and XIVa, which belong to the Firmicutes phylum, are the two most important butyrate producers [189]. Among these two taxa, Faecalibacterium (F) prausnitzii and Eubacterium (E) rectale are the two most dominant bacterial strains in the human colon, and they play a major role in producing butyrate [190]. F. prausnitzii possesses butyryl coenzyme A (CoA): acetate CoA transferase and acetate kinase activities, and it converts two molecules of acetyl-CoA to butyrate [191], whereas E. rectale catalyzes butyrate formation from lactate and acetate [192]. In addition to butyrate-producing bacteria, there are bacteria species that produce other SCFAs, such as bifidobacterium species (Phylum Actinobacteria), which ferment carbohydrates to generate acetate [193], and A. muciniphila (Phylum Verrucomicrobia) that produces acetate and propionate [194,195]. Dietary polyphenols have been shown to enhance the population of SCFA-producing bacteria in fecal microbiota from humans [196] (Table 2). For instance, exposure of human fecal bacteria to 150 mg/L catechins enhanced the growth of E. rectale, the butyrate-producing bacteria, but this effect was not observed until after 17 h of incubation period [196]. As the regulation of bacterial growth by polyphenols might be mediated by their metabolites, it is possible that a longer incubation time may be needed for this strain of bacteria to generate adequate amounts of metabolites necessary for stimulating bacteria growth. Similar to this finding, in another study, the incubation of human fecal bacteria with 10 μg/mL chlorogenic acid for 48 h increased the population of bifidobacterium species [197]. In addition, several dietary intervention studies in healthy individuals or in obese subjects showed that polyphenols enhanced the population of SCFA-producing bacteria. In this regard, it was shown that oral intake of pomegranate polyphenols (1000 mg/day for 4 weeks), which is a rich source of tannins, anthocyanins, and non-colored flavonoids, as well as lignans [198], increased the population of A. muciniphila in the fecal samples of healthy subjects [199]. Daily intake of 272 mL red wine (equivalent to 798 mg phenols, including 104.68 mg flavan-3-ols, 26.44 mg anthocyanins, and 21.49 mg hydroxybenzoic acids) for 15 days increased the proportion of F. prausnitzii in feces of obese [200], but did not alter its profile in healthy subjects [201], suggesting that grape phenolic compounds may exerts beneficial effects in humans via modulating gut bacteria composition. Relative to limited studies in humans, more research on the effect of polyphenols on gut bacteria has been done in rodent models. It was shown that feeding mice HFD containing 0.1% oolong tea polyphenols increased the proportion of F. prausnitzii and other SCFA-producing bacteria, concomitant with increase in the fecal contents of SCFAs in mice [202]. Similarly, oral intake of mango (Mangifera indicaL. cv. “Keitt”) polyphenol extracts for 9 weeks increased the butyrate-producing bacteria, Clostridium butyrium, and improved butyrate production by 150% in rats [203]. However, some studies reported inhibitory or no effect of polyphenols on the gut bacteria. Daily intake of 10 g/kg black tea or green tea polyphenols-containing diets, which contain 9.3% and 65.9% catechins, respectively, for 3 weeks reduced the abundance of Clostridium XIVa (butyrate producer) in the feces of rats, but had no significant effect on the cecal SCFA contents [204]. Dietary provision of 0.3% EGCG, one of the key polyphenols of green tea extract, reduced the abundance of fecal Clostridial IV and XIVa bacteria in rats, but had no effect on cecal SCFAs [205]. Most of the polyphenol extracts from beverages and fruits in the above-mentioned studies differ in polyphenol contents, compositions of polyphenolic compounds, and purity, which make it difficult to compare the effect of these polyphenol extracts on microbiota or microbial products. Furthermore, fecal samples are often used for analysis of gut microbiota population, but both cecal and fecal samples have been used for the analysis of SCFA production. However, the population of microbiota from cecum and feces are different, in particular, the Clostridium group, which has a lower proportion in cecal microbiota than in fecal microbiota [206]. Therefore, it is possible that the difference in the microbiota population is responsible for the observed difference in SCFA contents between feces and cecum in the same subject. In this regard, comparison between studies using different sample origins might lead to inconsistent outcomes.
3.3. Polyphenols and DPP-IV
In addition to directly stimulating GLP-1 secretion, some polyphenols may suppress DPP-IV activity [207] (Table 3), thus potentially increasing the half-life of GLP-1 in the circulation. For example, it was shown that bis-pyrano prenyl isoflavone isolated from leaves of P. molluginifolia inhibited 53% DPP-IV activity at 100 µM in a cell-free assay [208]. In addition, rutin, eriocitrin, eriodictyol, naringenin, naringin, hesperidin, and hesperetin exhibited inhibitory effects on DPP-IV activity in the cell-free assay, with the IC50 of 0.485, 5.44, 3.91, 5.5, 3.82, 5.7, and 5.7 mM, respectively [209]. Interestingly, the potency of polyphenols on inhibiting DPP-IV could vary between the species from which the enzyme is derived. For example, apigenin exhibited potent inhibition of porcine kidney DPP-IV with an IC50 at only 0.14 ± 0.02 μM [207], but it is much less potent in inhibiting recombinant human DPP-IV, with a 200 μM concentration only ablating the enzyme activity by 44% [210]. Although the exact reason for this difference is not clear, it could be due to the difference in DPP-IV amino acid sequence between different mammalian species. While the amino acid sequences of DPP-IV are highly conserved, porcine DPP-IV only shares 88% identity in its amino acid sequence with that of human DPP-IV [211]. Given this possibility, the species of DPP-IV should be considered in determining the effects of polyphenolic compounds on its activity. Physiologically, there are two forms of DPP-IV, a soluble form in the blood and a membrane-anchored form on the luminal surface of the vascular endothelium. Both forms of DPP-IV have identical catalytic region, but the soluble form of DPP-IV lacks the cytoplasmic and transmembrane domains [212]. Therefore, polyphenolic compounds could have different interaction modes with two forms of DPP-IV that may result in different inhibitory efficacy. A study showed that a single dose of 1 g/kg of GSPE via oral gavage reduced intestinal DPP-IV activity (34.4%), whereas it did not inhibit plasma DPP-IV activity [213]. It is possible that the relatively low inhibitory effect on the circulating DPP-IV could be due to the low bioavailability of polyphenolic compounds. It may also be caused by the fact that the majority of polyphenols and their metabolites (over 80%) in the blood are already associated with proteins, primarily albumin [210], which may prevent their further interaction with DPP-IV. Interestingly, a study showed that GSPE effectively inhibited the activity of recombinant human DPP-IV in a cell-free system, achieving 70% inhibition at the concentration of 200 mg/L, but this dose of GSPE only exerted a weak inhibitory action on the activity of DPP-IV that was isolated from human plasma [213]. Therefore, DPP-IV proteins from the blood should be used to better resemble the physiological environment for assessing the inhibitory action of polyphenols in vivo.
In addition to the sources of DPP-IV, the substrates used in DPP-IV activity assay could confound interpretation of the results. Three different substrates are used in the DPP-IV assay, Gly-Pro-p-nitroanilide hydrochloride in colorimetric assay, Gly-Pro-7-amido-4-methylcoumarin hydrobromide (GP-AMC) in fluorometric assay, and Gly-Pro-aminoluciferin in luminescent assay. The activity of DPP-IV was 10-fold higher using the fluorometric substrate compared to the colorimetric substrate, and it is even higher using luminescent substrate as compared to the fluorometric substrate [214]. For example, it was shown that flavone apigenin at 200 μM only inhibited DPP-IV activity by 20% using a fluorometric assay [210], but its value of IC50 for inhibiting DPP-IV was 0.14 μM using the luminescent substrate [207]. While the reasons for the discrepancy between these results are unclear, there is possibility that polyphenols may interact with colorigenic, fluorogenic, and/or luminescent substrates to alter the assay sensitivity. Indeed, it was shown that some flavonoids (cyanidin, pelargonidin, catechin, myrecetin and kaempferol) bind to proline residue [215], which could modulate the availability of these compounds for interaction with DPP-IV. Interestingly, a more recent study showed that coumarin, a component of GP-AMC, inhibits DPP-IV activity [216]. However, it is unknown whether GP-AMC also has ability to inhibit DPP-IV. Therefore, in determining the inhibitory effect of polyphenolic compounds on DPP-IV, the type of assay methods, the sources and species of the enzymes, and inclusion of positive control should all be carefully considered.
It appears that DPP-IV can be inhibited by a great number of structurally diverse compounds, ranging from a simple phenolic compound, such as gallic acid, to polyphenolic flavonoid glycosides, such as eriocitrin [207]. DPP-IV has three major active sites (S1, S2, and S3) for its enzymatic activity, including the side chains of a catalytic triad (Ser630, Asn710, and His740) in the S1, cavity near Glu205, Glu206, and Tyr662 in S2, and Ser209, Arg358, and Phe357 in S3 [217]. The inhibitory action of polyphenolic compounds in DPP-IV might be due to the formation of hydrogen bonds between the amino acid residues in the catalytically active sites of DPP-IV and the hydroxyl groups of polyphenols. Indeed, in vitro assay showed that resveratrol (100 µM) has an inhibitory effect on DPP-IV activity [218] and molecular docking analysis demonstrated that resveratrol can form a hydrogen bond between its hydroxyl groups and the hydroxyl group of the side chain of DPP-IV at Ser630 (S1) and Ser209 (S3) [207]. In addition, resveratrol may also partially inhibit DPP-IV activity by forming electrostatic interactions with DPP-IV active sites Glu205 and Glu206 (S2) [207]. Based on computer-based molecular docking study, flavonoids are also potential DPP-IV inhibitors. The common characteristics of binding mode between flavonoids such as luteolin and apigenin and DPP-IV include the π-interaction between the benzene ring of flavonoids and the benzene amino acids at the active sites of DPP-IV and a hydrogen bond between the oxygen on the C ring of flavonoids and the ammonium of a guanidine group of Arg358 in the active site S3 of DPP-IV [207]. It was found that chrysin, a flavone glycoside, can inhibit DPP-IV due to the π-interaction between the benzene ring of chrysin and the S1 pocket (Tyr547) of the enzyme [219]. However, the results of molecular docking analyses from different research groups were not all consistent. Fan et al. showed that quercetin and genistein suppress DPP-IV activity by forming hydrogen bond [207], but Srivastava et al. found that quercetin and genistein have no effect on DPP-IV activity [220]. The inconsistency in the findings might be caused by using various crystal structures of DPP-IV. Indeed, Fan et al. and Srivastava et al. used different DPP-IV structures for molecular docking analysis. In addition, the diverse algorithm parameters in docking software and the lack of the validation of prediction performance could lead to the inconsistent results. Thus, validation of the results from molecular docking analysis using enzyme and cells are essential for confirming the efficacy of polyphenols in inhibiting DPP-IV.
4. Conclusions
Loss of functional β-cell mass plays a central role in the deterioration of blood glucose control in T2D. The GLP-1 signaling pathway has been extensively explored for developing incretin-based therapies for T2D, as its activation can promote GSIS and β-cell mass. Polyphenols are a group of plant-derived compounds that may exert beneficial effects on human health. Emerging evidence shows that some phenolic compounds can induce GLP-1 secretion from intestinal L-cells, and thus may be helpful in improving glucose homeostasis. However, this is still largely an unexplored territory and there remain many interesting questions that need to be addressed. It is unclear which category of phenolic compounds in the diet is superior to the others in stimulating GLP-1 secretion. Therefore, studies should be carried out to compare the efficacy of different categories of polyphenolic compounds, which may provide a better understanding of the chemical structure-function relationship, and thus shed light on how a particular polyphenolic compound activates GLP-1 secretion. Given that most of the non-absorbed polyphenols were fermented by gut microbiota and some products derived from fermentation was found to induce GLP-1 secretion, it is appealing to investigate whether and how the gut bacteria play a role in mediating polyphenol regulation of GLP-1 secretion.
Author Contributions
Y.W. wrote the draft of manuscript under the supervision of D.L.; D.L. and H.A. revised the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work is partially supported by a grant from Diabetes Action Research and Education Foundation (USA) to D. Liu.
Acknowledgments
This paper is partially supported by Virginia Tech Open Access Subvention Fund.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AA | amino acid |
| cAMP | cyclic adenosine monophosphate |
| CaSR | calcium-sensing receptor |
| CoA | coenzyme A |
| CQA | caffeoylquinic acid |
| D3R | delphinidin 3-rutinoside |
| DAG | diacylglycerol |
| DPP-IV | dipeptidyl-peptidase-IV |
| EGCG | epigallocatechin gallate |
| ERK1/2 | extracellular signal-related kinase 1/2 |
| GIP | glucose-dependent insulinotropic peptide |
| GLP-1 | glucagon-like peptide 1 |
| GLP-1R | GLP-1 receptor |
| GPCR | G-protein coupled receptor |
| GP-AMC | Gly-Pro-7-amido-4-methylcoumarin hydrobromide |
| GSGS | glucose stimulated GLP-1 secretion |
| GSIS | glucose stimulated insulin secretion |
| GSPE | Grape seed-derived proanthocyanidins |
| FA | Fatty acid |
| FFAR | FA receptor |
| IP3 | inositol trisphosphate |
| PDE | phosphodiesterase |
| PEPT | peptide transporter |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PLC | phospholipase C |
| PK | protein kinase |
| PUFA | Polyunsaturated fatty acid |
| PYY | peptide tyrosine tyrosine |
| SCFA | short-chain FA |
| SGLT | sodium-dependent glucose transporter |
| T2D | type 2 diabetes |
| Tas1R | sweet taste receptor 1 |
| Tas2R | sweet taste receptor 2 |
| TRP | transient receptor potential |
| TRPA1 | transient receptor potential ion channel A1 |
| TRPM5 | transient receptor potential channel M5 |
References
- Roglic, G. Global Report on Diabetes; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- CDC. National Diabetes Statistics Report 2020; Centers for Disease Control Prevention: Atlanta, GA, USA, 2020.
- Kumanyika, S.; Jeffery, R.W.; Morabia, A.; Ritenbaugh, C.; Antipatis, V.J. Public Health Approaches to the Prevention of Obesity Working Group of the International Obesity Task, F., Obesity prevention: The case for action. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Kubota, N.; Kadowaki, T. Imbalanced Insulin Actions in Obesity and Type 2 Diabetes: Key Mouse Models of Insulin Signaling Pathway. Cell Metab. 2017, 25, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E. The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of Type 2 diabetes. Diabetologia 2003, 46, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Holst, J.J.; Gribble, F.; Horowitz, M.; Rayner, C.K. Roles of the Gut in Glucose Homeostasis. Diabetes Care 2016, 39, 884–892. [Google Scholar] [CrossRef] [Green Version]
- Tourrel, C.; Bailbé, D.; Meile, M.-J.; Kergoat, M.; Portha, B. Glucagon-like peptide-1 and exendin-4 stimulate β-cell neogenesis in streptozotocin-treated newborn rats resulting in persistently improved glucose homeostasis at adult age. Diabetes 2001, 50, 1562–1570. [Google Scholar] [CrossRef]
- Holst, J.J.; Deacon, C.F.; Vilsboll, T.; Krarup, T.; Madsbad, S. Glucagon-like peptide-1, glucose homeostasis and diabetes. Trends Mol. Med. 2008, 14, 161–168. [Google Scholar]
- Drucker, D.J. The role of gut hormones in glucose homeostasis. J. Clin. Investig. 2007, 117, 24–32. [Google Scholar] [CrossRef]
- Holst, J.J.; Orskov, C. The incretin approach for diabetes treatment: Modulation of islet hormone release by GLP-1 agonism. Diabetes 2004, 53 (Suppl. 3), S197–S204. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Aulinger, B.; D’Alessio, D.A. Effect of glycemia on plasma incretins and the incretin effect during oral glucose tolerance test. Diabetes 2012, 61, 2728–2733. [Google Scholar] [PubMed] [Green Version]
- Lim, G.E.; Brubaker, P.L. Glucagon-like peptide 1 secretion by the L-cell: The view from within. Diabetes 2006, 55 (Suppl. 2), S70–S77. [Google Scholar] [CrossRef] [Green Version]
- Thorens, B. Expression cloning of the pancreatic beta cell receptor for the gluco-incretin hormone glucagon-like peptide 1. Proc. Natl. Acad. Sci. USA 1992, 89, 8641–8645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, W.; Siegel, E.; Creutzfeldt, W. Glucagon-like peptide-1 but not glucagon-like peptide-2 stimulates insulin release from isolated rat pancreatic islets. Diabetologia 1985, 28, 704–707. [Google Scholar] [PubMed]
- Liu, Z.; Habener, J.F. Glucagon-like peptide-1 activation of TCF7L2-dependent Wnt signaling enhances pancreatic beta cell proliferation. J. Biol. Chem. 2008, 283, 8723–8735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spreckley, E.; Murphy, K.G. The L-Cell in Nutritional Sensing and the Regulation of Appetite. Front. Nutr. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Näslund, E.; Barkeling, B.; King, N.; Gutniak, M.; Blundell, J.; Holst, J.; Rössner, S.; Hellström, P. Energy intake and appetite are suppressed by glucagon-like peptide-1 (GLP-1) in obese men. Int. J. Obes. 1999, 23, 304. [Google Scholar]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Kho, Z.Y.; Lal, S.K. The human gut microbiome—A potential controller of wellness and disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.I.; Zaid, M.A.; Anis, R.; Mishra, N. Protective role of tea catechins against oxidation-induced damage of type 2 diabetic erythrocytes. Clin. Exp. Pharmacol. Physiol. 2005, 32, 70–75. [Google Scholar] [CrossRef]
- Dragan, S.; Andrica, F.; Serban, M.C.; Timar, R. Polyphenols-rich natural products for treatment of diabetes. Curr. Med. Chem. 2015, 22, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Natural Polyphenols and Diabetes: Understanding their Mechanism of Action. Curr. Med. Chem. 2015, 22, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Zunino, S.J.; Storms, D.H.; Stephensen, C.B. Diets rich in polyphenols and vitamin A inhibit the development of type I autoimmune diabetes in nonobese diabetic mice. J. Nutr. 2007, 137, 1216–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, G.; Stepaniak, U.; Micek, A.; Kozela, M.; Stefler, D.; Bobak, M.; Pajak, A. Dietary polyphenol intake and risk of type 2 diabetes in the Polish arm of the Health, Alcohol and Psychosocial factors in Eastern Europe (HAPIEE) study. Br. J. Nutr. 2017, 118, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, T.; Yamada, M.; Miura, T.; Nagashima, K.; Ogura, K.; Inagaki, N.; Maeda-Yamamoto, M. Chronic administration of apple polyphenols ameliorates hyperglycaemia in high-normal and borderline subjects: A randomised, placebo-controlled trial. Diabetes Res. Clin. Pract. 2017, 129, 43–51. [Google Scholar] [CrossRef]
- Zygmunt, K.; Faubert, B.; MacNeil, J.; Tsiani, E. Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem. Biophys. Res. Commun. 2010, 398, 178–183. [Google Scholar] [CrossRef]
- Li, J.M.; Che, C.T.; Lau, C.B.; Leung, P.S.; Cheng, C.H. Inhibition of intestinal and renal Na+-glucose cotransporter by naringenin. Int. J. Biochem. Cell Biol. 2006, 38, 985–995. [Google Scholar] [CrossRef]
- Eid, H.M.; Martineau, L.C.; Saleem, A.; Muhammad, A.; Vallerand, D.; Benhaddou-Andaloussi, A.; Nistor, L.; Afshar, A.; Arnason, J.T.; Haddad, P.S. Stimulation of AMP-activated protein kinase and enhancement of basal glucose uptake in muscle cells by quercetin and quercetin glycosides, active principles of the antidiabetic medicinal plant Vaccinium vitis-idaea. Mol. Nutr. Food Res. 2010, 54, 991–1003. [Google Scholar]
- Strobel, P.; Allard, C.; Perez-Acle, T.; Calderon, R.; Aldunate, R.; Leighton, F. Myricetin, quercetin and catechin-gallate inhibit glucose uptake in isolated rat adipocytes. Biochem. J. 2005, 386 Pt 3, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Johnston, K.; Sharp, P.; Clifford, M.; Morgan, L. Dietary polyphenols decrease glucose uptake by human intestinal Caco-2 cells. FEBS Lett. 2005, 579, 1653–1657. [Google Scholar]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Barnes, S.; Prasain, J.; D’Alessandro, T.; Arabshahi, A.; Botting, N.; Lila, M.A.; Jackson, G.; Janle, E.M.; Weaver, C.M. The metabolism and analysis of isoflavones and other dietary polyphenols in foods and biological systems. Food Funct. 2011, 2, 235–244. [Google Scholar]
- Heijboer, A.C.; Frans, A.; Lomecky, M.; Blankenstein, M.A. Analysis of glucagon-like peptide 1; what to measure? Clin. Chim. Acta 2011, 412, 1191–1194. [Google Scholar] [CrossRef]
- Fujii, Y.; Osaki, N.; Hase, T.; Shimotoyodome, A. Ingestion of coffee polyphenols increases postprandial release of the active glucagon-like peptide-1 (GLP-1 (7–36)) amide in C57BL/6J mice. J. Nutr. Sci. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- McCarty, M.F. A chlorogenic acid-induced increase in GLP-1 production may mediate the impact of heavy coffee consumption on diabetes risk. Med. Hypotheses 2005, 64, 848–853. [Google Scholar] [CrossRef]
- Nakajima, K.; Oiso, S.; Kariyazono, H. Inhibitory Effect of (−)-Epigallocatechin-3-O-gallate on Octanoylated Ghrelin Levels in Vitro and in Vivo. Biol. Pharm. Bull. 2018, 41, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Kao, Y.H.; Hiipakka, R.A.; Liao, S. Modulation of endocrine systems and food intake by green tea epigallocatechin gallate. Endocrinology 2000, 141, 980–987. [Google Scholar] [CrossRef]
- Zanzer, Y.C.; Plaza, M.; Dougkas, A.; Turner, C.; Björck, I.; Östman, E. Polyphenol-rich spice-based beverages modulated postprandial early glycaemia, appetite and pyy after breakfast challenge in healthy subjects: A randomized, single blind, crossover study. J. Funct. Foods 2017, 35, 574–583. [Google Scholar] [CrossRef]
- Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Murphy, K.G.; Bloom, S.R. Gut hormones and the regulation of energy homeostasis. Nature 2006, 444, 854–859. [Google Scholar] [CrossRef]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Gunawardene, A.R.; Corfe, B.M.; Staton, C.A. Classification and functions of enteroendocrine cells of the lower gastrointestinal tract. Int. J. Exp. Pathol. 2011, 92, 219–231. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, A.; Dey, A.; Norrbom, C.; Carroll, R.; Zhang, C.; Laurent, V.; Lindberg, I.; Ugleholdt, R.; Holst, J.J.; et al. Disruption of PC1/3 expression in mice causes dwarfism and multiple neuroendocrine peptide processing defects. Proc. Natl. Acad. Sci. USA 2002, 99, 10293–10298. [Google Scholar] [CrossRef] [Green Version]
- Vahl, T.P.; Paty, B.W.; Fuller, B.D.; Prigeon, R.L.; D’alessio, D.A. Effects of GLP-1-(7–36) NH2, GLP-1-(7–37), and GLP-1-(9–36) NH2 on intravenous glucose tolerance and glucose-induced insulin secretion in healthy humans. J. Clin. Endocrinol. Metab. 2003, 88, 1772–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissele, R.; Goke, R.; Willemer, S.; Harthus, H.P.; Vermeer, H.; Arnold, R.; Goke, B. Glucagon-like peptide-1 cells in the gastrointestinal tract and pancreas of rat, pig and man. Eur. J. Clin. Investig. 1992, 22, 283–291. [Google Scholar] [CrossRef]
- MacDonald, P.E.; El-Kholy, W.; Riedel, M.J.; Salapatek, A.M.; Light, P.E.; Wheeler, M.B. The multiple actions of GLP-1 on the process of glucose-stimulated insulin secretion. Diabetes 2002, 51 (Suppl. 3), S434–S442. [Google Scholar] [CrossRef] [Green Version]
- Buteau, J. GLP-1 receptor signaling: Effects on pancreatic beta-cell proliferation and survival. Diabetes Metab. 2008, 34 (Suppl. 2), S73–S77. [Google Scholar] [CrossRef]
- Rowlands, J.; Heng, J.; Newsholme, P.; Carlessi, R. Pleiotropic Effects of GLP-1 and Analogs on Cell Signaling, Metabolism, and Function. Front. Endocrinol. 2018, 9, 672. [Google Scholar] [CrossRef] [Green Version]
- Bifari, F.; Manfrini, R.; Dei Cas, M.; Berra, C.; Siano, M.; Zuin, M.; Paroni, R.; Folli, F. Multiple target tissue effects of GLP-1 analogues on non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). Pharmacol. Res. 2018, 137, 219–229. [Google Scholar] [CrossRef]
- Ranganath, L.R.; Beety, J.M.; Morgan, L.M.; Wright, J.W.; Howland, R.; Marks, V. Attenuated GLP-1 secretion in obesity: Cause or consequence? Gut 1996, 38, 916–919. [Google Scholar]
- Mannucci, E.; Ognibene, A.; Cremasco, F.; Bardini, G.; Mencucci, A.; Pierazzuoli, E.; Ciani, S.; Fanelli, A.; Messeri, G.; Rotella, C.M. Glucagon-Like peptide (GLP)-1 and leptin concentrations in obese patients with Type 2 diabetes mellitus. Diabet. Med. 2000, 17, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Lastya, A.; Saraswati, M.R.; Suastika, K. The low level of glucagon-like peptide-1 (glp-1) is a risk factor of type 2 diabetes mellitus. BMC Res. Notes 2014, 7, 849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faerch, K.; Torekov, S.S.; Vistisen, D.; Johansen, N.B.; Witte, D.R.; Jonsson, A.; Pedersen, O.; Hansen, T.; Lauritzen, T.; Sandbaek, A.; et al. GLP-1 Response to Oral Glucose Is Reduced in Prediabetes, Screen-Detected Type 2 Diabetes, and Obesity and Influenced by Sex: The ADDITION-PRO Study. Diabetes 2015, 64, 2513–2525. [Google Scholar] [CrossRef] [Green Version]
- Calanna, S.; Christensen, M.; Holst, J.J.; Laferrere, B.; Gluud, L.L.; Vilsboll, T.; Knop, F.K. Secretion of glucagon-like peptide-1 in patients with type 2 diabetes mellitus: Systematic review and meta-analyses of clinical studies. Diabetologia 2013, 56, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Vella, A.; Cobelli, C. Defective Glucagon-Like Peptide 1 Secretion in Prediabetes and Type 2 Diabetes Is Influenced by Weight and Sex. Chicken, Egg, or None of the Above? Diabetes 2015, 64, 2324–2325. [Google Scholar] [CrossRef] [Green Version]
- Toft-Nielsen, M.B.; Madsbad, S.; Holst, J.J. Continuous subcutaneous infusion of glucagon-like peptide 1 lowers plasma glucose and reduces appetite in type 2 diabetic patients. Diabetes Care 1999, 22, 1137–1143. [Google Scholar] [CrossRef]
- Bond, A. Exenatide (Byetta) as a Novel Treatment Option for Type 2 Diabetes Mellitus; Baylor University Medical Center Proceedings; Taylor & Francis: Abingdon, UK, 2006; pp. 281–284. [Google Scholar]
- Buse, J.B.; Rosenstock, J.; Sesti, G.; Schmidt, W.E.; Montanya, E.; Brett, J.H.; Zychma, M.; Blonde, L.; Group, L.-S. Liraglutide once a day versus exenatide twice a day for type 2 diabetes: A 26-week randomised, parallel-group, multinational, open-label trial (LEAD-6). Lancet 2009, 374, 39–47. [Google Scholar] [CrossRef]
- Rask, E.; Olsson, T.; Soderberg, S.; Johnson, O.; Seckl, J.; Holst, J.J.; Ahren, B. Northern Sweden Monitoring of Trends and Determinants in Cardiovascular Disease. Impaired incretin response after a mixed meal is associated with insulin resistance in nondiabetic men. Diabetes Care 2001, 24, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Fu-Cheng, X.; Anini, Y.; Chariot, J.; Castex, N.; Galmiche, J.P.; Roze, C. Mechanisms of peptide YY release induced by an intraduodenal meal in rats: Neural regulation by proximal gut. Pflug. Arch. 1997, 433, 571–579. [Google Scholar] [CrossRef]
- Rocca, A.S.; Brubaker, P.L. Role of the vagus nerve in mediating proximal nutrient-induced glucagon-like peptide-1 secretion. Endocrinology 1999, 140, 1687–1694. [Google Scholar] [CrossRef]
- Schwartz, G.J.; Moran, T.H. Duodenal nutrient exposure elicits nutrient-specific gut motility and vagal afferent signals in rat. Am. J. Physiol. 1998, 274, R1236–R1242. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Anini, Y.; Hansotia, T.; Brubaker, P.L. Muscarinic receptors control postprandial release of glucagon-like peptide-1: In vivo and in vitro studies in rats. Endocrinology 2002, 143, 2420–2426. [Google Scholar] [CrossRef] [PubMed]
- Anini, Y.; Brubaker, P.L. Muscarinic receptors control glucagon-like peptide 1 secretion by human endocrine L cells. Endocrinology 2003, 144, 3244–3250. [Google Scholar] [PubMed] [Green Version]
- Brubaker, P.L.; Schloos, J.; Drucker, D.J. Regulation of glucagon-like peptide-1 synthesis and secretion in the GLUTag enteroendocrine cell line. Endocrinology 1998, 139, 4108–4114. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef] [Green Version]
- Lindqvist, A.; Shcherbina, L.; Fischer, A.T.; Wierup, N. Ghrelin is a Regulator of Glucagon-Like Peptide 1 Secretion and Transcription in Mice. Front. Endocrinol. (Lausanne) 2017, 8, 135. [Google Scholar]
- Gagnon, J.; Baggio, L.L.; Drucker, D.J.; Brubaker, P.L. Ghrelin Is a Novel Regulator of GLP-1 Secretion. Diabetes 2015, 64, 1513–1521. [Google Scholar]
- Hagemann, D.; Holst, J.J.; Gethmann, A.; Banasch, M.; Schmidt, W.E.; Meier, J.J. Glucagon-like peptide 1 (GLP-1) suppresses ghrelin levels in humans via increased insulin secretion. Regul. Pept. 2007, 143, 64–68. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein–coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, R.E.; Gerspach, A.C.; Gutmann, H.; Asarian, L.; Drewe, J.; Beglinger, C. The functional involvement of gut-expressed sweet taste receptors in glucose-stimulated secretion of glucagon-like peptide-1 (GLP-1) and peptide YY (PYY). Clin. Nutr. 2011, 30, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Rozengurt, E. Taste receptors in the gastrointestinal tract. I. Bitter taste receptors and alpha-gustducin in the mammalian gut. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G171-7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-Expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerspach, A.C.; Steinert, R.E.; Schonenberger, L.; Graber-Maier, A.; Beglinger, C. The role of the gut sweet taste receptor in regulating GLP-1, PYY, and CCK release in humans. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E317–E325. [Google Scholar] [CrossRef] [PubMed]
- Reimann, F.; Gribble, F.M. Glucose-Sensing in glucagon-like peptide-1-secreting cells. Diabetes 2002, 51, 2757–2763. [Google Scholar] [CrossRef] [Green Version]
- Tolhurst, G.; Zheng, Y.; Parker, H.E.; Habib, A.M.; Reimann, F.; Gribble, F.M. Glutamine triggers and potentiates glucagon-like peptide-1 secretion by raising cytosolic Ca2+ and cAMP. Endocrinology 2011, 152, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Sun, E.W.; de Fontgalland, D.; Rabbitt, P.; Hollington, P.; Sposato, L.; Due, S.L.; Wattchow, D.A.; Rayner, C.K.; Deane, A.M.; Young, R.L.; et al. Mechanisms Controlling Glucose-Induced GLP-1 Secretion in Human Small Intestine. Diabetes 2017, 66, 2144–2149. [Google Scholar] [CrossRef] [Green Version]
- Edfalk, S.; Steneberg, P.; Edlund, H. Gpr40 is expressed in enteroendocrine cells and mediates free fatty acid stimulation of incretin secretion. Diabetes 2008, 57, 2280–2287. [Google Scholar] [CrossRef] [Green Version]
- Christensen, L.W.; Kuhre, R.E.; Janus, C.; Svendsen, B.; Holst, J.J. Vascular, but not luminal, activation of FFAR1 (GPR40) stimulates GLP-1 secretion from isolated perfused rat small intestine. Physiol. Rep. 2015, 3, e12551. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauge, M.; Vestmar, M.A.; Husted, A.S.; Ekberg, J.P.; Wright, M.J.; Di Salvo, J.; Weinglass, A.B.; Engelstoft, M.S.; Madsen, A.N.; Lückmann, M. GPR40 (FFAR1)–combined Gs and Gq signaling in vitro is associated with robust incretin secretagogue action ex vivo and in vivo. Mol. Metab. 2015, 4, 3–14. [Google Scholar] [CrossRef]
- Rozengurt, E.; Sternini, C. Taste receptor signaling in the mammalian gut. Curr. Opin. Pharmacol. 2007, 7, 557–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nøhr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; Sichlau, R.M.; Grunddal, K.V.; Seier Poulsen, S.; Han, S. GPR41/FFAR3 and GPR43/FFAR2 as cosensors for short-chain fatty acids in enteroendocrine cells vs FFAR3 in enteric neurons and FFAR2 in enteric leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.C.; Kien, C.L.; Bouthillier, L.; Levy, E. Short-Chain fatty acids: Ready for prime time? Nutr. Clin. Pract. 2006, 21, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.-Y.; Lannoy, V.; Decobecq, M.-E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lejeune, M.P.; Westerterp, K.R.; Adam, T.C.; Luscombe-Marsh, N.D.; Westerterp-Plantenga, M.S. Ghrelin and glucagon-like peptide 1 concentrations, 24-h satiety, and energy and substrate metabolism during a high-protein diet and measured in a respiration chamber. Am. J. Clin. Nutr. 2006, 83, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Reimer, R.A.; Russell, J.C. Glucose tolerance, lipids, and GLP-1 secretion in JCR:LA-cp rats fed a high protein fiber diet. Obesity (Silver Spring) 2008, 16, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Diakogiannaki, E.; Pais, R.; Tolhurst, G.; Parker, H.E.; Horscroft, J.; Rauscher, B.; Zietek, T.; Daniel, H.; Gribble, F.M.; Reimann, F. Oligopeptides stimulate glucagon-like peptide-1 secretion in mice through proton-coupled uptake and the calcium-sensing receptor. Diabetologia 2013, 56, 2688–2696. [Google Scholar] [CrossRef] [Green Version]
- Vig, B.S.; Stouch, T.R.; Timoszyk, J.K.; Quan, Y.; Wall, D.A.; Smith, R.L.; Faria, T.N. Human PEPT1 pharmacophore distinguishes between dipeptide transport and binding. J. Med. Chem. 2006, 49, 3636–3644. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Aliaga, I.; Daniel, H. Peptide transporters and their roles in physiological processes and drug disposition. Xenobiotica 2008, 38, 1022–1042. [Google Scholar] [CrossRef] [PubMed]
- Hebert, S.C.; Cheng, S.; Geibel, J. Functions and roles of the extracellular Ca2+-sensing receptor in the gastrointestinal tract. Cell Calcium 2004, 35, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Conigrave, A.D.; Mun, H.C.; Lok, H.C. Aromatic L-amino acids activate the calcium-sensing receptor. J. Nutr. 2007, 137 (Suppl. 1), 1524S–1527S, discussion 1548S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamshah, A.; Spreckley, E.; Norton, M.; Kinsey-Jones, J.S.; Amin, A.; Ramgulam, A.; Cao, Y.; Johnson, R.; Saleh, K.; Akalestou, E.; et al. l-phenylalanine modulates gut hormone release and glucose tolerance, and suppresses food intake through the calcium-sensing receptor in rodents. Int. J. Obes. 2017, 41, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Acar, I.; Cetinkaya, A.; Lay, I.; Ileri-Gurel, E. The role of calcium sensing receptors in GLP-1 and PYY secretion after acute intraduodenal administration of L-Tryptophan in rats. Nutr. Neurosci. 2018, 1–9. [Google Scholar] [CrossRef]
- Mace, O.J.; Schindler, M.; Patel, S. The regulation of K- and L-cell activity by GLUT2 and the calcium-sensing receptor CasR in rat small intestine. J. Physiol. 2012, 590, 2917–2936. [Google Scholar] [CrossRef]
- Oya, M.; Kitaguchi, T.; Pais, R.; Reimann, F.; Gribble, F.; Tsuboi, T. The G protein-coupled receptor family C group 6 subtype A (GPRC6A) receptor is involved in amino acid-induced glucagon-like peptide-1 secretion from GLUTag cells. J. Biol. Chem. 2013, 288, 4513–4521. [Google Scholar] [CrossRef] [Green Version]
- Reimann, F.; Williams, L.; da Silva Xavier, G.; Rutter, G.A.; Gribble, F.M. Glutamine potently stimulates glucagon-like peptide-1 secretion from GLUTag cells. Diabetologia 2004, 47, 1592–1601. [Google Scholar] [CrossRef] [Green Version]
- Andersson, L.E.; Shcherbina, L.; Al-Majdoub, M.; Vishnu, N.; Arroyo, C.B.; Aste Carrara, J.; Wollheim, C.B.; Fex, M.; Mulder, H.; Wierup, N.; et al. Glutamine-Elicited Secretion of Glucagon-Like Peptide 1 Is Governed by an Activated Glutamate Dehydrogenase. Diabetes 2018, 67, 372–384. [Google Scholar]
- Dinda, B.; Debnath, S.; Banik, R. Naturally occurring iridoids and secoiridoids. An updated review, part 4. Chem. Pharm. Bull. 2011, 59, 803–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, K. Occurrence and content of hydroxycinnamic and hydroxybenzoic acid compounds in foods. Crit. Rev. Food Sci. Nutr. 1989, 28, 315–347. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.; Samman, S. Flavonoids—Chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Namiki, M. The chemistry and physiological functions of sesame. Food Rev. Int. 2009, 11, 281–329. [Google Scholar] [CrossRef]
- Mazur, W.; Fotsis, T.; Wähälä, K.; Ojala, S.; Salakka, A.; Adlercreutz, H. Isotope dilution gas chromatographic–mass spectrometric method for the determination of isoflavonoids, coumestrol, and lignans in food samples. Anal. Biochem. 1996, 233, 169–180. [Google Scholar] [CrossRef]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J. Nutr. 2005, 135, 48–52. [Google Scholar]
- Piskula, M.K. Factors affecting flavonoids absorption. Biofactors 2000, 12, 175–180. [Google Scholar] [CrossRef]
- Walle, T.; Otake, Y.; Walle, U.K.; Wilson, F.A. Quercetin glucosides are completely hydrolyzed in ileostomy patients before absorption. J. Nutr. 2000, 130, 2658–2661. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.; Morgan, M.R.; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver beta-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, K.; Plumb, G.W.; Berrin, J.G.; Juge, N.; Jacob, R.; Naim, H.Y.; Williamson, G.; Swallow, D.M.; Kroon, P.A. Deglycosylation by small intestinal epithelial cell beta-glucosidases is a critical step in the absorption and metabolism of dietary flavonoid glycosides in humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Vari, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P.E.; Chowrimootoo, G.; Choudhury, R.; Debnam, E.S.; Srai, S.K.; Rice-Evans, C. The small intestine can both absorb and glucuronidate luminal flavonoids. FEBS Lett. 1999, 458, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Piskula, M.K.; Terao, J. Accumulation of (-)-epicatechin metabolites in rat plasma after oral administration and distribution of conjugation enzymes in rat tissues. J. Nutr. 1998, 128, 1172–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Morand, C.; Texier, O.; Favier, M.L.; Agullo, G.; Demigne, C.; Regerat, F.; Remesy, C. Quercetin metabolites in plasma of rats fed diets containing rutin or quercetin. J. Nutr. 1995, 125, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Updated knowledge about polyphenols: Functions, bioavailability, metabolism, and health. Crit. Rev. Food Sci. Nutr. 2012, 52, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Crespy, V.; Morand, C.; Manach, C.; Besson, C.; Demigne, C.; Remesy, C. Part of quercetin absorbed in the small intestine is conjugated and further secreted in the intestinal lumen. Am. J. Physiol. Gastrointest. Liver Physiol. 1999, 277, G120–G126. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Jung, E.A.; Sohng, I.S.; Han, J.A.; Kim, T.H.; Han, M.J. Intestinal bacterial metabolism of flavonoids and its relation to some biological activities. Arch. Pharm. Res. 1998, 21, 17–23. [Google Scholar] [CrossRef]
- Meselhy, M.R.; Nakamura, N.; Hattori, M. Biotransformation of (−)-epicatechin 3-O-gallate by human intestinal bacteria. Chem. Pharm. Bull. 1997, 45, 888–893. [Google Scholar] [CrossRef] [Green Version]
- Takikawa, M.; Kurimoto, Y.; Tsuda, T. Curcumin stimulates glucagon-like peptide-1 secretion in GLUTag cells via Ca2+/calmodulin-dependent kinase II activation. Biochem. Biophys. Res. Commun. 2013, 435, 165–170. [Google Scholar] [CrossRef]
- Alli-Oluwafuyi, A.M.; Luis, P.B.; Nakashima, F.; Gimenez-Bastida, J.A.; Presley, S.H.; Duvernay, M.T.; Iwalewa, E.O.; Schneider, C. Curcumin induces secretion of glucagon-like peptide-1 through an oxidation-dependent mechanism. Biochimie 2019, 165, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Nishikawa, S.; Ikehata, A.; Dochi, K.; Tani, T.; Takahashi, T.; Imaizumi, A.; Tsuda, T. Curcumin improves glucose tolerance via stimulation of glucagon-like peptide-1 secretion. Mol. Nutr. Food Res. 2017, 61, 1600471. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Tani, T.; Terahara, N.; Tsuda, T. The Anthocyanin Delphinidin 3-Rutinoside Stimulates Glucagon-Like Peptide-1 Secretion in Murine GLUTag Cell Line via the Ca2+/Calmodulin-Dependent Kinase II Pathway. PLoS ONE 2015, 10, e0126157. [Google Scholar] [CrossRef] [PubMed]
- Tani, T.; Nishikawa, S.; Kato, M.; Tsuda, T. Delphinidin 3-rutinoside-rich blackcurrant extract ameliorates glucose tolerance by increasing the release of glucagon-like peptide-1 secretion. Food Sci. Nutr. 2017, 5, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, Y.; Ozeki, A.; Tani, T.; Tsuda, T. Blackcurrant extract ameliorates hyperglycemia in type 2 diabetic mice in association with increased basal secretion of glucagon-like peptide-1 and activation of AMP-activated protein kinase. J. Nutr. Sci. Vitaminol. 2018, 64, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Castro-Acosta, M.L.; Smith, L.; Miller, R.J.; McCarthy, D.I.; Farrimond, J.A.; Hall, W.L. Drinks containing anthocyanin-rich blackcurrant extract decrease postprandial blood glucose, insulin and incretin concentrations. J. Nutr. Biochem. 2016, 38, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Hirasawa, A.; Sun, Q.; Sadakane, K.; Itsubo, C.; Iga, T.; Adachi, T.; Koshimizu, T.-A.; Hashimoto, T.; Asakawa, Y. Novel selective ligands for free fatty acid receptors GPR120 and GPR40. Naunyn Schmiedeberg’s Arch. Pharmacol. 2009, 380, 247–255. [Google Scholar] [CrossRef]
- Cho, H.; Kim, K.; Kim, N.; Woo, M.; Kim, H.Y. Effect of propolis phenolic compounds on free fatty acid receptor 4 activation. Food Sci. Biotechnol. 2019, 1–6. [Google Scholar] [CrossRef]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.; Satta, Y.; Takenaka, O.; Takahata, N. Lineage-Specific loss of function of bitter taste receptor genes in humans and nonhuman primates. Genetics 2005, 170, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Rozengurt, N.; Wu, S.V.; Chen, M.C.; Huang, C.; Sternini, C.; Rozengurt, E. Colocalization of the α-subunit of gustducin with PYY and GLP-1 in L cells of human colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291. [Google Scholar] [CrossRef]
- Wu, S.V.; Chen, M.C.; Rozengurt, E. Genomic organization, expression, and function of bitter taste receptors (T2R) in mouse and rat. Physiol. Genom. 2005, 22, 139–149. [Google Scholar] [CrossRef]
- Wu, S.V.; Rozengurt, N.; Yang, M.; Young, S.H.; Sinnett-Smith, J.; Rozengurt, E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2392–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolskee, R.F. Molecular mechanisms of bitter and sweet taste transduction. J. Biol. Chem. 2002, 277, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canivenc-Lavier, M.-C.; Neiers, F.; Briand, L. Plant polyphenols, chemoreception, taste receptors and taste management. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 472–478. [Google Scholar] [PubMed]
- Zhang, Y.; Wang, X.; Li, X.; Peng, S.; Wang, S.; Huang, C.Z.; Huang, C.Z.; Zhang, Q.; Li, D.; Jiang, J.; et al. Identification of a specific agonist of human TAS2R14 from Radix Bupleuri through virtual screening, functional evaluation and binding studies. Sci. Rep. 2017, 7, 12174. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Silva, M.S.; Garcia-Estevez, I.; Groβmann, P.; Brás, N.R.; Brandão, E.; Mateus, N.; de Freitas, V.; Behrens, M.; Meyerhof, W. Human bitter taste receptors are activated by different classes of polyphenols. J. Agric. Food Chem. 2018, 66, 8814–8823. [Google Scholar] [CrossRef]
- Soares, S.; Kohl, S.; Thalmann, S.; Mateus, N.; Meyerhof, W.; De Freitas, V. Different phenolic compounds activate distinct human bitter taste receptors. J. Agric. Food Chem. 2013, 61, 1525–1533. [Google Scholar] [CrossRef]
- Kurogi, M.; Miyashita, M.; Emoto, Y.; Kubo, Y.; Saitoh, O. Green tea polyphenol epigallocatechin gallate activates TRPA1 in an intestinal enteroendocrine cell line, STC-1. Chem. Senses 2012, 37, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.-Y.; Aihara, Y.; Hashimoto, T.; Kanazawa, K.; Mizuno, M. (−)-Epigallocatechin-3-gallate induces secretion of anorexigenic gut hormones. J. Clin. Biochem. Nutr. 2015, 57, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.-Y.; Huang, C.-J.; Huang, L.-H.; Chen, I.-J.; Chiu, J.-P.; Hsu, C.-H. Effects of green tea extract on insulin resistance and glucagon-like peptide 1 in patients with type 2 diabetes and lipid abnormalities: A randomized, double-blinded, and placebo-controlled trial. PLoS ONE 2014, 9, e91163. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sayama, K. The combined administration of EGCG and caffeine induces not only suppression of fat accumulation but also anorexigenic action in mice. J. Funct. Foods 2018, 47, 156–162. [Google Scholar] [CrossRef]
- Kim, K.S.; Egan, J.M.; Jang, H.J. Denatonium induces secretion of glucagon-like peptide-1 through activation of bitter taste receptor pathways. Diabetologia 2014, 57, 2117–2125. [Google Scholar]
- Casanova-Martí, À.; Serrano, J.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. Acute selective bioactivity of grape seed proanthocyanidins on enteroendocrine secretions in the gastrointestinal tract. Food Nutr. Res. 2017, 61, 1321347. [Google Scholar] [CrossRef] [Green Version]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Green, B.D.; Pinent, M.; Ardévol, A. Grape-Seed procyanidins modulate cellular membrane potential and nutrient-induced GLP-1 secretion in STC-1 cells. Am. J. Physiol. Cell Physiol. 2013, 306, C485–C492. [Google Scholar] [CrossRef] [Green Version]
- Reimann, F.; Tolhurst, G.; Gribble, F.M. G-protein-coupled receptors in intestinal chemosensation. Cell Metab. 2012, 15, 421–431. [Google Scholar]
- Lacroix, S.; Badoux, J.K.; Scott-Boyer, M.-P.; Parolo, S.; Matone, A.; Priami, C.; Morine, M.J.; Kaput, J.; Moco, S. A computationally driven analysis of the polyphenol-protein interactome. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mace, O.; Tehan, B.; Marshall, F. Pharmacology and physiology of gastrointestinal enteroendocrine cells. Pharmacol. Res. Perspect. 2015, 3, e00155. [Google Scholar] [CrossRef]
- Yu, Z.; Jin, T. New insights into the role of cAMP in the production and function of the incretin hormone glucagon-like peptide-1 (GLP-1). Cell. Signal. 2010, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Glass, L.L.; Sharp, S.J.; Reimann, F.; Gribble, F.M. Galanin inhibits GLP-1 and GIP secretion via the GAL1 receptor in enteroendocrine L and K cells. Br. J. Pharmacol. 2016, 173, 888–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.A.; Velasquez, M.T.; Hansen, C.T.; Mohamed, A.I.; Bhathena, S.J. Modulation of carbohydrate metabolism and peptide hormones by soybean isoflavones and probiotics in obesity and diabetes. J. Nutr. Biochem. 2005, 16, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Mezei, O.; Banz, W.J.; Steger, R.W.; Peluso, M.R.; Winters, T.A.; Shay, N. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [CrossRef] [Green Version]
- Atteritano, M.; Marini, H.; Minutoli, L.; Polito, F.; Bitto, A.; Altavilla, D.; Mazzaferro, S.; D’Anna, R.; Cannata, M.L.; Gaudio, A.; et al. Effects of the phytoestrogen genistein on some predictors of cardiovascular risk in osteopenic, postmenopausal women: A two-year randomized, double-blind, placebo-controlled study. J. Clin. Endocrinol. Metab. 2007, 92, 3068–3075. [Google Scholar] [CrossRef] [PubMed]
- Villa, P.; Costantini, B.; Suriano, R.; Perri, C.; Macri, F.; Ricciardi, L.; Panunzi, S.; Lanzone, A. The differential effect of the phytoestrogen genistein on cardiovascular risk factors in postmenopausal women: Relationship with the metabolic status. J. Clin. Endocrinol. Metab. 2009, 94, 552–558. [Google Scholar] [PubMed] [Green Version]
- Liu, D.; Zhen, W.; Yang, Z.; Carter, J.D.; Si, H.; Reynolds, K.A. Genistein acutely stimulates insulin secretion in pancreatic beta-cells through a cAMP-dependent protein kinase pathway. Diabetes 2006, 55, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.S.; Jung, U.J.; Yeo, J.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res. Rev. 2008, 24, 74–81. [Google Scholar] [CrossRef]
- Guo, T.L.; Germolec, D.R.; Zheng, J.F.; Kooistra, L.; Auttachoat, W.; Smith, M.J.; White, K.L.; Elmore, S.A. Genistein protects female nonobese diabetic mice from developing type 1 diabetes when fed a soy- and alfalfa-free diet. Toxicol. Pathol. 2015, 43, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Wang, A.; Zhen, W.; Wang, Y.; Si, H.; Jia, Z.; Alkhalidy, H.; Cheng, Z.; Gilbert, E.; Xu, B.; et al. Phytonutrient genistein is a survival factor for pancreatic beta-cells via GPR30-mediated mechanism. J. Nutr. Biochem. 2018, 58, 59–70. [Google Scholar] [CrossRef]
- Fu, Z.; Zhang, W.; Zhen, W.; Lum, H.; Nadler, J.; Bassaganya-Riera, J.; Jia, Z.; Wang, Y.; Misra, H.; Liu, D. Genistein induces pancreatic beta-cell proliferation through activation of multiple signaling pathways and prevents insulin-deficient diabetes in mice. Endocrinology 2010, 151, 3026–3037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, K.; Ali, M.B.; Akash, M.S.H. Genistein enhances the secretion of glucagon-like peptide-1 (GLP-1) via downregulation of inflammatory responses. Biomed. Pharmacother. 2019, 112, 108670. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, A.; Alkhalidy, H.; Luo, J.; Moomaw, E.; Neilson, A.P.; Liu, D. Flavone Hispidulin Stimulates Glucagon-Like Peptide-1 Secretion and Ameliorates Hyperglycemia in Streptozotocin-Induced Diabetic Mice. Mol. Nutr. Food Res. 2020, 64, e1900978. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Qin, W.; Wu, F.; Wang, S.; Pan, C.; Wang, L.; Zeng, B.; Ma, S.; Liang, J. Apigenin and naringenin regulate glucose and lipid metabolism, and ameliorate vascular dysfunction in type 2 diabetic rats. Eur. J. Pharmacol. 2016, 773, 13–23. [Google Scholar] [CrossRef]
- Xu, N.; Zhang, L.; Dong, J.; Zhang, X.; Chen, Y.G.; Bao, B.; Liu, J. Low-Dose diet supplement of a natural flavonoid, luteolin, ameliorates diet-induced obesity and insulin resistance in mice. Mol. Nutr. Food Res. 2014, 58, 1258–1268. [Google Scholar]
- Chang, K.-J.; Lin, J.-A.; Chen, S.-Y.; Weng, M.-H.; Yen, G.-C. Silymarin protects against high fat diet-evoked metabolic injury by induction of glucagon-like peptide 1 and sirtuin 1. J. Funct. Foods 2019, 56, 136–144. [Google Scholar] [CrossRef]
- Meng, R.; Mahadevan, J.; Oseid, E.; Vallerie, S.; Robertson, R.P. Silymarin activates c-AMP phosphodiesterase and stimulates insulin secretion in a glucose-dependent manner in HIT-T15 cells. Antioxidants 2016, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Jokura, H.; Watanabe, I.; Umeda, M.; Hase, T.; Shimotoyodome, A. Coffee polyphenol consumption improves postprandial hyperglycemia associated with impaired vascular endothelial function in healthy male adults. Nutr. Res. 2015, 35, 873–881. [Google Scholar] [CrossRef]
- Nordestgaard, A.T.; Thomsen, M.; Nordestgaard, B.G. Coffee intake and risk of obesity, metabolic syndrome and type 2 diabetes: A Mendelian randomization study. Int. J. Epidemiol. 2015, 44, 551–565. [Google Scholar] [CrossRef]
- Montoya, G.A.; Bakuradze, T.; Eirich, M.; Erk, T.; Baum, M.; Habermeyer, M.; Eisenbrand, G.; Richling, E. Modulation of 3′, 5′-cyclic AMP homeostasis in human platelets by coffee and individual coffee constituents. Br. J. Nutr. 2014, 112, 1427–1437. [Google Scholar]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial community variation in human body habitats across space and time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Costello, E.K.; Berg-Lyons, D.; Gonzalez, A.; Stombaugh, J.; Knights, D.; Gajer, P.; Ravel, J.; Fierer, N.; et al. Moving pictures of the human microbiome. Genome Biol. 2011, 12, R50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- McOrist, A.L.; Miller, R.B.; Bird, A.R.; Keogh, J.B.; Noakes, M.; Topping, D.L.; Conlon, M.A. Fecal Butyrate Levels Vary Widely among Individuals but Are Usually Increased by a Diet High in Resistant Starch. J. Nutr. 2011, 141, 883–889. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.F.; Maier, C.S. The Chemistry of Gut Microbial Metabolism of Polyphenols. Phytochem. Rev. 2016, 15, 425–444. [Google Scholar] [CrossRef] [Green Version]
- Schneider, H.; Simmering, R.; Hartmann, L.; Pforte, H.; Blaut, M. Degradation of quercetin-3-glucoside in gnotobiotic rats associated with human intestinal bacteria. J. Appl. Microbiol. 2000, 89, 1027–1037. [Google Scholar] [CrossRef]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuno, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Roopchand, D.E.; Carmody, R.N.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.J.; Raskin, I. Dietary polyphenols promote growth of the gut bacterium Akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [PubMed] [Green Version]
- Seo, D.B.; Jeong, H.W.; Cho, D.; Lee, B.J.; Lee, J.H.; Choi, J.Y.; Bae, I.H.; Lee, S.J. Fermented green tea extract alleviates obesity and related complications and alters gut microbiota composition in diet-induced obese mice. J. Med. Food 2015, 18, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Belzer, C.; Goossens, M.; Kleerebezem, M.; De Vos, W.M.; Thas, O.; De Weirdt, R.; Kerckhof, F.M.; Van de Wiele, T. Butyrate-Producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 2013, 7, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar]
- Duncan, S.H.; Barcenilla, A.; Stewart, C.S.; Pryde, S.E.; Flint, H.J. Acetate utilization and butyryl coenzyme A (CoA):acetate-CoA transferase in butyrate-producing bacteria from the human large intestine. Appl. Environ. Microbiol. 2002, 68, 5186–5190. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-Utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.L.; Wolin, M.J. Pathways of acetate, propionate, and butyrate formation by the human fecal microbial flora. Appl. Environ. Microbiol. 1996, 62, 1589–1592. [Google Scholar] [CrossRef] [Green Version]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar]
- Henning, S.M.; Summanen, P.H.; Lee, R.P.; Yang, J.; Finegold, S.M.; Heber, D.; Li, Z. Pomegranate ellagitannins stimulate the growth of Akkermansia muciniphila in vivo. Anaerobe 2017, 43, 56–60. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sanchez-Alcoholado, L.; Perez-Martinez, P.; Andres-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuno, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Cuervo, A.; Reyes-Gavilán, C.G.D.L.; Ruas-Madiedo, P.; Lopez, P.; Suarez, A.; Gueimonde, M.; González, S. Red wine consumption is associated with fecal microbiota and malondialdehyde in a human population. J. Am. Coll. Nutr. 2015, 34, 135–141. [Google Scholar]
- Cheng, M.; Zhang, X.; Zhu, J.; Cheng, L.; Cao, J.; Wu, Z.; Weng, P.; Zheng, X. A metagenomics approach to the intestinal microbiome structure and function in high fat diet-induced obesity mice fed with oolong tea polyphenols. Food Funct. 2018, 9, 1079–1087. [Google Scholar] [CrossRef]
- Kim, H.; Krenek, K.A.; Fang, C.; Minamoto, Y.; Markel, M.E.; Suchodolski, J.S.; Talcott, S.T.; Mertens-Talcott, S.U. Polyphenolic derivatives from mango (Mangifera indica L.) modulate fecal microbiome, short-chain fatty acids production and the HDAC1/AMPK/LC3 axis in rats with DSS-induced colitis. J. Funct. Foods 2018, 48, 243–251. [Google Scholar]
- Unno, T.; Osakabe, N. Green tea extract and black tea extract differentially influence cecal levels of short-chain fatty acids in rats. Food Sci. Nutr. 2018, 6, 728–735. [Google Scholar] [CrossRef]
- Unno, T.; Sakuma, M.; Mitsuhashi, S. Effect of dietary supplementation of (−)-epigallocatechin gallate on gut microbiota and biomarkers of colonic fermentation in rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2014, 60, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marteau, P.; Pochart, P.; Doré, J.; Béra-Maillet, C.; Bernalier, A.; Corthier, G. Comparative study of bacterial groups within the human cecal and fecal microbiota. Appl. Environ. Microbiol. 2001, 67, 4939–4942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; de Mejia, E.G. Berry and Citrus Phenolic Compounds Inhibit Dipeptidyl Peptidase IV: Implications in Diabetes Management. Evid. Based Complement. Altern. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef] [Green Version]
- Altenhofen, D.; da Luz, G.; Frederico, M.J.; Venzke, D.; Brich, M.; Vigil, S.; Frode, T.S.; Linares, C.E.; Pizzolatti, M.G.; Silva, F.R. Bis-Pyrano Prenyl Isoflavone Improves Glucose Homeostasis by Inhibiting Dipeptidyl Peptidase-4 in Hyperglycemic Rats. J. Cell. Biochem. 2017, 118, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Jacobson, G.A.; Burgess, J.R.; Jelinek, H.F.; Nichols, D.S.; Narkowicz, C.K.; Al-Aubaidy, H.A. Citrus bioflavonoids dipeptidyl peptidase-4 inhibition compared with gliptin antidiabetic medications. Biochem. Biophys. Res. Commun. 2018, 503, 21–25. [Google Scholar] [CrossRef]
- Proenca, C.; Freitas, M.; Ribeiro, D.; Tome, S.M.; Araujo, A.N.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. The dipeptidyl peptidase-4 inhibitory effect of flavonoids is hindered in protein rich environments. Food Funct. 2019, 10, 5718–5731. [Google Scholar]
- Bar, J.; Weber, A.; Hoffmann, T.; Stork, J.; Wermann, M.; Wagner, L.; Aust, S.; Gerhartz, B.; Demuth, H.U. Characterisation of human dipeptidyl peptidase IV expressed in Pichia pastoris. A structural and mechanistic comparison between the recombinant human and the purified porcine enzyme. Biol. Chem. 2003, 384, 1553–1563. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Pujadas, G.; Garcia-Vallvé, S.; Pinent, M.; Ardévol, A. Grape seed-derived procyanidins decrease dipeptidyl-peptidase 4 activity and expression. J. Agric. Food Chem. 2012, 60, 9055–9061. [Google Scholar] [CrossRef]
- Matheeussen, V.; Lambeir, A.M.; Jungraithmayr, W.; Gomez, N.; Mc Entee, K.; Van der Veken, P.; Scharpe, S.; De Meester, I. Method comparison of dipeptidyl peptidase IV activity assays and their application in biological samples containing reversible inhibitors. Clin. Chim. Acta 2012, 413, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Ansari, J.A.; Naz, S.; Tarar, O.M.; Siddiqi, R.; Haider, M.S.; Jamil, K. Binding effect of proline-rich-proteins (PRPs) on in vitro antimicrobial activity of the flavonoids. Braz. J. Microbiol. 2015, 46, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Patel, P.K.; Choudhary, K.; Joshi, J.; Yadav, D.; Jin, J.O. Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo. Biomolecules 2020, 10, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, M.; Yu, D.M.; O’Connor, S.; Gorrell, M.D. Inhibitor selectivity in the clinical application of dipeptidyl peptidase-4 inhibition. Clin. Sci. (Lond.) 2009, 118, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.K.; Lin, S.R.; Chang, C.H.; Tsai, M.J.; Lee, D.N.; Weng, C.F. Natural phenolic compounds potentiate hypoglycemia via inhibition of Dipeptidyl peptidase IV. Sci. Rep. 2019, 9, 15585. [Google Scholar]
- Kalhotra, P.; Chittepu, V.; Osorio-Revilla, G.; Gallardo-Velázquez, T. Structure-Activity Relationship and Molecular Docking of Natural Product Library Reveal Chrysin as a Novel Dipeptidyl Peptidase-4 (DPP-4) Inhibitor: An Integrated In Silico and In Vitro Study. Molecules 2018, 23, 1368. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Shree, P.; Tripathi, Y.B. Active phytochemicals of Pueraria tuberosa for DPP-IV inhibition: In silico and experimental approach. J. Diabetes Metab. Disord. 2017, 16, 46. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic overview of nutrient-induced GLP-1 secretion from L-cells. AC: adenylyl cyclase; DAG: diacylglycerol; GLP-1: glucagon-like peptide-1; GPCRs: G-protein coupled receptors; IP3: inositol trisphosphate; PLC-β: phospholipase C-β; PKA: protein kinase A; PKC: protein kinase C; PIP2: phosphatidylinositol 4,5-biphosphate; SGLT1: sodium/glucose co-transporter 1; TRPM5: transient receptor potential channel M5.
Figure 1.
Schematic overview of nutrient-induced GLP-1 secretion from L-cells. AC: adenylyl cyclase; DAG: diacylglycerol; GLP-1: glucagon-like peptide-1; GPCRs: G-protein coupled receptors; IP3: inositol trisphosphate; PLC-β: phospholipase C-β; PKA: protein kinase A; PKC: protein kinase C; PIP2: phosphatidylinositol 4,5-biphosphate; SGLT1: sodium/glucose co-transporter 1; TRPM5: transient receptor potential channel M5.

Figure 2.
Chemical structures of polyphenols from four categories.


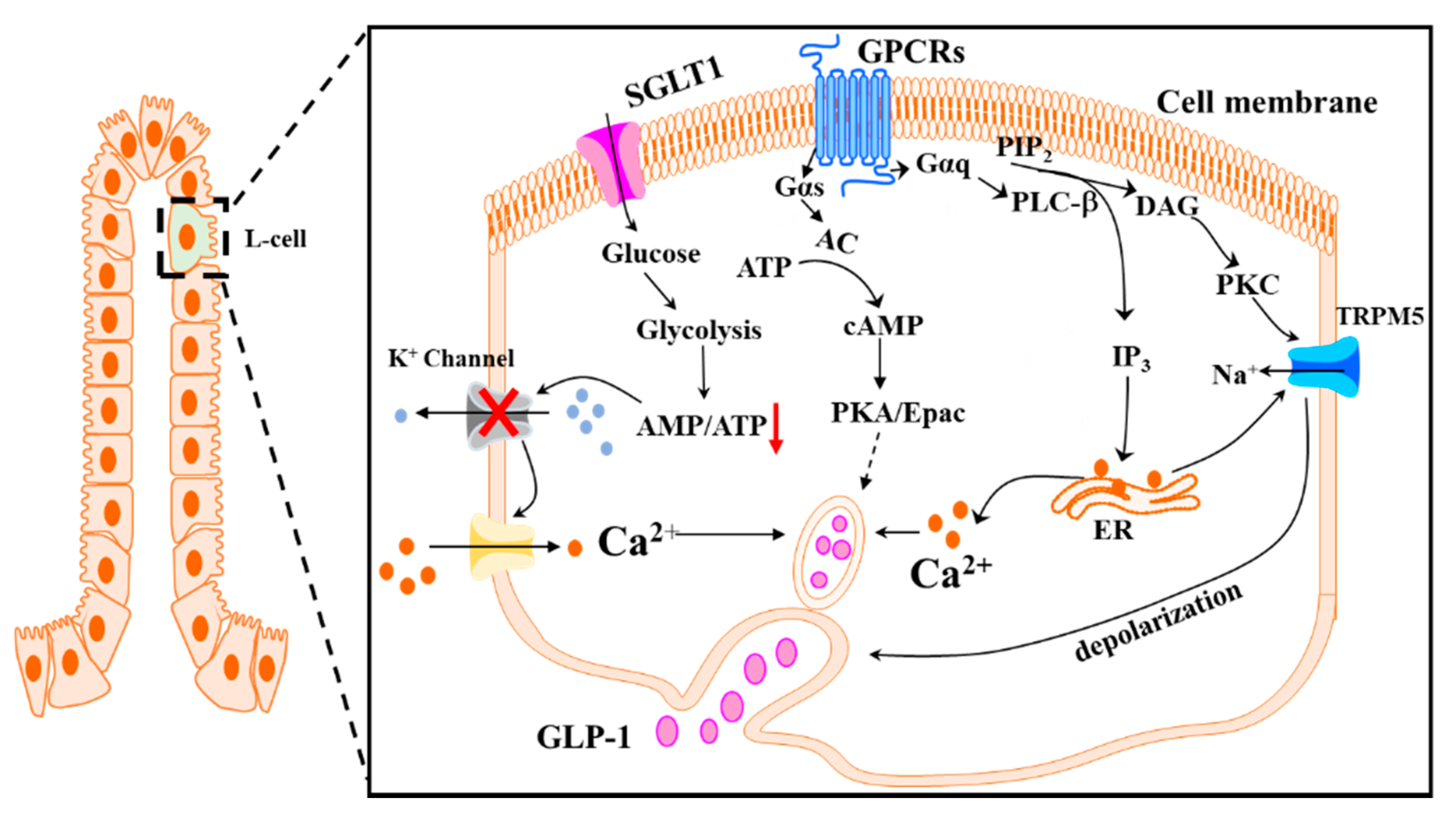{kind=link}
{kind=link}
Table 1.
Polyphenolic compounds that regulate GLP-1 secretion via G protein coupled receptor (GPCR)- or cAMP- mediated mechanism.
Table 1.
Polyphenolic compounds that regulate GLP-1 secretion via G protein coupled receptor (GPCR)- or cAMP- mediated mechanism.
| Polyphenolic Compounds | Chemical Structure | Dose | Model | Effect on GLP1 | Possible Mechanisms |
|---|---|---|---|---|---|
| Curcumin [125,127] |  | 10 μM 1.5 mg/kg b.w. | Glutag L-cells, Rat | Increase GLP-1 secretion; increase plasma GLP-1 level | GPR40/120-dependent pathway |
| Delphinidin 3-rutinoside [128] |  | 10 μM | Glutag L-cells | Increase GLP-1 secretion | GPR40/120-dependent pathway |
| EGCG [145,146,147] |  | 300 μM 500 mg | Mouse ileum segment, diabetic patients | Increase GLP-1 secretion; increase plasma GLP-1 level | Tas2Rs-dependent pathway |
| Genistein [165] |  | 20 mg/kg b.w. | Alloxan-induced insulin deficient diabetic rats [165] | Increase tissue content of GLP-1 | cAMP signaling by activating AC |
| Grifolic acid [132] |  | 30 μM | STC-1 cells | Increase GLP-1 secretion | GPR120/Ca2+ signaling |
| Grifolic acid methyl ether [132] |  | 30 μM | STC-1 cells | Increase GLP-1 secretion | GPR120/Ca2+ signaling |
| Hispidulin [166] |  | 1 μM 20 mg/kg b.w. | Glutag L-cell, STZ-induced diabetic mice | Increase GLP-1 secretion; increase plasma GLP-1 level | Inhibiting PDE activity |
| Silymarin [169] |  | 100 mg/kg b.w. | Obese rats | Increased serum GLP-1 level | Inhibiting PDE activity |
| Caffeoylquinic acid [173] |  | IC50: 0.49 mM | Human plasma platelets | Not applicable | Inhibiting PDE activity |
| Caffeic acid [173] |  | IC50: 0.48 mM | Human plasma platelets | Not applicable | Inhibiting PDE activity |
Table 2.
Polyphenols regulates short chain fatty acid (SCFA)-producing microbiota.
| Polyphenols | Polyphenolic Composition | Dose | Model | Effects on Microbiota |
|---|---|---|---|---|
| Catechins [196] | (+)-Catechin | 150 mg/mL | Ex vivo: human fecal microbiota | Increase the growth of E. rectale |
| Chlorogenic acid [197] | Chlorogenic acid | 10 μg/mL | Ex vivo: human fecal microbiota | Increase the population of bifidobacterium species |
| Pomegranate polyphenols [199] | Not available | 1000 mg/day | Healthy subjects | Increase the population of A. muciniphila |
| Red wine [200] | 14% flavan-3-ols, 3.6% anthocyanins, 2.9% hydroxybenzoic acids | 272 mL/day | Healthy and obese subjects | Increase the proportion of F. prausnitzii in obese subjects |
| Oolong tea polyphenols [202] | Catechins (88.79 μg/g) | 0.1% oolong tea polyphenols in the high-fat diet | Mice | Increase the proportion of F. prausnitzii and other SCFA-producing bacteria, as well as increase in the fecal contents of SCFAs |
| Mango polyphenol extracts [40] | Gallic acid (7.35 mg/L) | ad libitum | Rats | Increase Clostridium butyrium, and improve butyrate production |
| Black tea or green tea polyphenols [204] | Catechines (275 g/kg in green tea, and 35 g/kg in black tea) | 10 g/kg/day | Rats | Reduce the abundance of Clostridium XIVa |
| EGCG [205] | EGCG | 0.3% in the diet | Rats | Reduce the abundance of fecal Clostridial IV and XIVa bacteria |
Table 3.
Polyphenols regulates dipeptidyl peptidase IV (DPP-IV) activity.
| Polyphenolic Compounds | Effective Concentration | Detection Assay | Effect on DPP-IV Activity | Possible Mechanism |
|---|---|---|---|---|
| Bis-pyrano prenyl isoflavone [208] | 100 μM | DPP-4 fluorometric assay | 53% inhibition | Not available |
| Rutin [209] | IC50: 0.485 mM | DPP-4 fluorometric assay | 50% inhibition | Not available |
| Eriocitrin [207,209] | IC50: 5.44 mM; 10.36 ± 0.09 μM | DPP-4 fluorometric assay | 50% inhibition | H Bonds and π interactions |
| Eriodictyol [209] | IC50: 3.91 mM | DPP-4 fluorometric assay | 50% inhibition | Not available |
| Naringenin [207,209] | IC50: 5.5 mM; 0.24 ± 0.03 μM | DPP-4 fluorometric assay | 50% inhibition | H Bonds and π interactions |
| Naringin [209] | IC50: 3.82 mM | DPP-4 fluorometric assay | 50% inhibition | Not available |
| Hesperidin [209] | IC50: 5.7 mM | DPP-4 fluorometric assay | 50% inhibition | H Bonds and π interactions |
| Hesperetin [207,209] | IC50: 5.7 mM; 0.28 ± 0.07 μM | DPP-4 fluorometric assay | 50% inhibition | H Bonds and π interactions |
| Apigenin [207,210] | IC50: 0.14 ± 0.02 μM; 200 μM | DPP-IV GloTM Protease Assay | 50% inhibition | H Bonds and π interactions |
| Quercetin [207] | IC50: 2.92 ± 0.68 μM | DPP-IV GloTM Protease Assay | 50% inhibition | H Bonds and π interactions |
| Genistein [207] | IC50: 0.48 ± 0.04 μM | DPP-IV GloTM Protease Assay | 50% inhibition | H Bonds and π interactions |
| Resveratrol [207,218] | IC50: 0.0006 ± 0.0004 μM | DPP-IV GloTM Protease Assay | 50% inhibition | H Bonds |
| Gallic acid [207] | IC50: 4.65 ± 0.99 μM | DPP-IV GloTM Protease Assay | 50% inhibition | H Bonds and π interactions |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Alkhalidy, H.; Liu, D. The Emerging Role of Polyphenols in the Management of Type 2 Diabetes. Molecules 2021, 26, 703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030703
AMA Style
Wang Y, Alkhalidy H, Liu D. The Emerging Role of Polyphenols in the Management of Type 2 Diabetes. Molecules. 2021; 26(3):703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030703
Chicago/Turabian StyleWang, Yao, Hana Alkhalidy, and Dongmin Liu. 2021. "The Emerging Role of Polyphenols in the Management of Type 2 Diabetes" Molecules 26, no. 3: 703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26030703