
Preservative Effect of Aqueous and Ethanolic Extracts of the Macroalga Bifurcaria bifurcata on the Quality of Chilled Hake (Merluccius merluccius)



Abstract
:1. Introduction
2. Results and Discussion
2.1. Evolution of Microbial Development during Chilled Storage of Hake
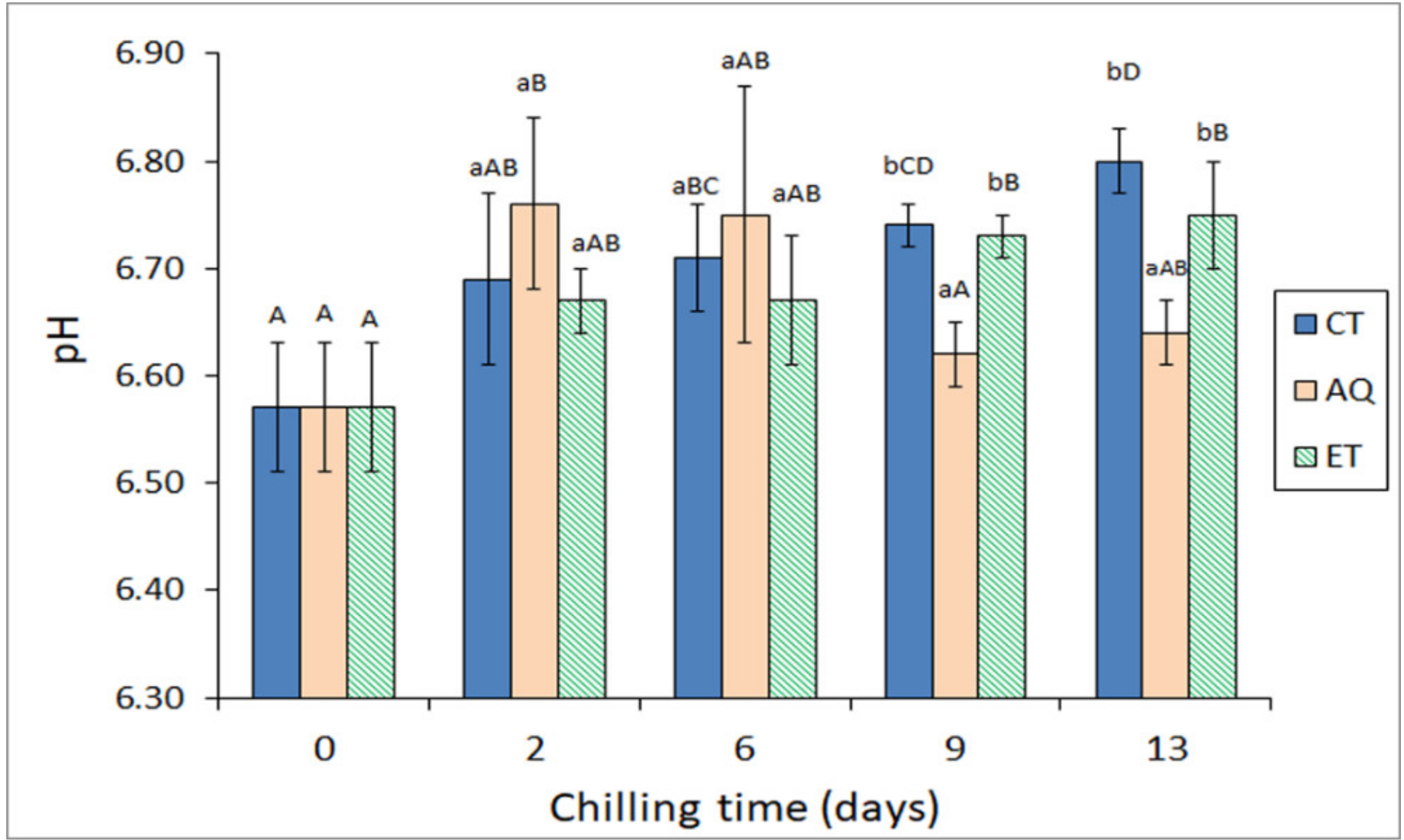
2.2. Comparative Evolution of pH Value and Free Fatty Acid (FFA) Content during Chilled Storage of Hake
2.3. Lipid Oxidation Development during Chilled Storage of Hake
3. Materials and Methods
3.1. Starting Macroalga B. bifurcata and Preparation of Icing Systems
3.2. Evolution of Microbial Development during Chilled Storage of Hake
3.3. Determination of Microbial Development
3.4. Assessment of Chemical Indices Related to Quality Loss
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Roohinejad, S.; Koubaa, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Grenier, R. Applications of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Int. 2017, 99, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rubio, M.E.; Pérez-Jiménez, J.; Saura-Calixto, F. Dietary fiber and antioxidant capacity in Fucus vesiculosus products. Int. J. Food Sci. Nutr. 2009, 60, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Ferreira Patarra, R.; Neto, A.; Baptista, J. Edible Azorean macroalgae as source of rich nutrients with impact on human health. Food Chem. 2014, 164, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Sandsdalen, E.; Haug, T.; Stensvag, K.; Styrvold, O. The antibacterial effect of a polyhydroxylated fucophlorethol from the marine brown alga, Fucus vesiculosus. World J. Microb. Biotechnol. 2003, 19, 777–782. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jonsdóttir, R.; Olafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Halldorsdóttir, S.; Sveinsdóttir, H.; Gudmundsdóttir, A.; Thorkelsson, G.; Kristinsson, H. High quality fish protein hydrolysates prepared from by-product material with Fucus vesiculosus extract. J. Funct. Foods 2014, 9, 10–17. [Google Scholar] [CrossRef]
- Campos, C.A.; Gliemmo, M.; Aubourg, S.P.; Barros-Velázquez, J. Novel technologies for the preservation of chilled aquatic food products. In Novel Technologies in Food Science; McElhatton, A., Amaral Sobral, P., Eds.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Pastoriza, L.; Bernárdez, M.; Sampedro, G.; Cabo, M.; Herrera, J. The use of water and ice with bactericide to prevent onboard and onshore spoilage of refrigerated megrim (Lepidorhombus whiffiagonis). Food Chem. 2008, 110, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Kuley, E.; Özoğul, F.; Polat, A. Advances in packaging. In Innovative Technologies in Seafood Processing; Özoğul, Y., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2020. [Google Scholar]
- Ceylan, Z.; Özoğul, Y. Irradiation Technology. In Innovative Technologies in Seafood Processing; Özoğul, Y., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2020. [Google Scholar]
- Aubourg, S.P. Impact of high-pressure processing on chemical constituents and nutritional properties in aquatic foods: A review. Int. J. Food Sci. Technol. 2018, 53, 873–891. [Google Scholar] [CrossRef]
- Mei, J.; Ma, X.; Xie, J. Review on natural preservatives for extending fish shelf life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [Green Version]
- Sanjuás-Rey, M.; García-Soto, B.; Fuertes-Gamundi, J.R.; Aubourg, S.P.; Barros-Velázquez, J. Effect of a natural organic acid-icing system on the microbiological quality of commercially relevant chilled fish species. LWT Food Sci. Technol. 2012, 46, 217–223. [Google Scholar]
- García-Soto, B.; Aubourg, S.P.; Calo-Mata, P.; Barros-Velázquez, J. Extension of the shelf-life of chilled-hake (Merluccius merluccius) by a novel icing medium containing natural organic acids. Food Cont. 2013, 34, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Quitral, V.; Donoso, M.L.; Ortiz, J.; Herrera, M.V.; Araya, H.; Aubourg, S.P. Chemical changes during the chilled storage of Chilean jack mackerel (Trachurus murphyi): Effect of a plant extract-icing system. LWT Food Sci. Technol. 2009, 42, 1450–1454. [Google Scholar] [CrossRef]
- Özyurt, G.; Kuley, E.; Balikçi, E.; Kaçar, Ç.; Gökdogan, S.; Etyemez, M.; Özogul, F. Effect of the icing with rosemary extract on the oxidative stability and biogenic amine formation in sardine (Sardinella aurita) during chilled storage. Food Bioprocess Technol. 2012, 5, 2777–2786. [Google Scholar] [CrossRef]
- European Council Regulation. European Community (EC), No 258/97, 27 January 1997 concerning novel foods and novel food ingredients. Off. J. Eur. Commun. 1997, 40, 1–7. [Google Scholar]
- Miranda, J.; Ortiz, J.; Barros-Velázquez, J.; Aubourg, S.P. Quality enhancement of chilled fish by including alga Bifurcaria bifurcata extract in the icing medium. Food Bioprocess Technol. 2016, 9, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Trigo, M.; López, M.; Dovale, G.; Ortiz, J.; Rodríguez, A.; Aubourg, S.P. Enhancement of sensory acceptance of frozen mackerel by alga-extract glazing. Bulg. Chem. Commun. 2019, 51, 216–223. [Google Scholar]
- Ortiz, J.; Vivanco, J.P.; Aubourg, S.P. Lipid and sensory quality of canned Atlantic salmon (Salmo salar): Effect of the use of different seaweed extracts as covering liquids. Eur. J. Lipid Sci. Technol. 2014, 116, 596–605. [Google Scholar] [CrossRef]
- Miranda, J.M.; Carrera, M.; Barros-Velázquez, J.; Aubourg, S.P. Impact of previous active dipping in Fucus spiralis extract on the quality enhancement of chilled lean fish. Food Control 2018, 90, 407–414. [Google Scholar] [CrossRef]
- Tierney, M.; Smyth, T.; Rai, D.; Soler-Vila, A.; Croft, A.; Brunton, N. Enrichment of phenol contents and antioxidant activity of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef]
- Babakhani, A.; Farvin, K.; Jacobsen, C. Antioxidative effect of seaweed extracts in chilled storage of minced Atlantic mackerel (Scomber scombrus): effect on lipid and protein oxidation. Food Bioprocess Technol. 2016, 9, 352–364. [Google Scholar] [CrossRef] [Green Version]
- Farvin, K.; Jacobsen, C. Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.; Critchley, A.; van de Velde, F.; Ribeiro-Claro, P. Identification of selected seaweed polysaccharides (phycocolloides) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Kuda, T.; Ikemori, T. Minerals, polysaccharides and antioxidant properties of aqueous solutions obtained from macroalgal beach-casts in the Noto Peninsula, Ishikawa, Japan. Food Chem. 2009, 112, 575–581. [Google Scholar] [CrossRef]
- Barros-Velázquez, J.; Miranda, J.M.; Ezquerra-Brauer, J.M.; Aubourg, S.P. Impact of icing systems with aqueous, ethanolic and ethanolic-aqueous extracts of alga Fucus spiralis on microbial and biochemical quality of chilled hake (Merluccius merluccius). Int. J. Food Sci. Technol. 2016, 51, 2081–2089. [Google Scholar] [CrossRef] [Green Version]
- Le Lann, K.; Jégou, C.; Stiger-Pouvreau, V. Effect of different conditioning treatments on total phenolic content and antioxidant activities in two Sargassacean species: Comparison of the frondose Sargassum muticum (Yendo) Fensholt and the cylindrical Bifurcaria bifurcata R. Ross. Phycol. Res. 2008, 56, 238–245. [Google Scholar] [CrossRef]
- Braune, W. Meeresalgen. Ein Farbbilführer zu den Verbreiteten Benthischen Grün-Braun- und Rotalgen der Weltmeere; ARG Gantner Verlag: Ruggel, Liechtenstein, 2008; pp. 1–596. [Google Scholar]
- Gómez-Ordoñez, E.; Jiménez-Escrig, A.; Rupérez, P. Dietary fibre and physicochemical properties of several edible seaweeds from the northwestern Spanish coast. Food Res. Int. 2010, 43, 2289–2294. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Rupérez, P. FTIR-ATR spectroscopy as a tool for polysaccharide identification in edible brown and red seaweeds. Food Hydrocol. 2011, 25, 1514–1520. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Domínguez, R.; Carballo, J.; Lorenzo, J.M. Phenolic compounds from three brown seaweed species using LC-DAD-ESI-MS/MS. Food Res. Int. 2017, 99, 979–985. [Google Scholar] [CrossRef]
- Zubia, M.; Fabre, M.S.; Kerjean, V.; Le Lann, K.; Stiger-Pouvreau, V.; Fauchon, M.; Deslandes, E. Antioxidant and antitumoral activities of some Phaeophyta from Britanny coasts. Food Chem. 2009, 116, 693–701. [Google Scholar] [CrossRef]
- Bouzidi, N.; Daghbouche, Y.; El Hattab, M.; Aliche, Z.; Culioli, G.; Piovetti, L.; Garrigues, S.; De la Guardia, M. Determination of total sterols in brown algae by Fourier transform infrared spectroscopy. Anal. Chim. Acta 2008, 616, 185–189. [Google Scholar] [CrossRef]
- Alves, C.; Pinteus, S.; Simoes, T.; Horta, A.; Silva, J.; Tecelão, C.; Pedrosa, R. Bifurcaria bifurcata: A key macro-alga as a source of bioactive compounds and functional ingredients. Int. J. Food Sci. Technol. 2016, 51, 1638–1646. [Google Scholar] [CrossRef]
- Broekaert, K.; Heyndrickx, M.; Herman, L.; Devlighere, F.; Vlaemynk, G. Seafood quality analysis: Molecular identification of dominant microbiota after ice storage on several general growth media. Food Microbiol. 2011, 18, 1162–1169. [Google Scholar] [CrossRef]
- Smit, A. Medicinal and pharmaceutical uses of seaweed natural products: A Review. J. App. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Glombitza, K.; Rösener, H.; Koch, M. Polyhydroxyoligophenyle und phenyläther aus Bifurcaria bifurcata. Phytochemistry 1976, 15, 1279–1281. [Google Scholar] [CrossRef]
- Culioli, G.; Daoudi, M.; Ortalo-Magne, A.; Valls, R.; Piovetti, L. (S)-12-hydroxygeranylgeraniol-derived diterpenes from the brown alga Bifurcaria bifurcata. Phytochemistry 2001, 57, 529–535. [Google Scholar] [CrossRef]
- Campos, C.A.; Miranda, J.M.; Trigo, M.; Barros-Velázquez, J.; Aubourg, S.P. Effect of alga Undaria pinnatifida (“wakame”) extract on the quality evolution of chilled megrim (Lepidorhombus whiffiagonis). Bulg. Chem. Commun. 2019, 51, 137–143. [Google Scholar]
- Bensid, A.; Ucar, Y.; Bendeddouche, B.; Özogul, F. Effect of the icing with thyme, oregano and clove extracts on quality parameters of gutted and beheaded anchovy (Engraulis encrasicholus) during chilled storage. Food Chem. 2014, 145, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Viji, P.; Binsi, P.K.; Visnuvinayagam, S.; Ravishankar, C.N.; Venkateshwarlu, G.; Gopal, T.K.S. Modified icing system containing mint leaf and citrus peel extracts: Effects on quality changes and shelf life of Indian mackerel. Indian J. Fish. 2016, 63, 93–101. [Google Scholar] [CrossRef]
- Apang, T.; Martin Xavier, K.A.; Lekshmi, M.; Kannuchamy, N.; Layana, P.; Balange, A.K. Garcinia spp. extract incorporated icing medium as a natural preservative for shelf life enhancement of chilled Indian mackerel (Rastrelliger kanagurta). LWT Food Sci. Technol. 2020, 133, 110086. [Google Scholar] [CrossRef]
- Aubourg, S.P. Fluorescence study of the prooxidant activity of free fatty acids on marine lipids. J. Sci. Food Agric. 2001, 81, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J.; Morançais, M.; Dumay, J.; Decottignies, P.; Turpin, V.; Munier, M.; García-Bueno, N.; Jaouen, P. What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends Food Sci. Technol. 2012, 27, 57–61. [Google Scholar] [CrossRef]
- Burtin, P. Nutritional value of seaweeds. Electron. J. Environ. Agric. Food Chem. 2003, 2, 498–503. [Google Scholar]
- Agregán, R.; Munekata, P.E.; Dominguez, R.; Carballo, J.; Franco, D.; Lorenzo, J.M. Proximate composition, phenolic content and in vitro antioxidant activity of aqueous extracts of the seaweeds Ascophyllum nodosum, Bifurcaria bifurcata and Fucus vesiculosus. Effect of addition of the extracts on the oxidative stability of canola oil under accelerated storage conditions. Food Res. Int. 2017, 99, 986–994. [Google Scholar]
- Rodríguez, O.; Losada, V.; Aubourg, S.P.; Barros-Velázquez, J. Sensory, microbial and chemical effects of a slurry ice system on horse mackerel (Trachurus trachurus). J. Sci. Food Agric. 2005, 85, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.; Dyer, W. A rapid method of total extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Lowry, R.; Tinsley, I. Rapid colorimetric determination of free fatty acids. J. Am. Oil Chem. Soc. 1976, 53, 470–472. [Google Scholar] [CrossRef]
- Chapman, R.; McKay, J. The estimation of peroxides in fats and oils by the ferric thiocyanate method. J. Am. Oil Chem. Soc. 1949, 26, 360–363. [Google Scholar] [CrossRef]
- Vyncke, W. Direct determination of the thiobarbituric acid value in trichloracetic acid extracts of fish as a measure of oxidative rancidity. Fette Seifen Anstrichm. 1970, 72, 1084–1087. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Microbial Group | Chilling Time (Days) | Icing Condition | ||
|---|---|---|---|---|
| CT | AQ | ET | ||
| Aerobes | 0 | 2.52 A (0.24) | 2.52 A (0.24) | 2.52 A (0.24) |
| 2 | 2.35 aA (0.37) | 2.77 aAB (0.81) | 2.20 aA (0.17) | |
| 6 | 3.92 aB (0.15) | 3.71 aB (0.74) | 4.04 aB (0.52) | |
| 9 | 5.28 abC (0.51) | 5.09 aC( (0.46) | 5.95 bC (0.32) | |
| 13 | 5.71 aC (0.59) | 5.44 aC (0.25) | 5.61 aC (0.48) | |
| Psychrotrophs | 0 | 3.38 A (0.41) | 3.38 A (0.41) | 3.38 B (0.41) |
| 2 | 3.20 bA (0.13) | 3.56 bA (0.26) | 2.20 aA (0.35) | |
| 6 | 5.44 aB (0.24) | 5.34 aB (0.03) | 5.40 aC (0.28) | |
| 9 | 6.30 aC (0.45) | 5.91 aC (0.34) | 5.97 aC (0.47) | |
| 13 | 7.81 bD (0.35) | 6.80 aD (0.23) | 6.97 aD (0.46) | |
| Enterobacteria-ceae *** | 0 | 1 (0.0) | 1 (0.0) | 1 (0.0) |
| 2 | 1 (0.0) | 1 (0.0) | 1 (0.0) | |
| 6 | 1 (0.0) | 1 (0.0) | 1 (0.0) | |
| 9 | 1 (0.0) | 1.10 (0.17) | 1.66 (0.58) | |
| 13 | 1.36 (0.32) | 1 (0.0) | 1 (0.0) | |
| Proteolytics | 0 | 2.40 A 0.46) | 2.40 A (0.46) | 2.40 A (0.46) |
| 2 | 2.20 aA 0.17) | 2.42 aA (0.39) | 2.00 aA (0.33) | |
| 6 | 3.44 aB (0.10) | 3.32 aB (0.15) | 3.55 aB (0.33) | |
| 9 | 5.29 aC (0.08) | 5.27 abC (0.49) | 5.64 bC (0.03) | |
| 13 | 6.78 aD (0.47) | 6.55 aD (0.23) | 6.26 aD (0.43) | |
| Extraction Medium | Bioactive Compound | Reference |
|---|---|---|
| aq. 80% ethanol | phenols | Glombitza et al. 1976 [39] |
| ethyl ether | diterpenes | Culioli et al. 2001 [40] |
| methanol:chloroform (1:1) | Sterols, i.e., fucosterol | Bouzidi et al. 2008 [35] |
| methanol | polyphenols | Alves et al. 2016 [36] |
| water | polysaccharides, i.e., alginate | Gómez-Ordóñez and Rupérez, 2011 [32] |
| water | total dietary fibre; soluble and insoluble fibre | Gómez-Ordóñez et al. 2010 [31] |
| water | pholorotannins, phenmolic acids, flavonoids, fuhalols, hydroxyl-fuhalols, eckol derivatives, and rosmarinic acid | Agregán et al. 2017 [33] |
| Chemical Parameter | Chilling Time (Days) | Icing Condition | ||
|---|---|---|---|---|
| CT | AQ | ET | ||
| Free fatty acids (mg·kg−1 muscle) | 0 | 64.38 A (6.52) | 64.38 A (6.58) | 64.38 A (6.56) |
| 2 | 78.88 cB (1.98) | 66.75 bA (1.36) | 58.41 aA (2.07) | |
| 6 | 94.70 cC (3.41) | 87.58 bB (2.86) | 70.21 aB (1.79) | |
| 9 | 113.67 bD (7.19) | 102.72 bC (5.44) | 95.97 aC (1.89) | |
| 13 | 139.50 bE (6.17) | 105.88 aC (7.97) | 93.23 aC (3.95) | |
| Peroxide value (meq. active oxygen·kg−1 lipids) | 0 | 0.14 A (0.00) | 0.14 A (0.00) | 0.14 A (0.00) |
| 2 | 2.28 aB (0.15) | 2.69 aB (0.55) | 3.75 bB (0.50) | |
| 6 | 2.50 aB (0.07) | 2.45 aB (0.71) | 3.50 bB (0.18) | |
| 9 | 3.80 aB (2.62) | 2.66 aB (1.05) | 7.28 bC (1.14) | |
| 13 | 4.37 aB (2.67) | 3.69 aB (0.95) | 7.24 bC (0.57) | |
| Thiobarbituric acid index (mg malondialde-hyde·kg−1 muscle) | 0 | 0.13 A (0.09) | 0.13 A (0.09) | 0.13 A (0.09) |
| 2 | 0.35 bB (0.09) | 0.17 aA (0.06) | 0.26 abAB (0.06) | |
| 6 | 0.36 aAB (0.15) | 0.35 aB (0.10) | 0.48 aD (0.05) | |
| 9 | 0.37 aB (0.05) | 0.51 a (0.07) | 0.45 aCD (0.05) | |
| 13 | 0.70 bC (0.07) | 0.41 aB (0.07) | 0.36 aBC (0.05) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, J.M.; Zhang, B.; Barros-Velázquez, J.; Aubourg, S.P. Preservative Effect of Aqueous and Ethanolic Extracts of the Macroalga Bifurcaria bifurcata on the Quality of Chilled Hake (Merluccius merluccius). Molecules 2021, 26, 3774. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123774
Miranda JM, Zhang B, Barros-Velázquez J, Aubourg SP. Preservative Effect of Aqueous and Ethanolic Extracts of the Macroalga Bifurcaria bifurcata on the Quality of Chilled Hake (Merluccius merluccius). Molecules. 2021; 26(12):3774. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123774
Chicago/Turabian StyleMiranda, José M., Bin Zhang, Jorge Barros-Velázquez, and Santiago P. Aubourg. 2021. "Preservative Effect of Aqueous and Ethanolic Extracts of the Macroalga Bifurcaria bifurcata on the Quality of Chilled Hake (Merluccius merluccius)" Molecules 26, no. 12: 3774. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123774