
Structural, Mechanistic, and Functional Insights into an Arthrobacter nicotinovorans Molybdenum Hydroxylase Involved in Nicotine Degradation

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
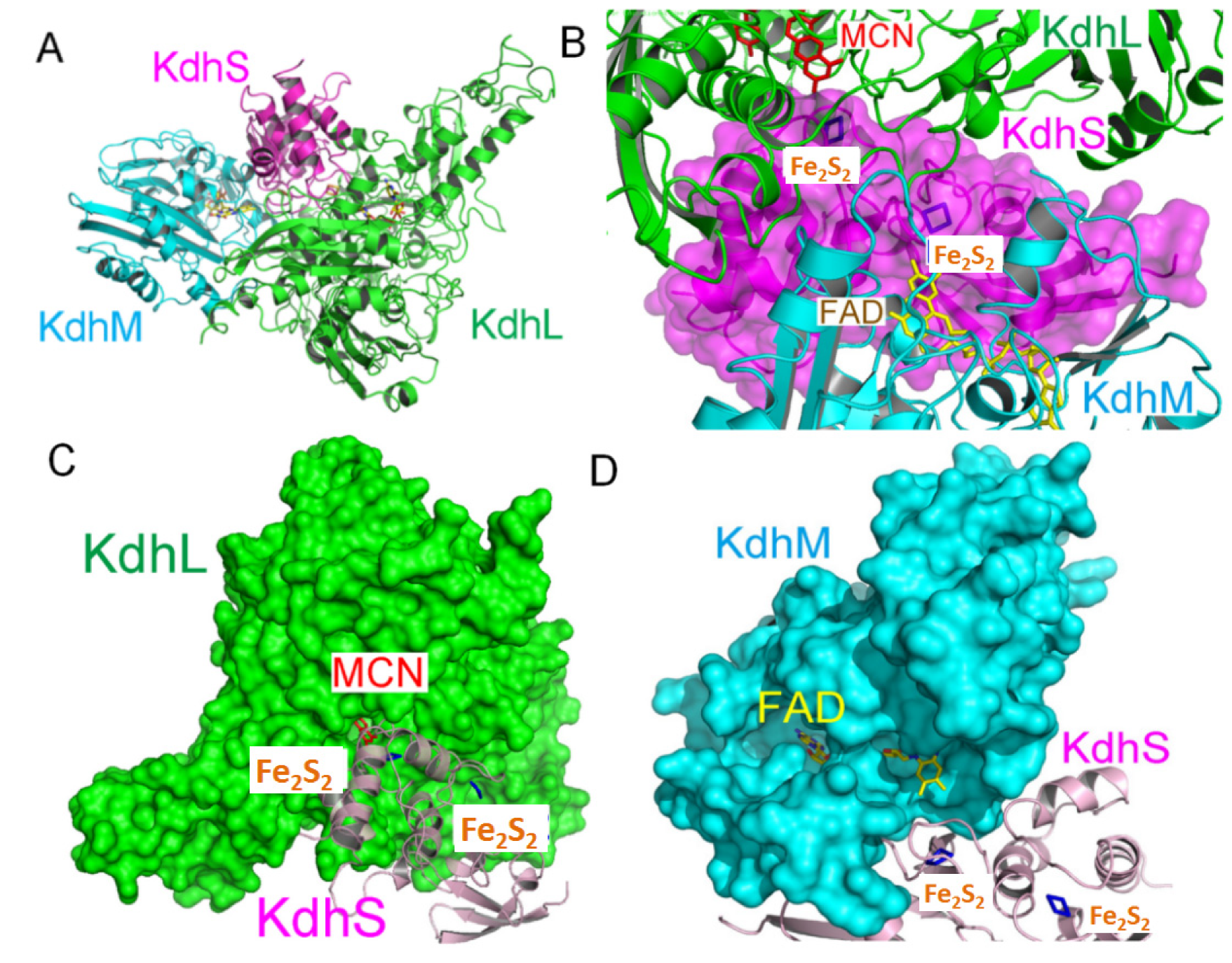
2.1. Overall Structure of the Kdh Holoenzyme
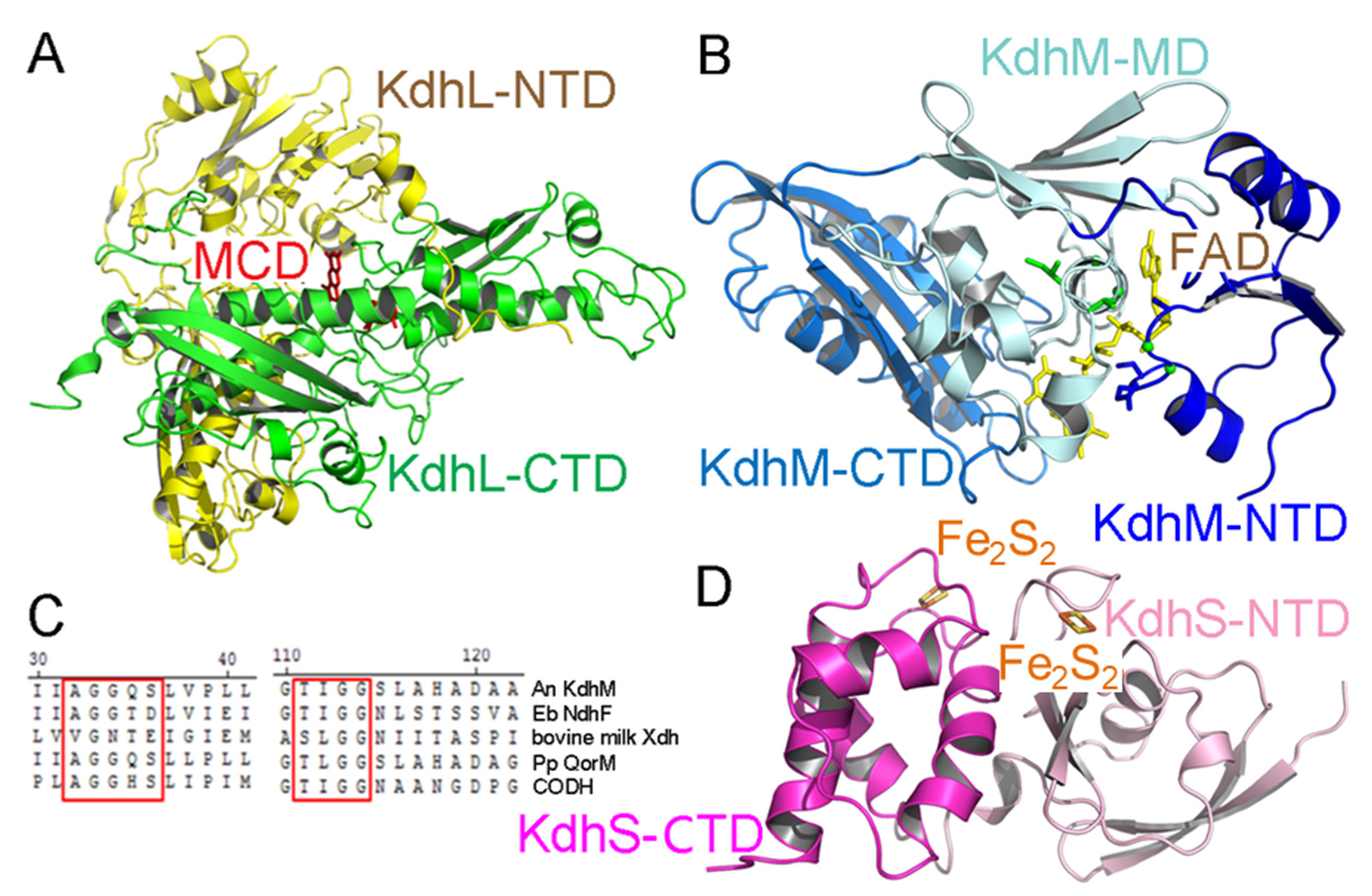
2.2. Structure of Each Subunit of Kdh
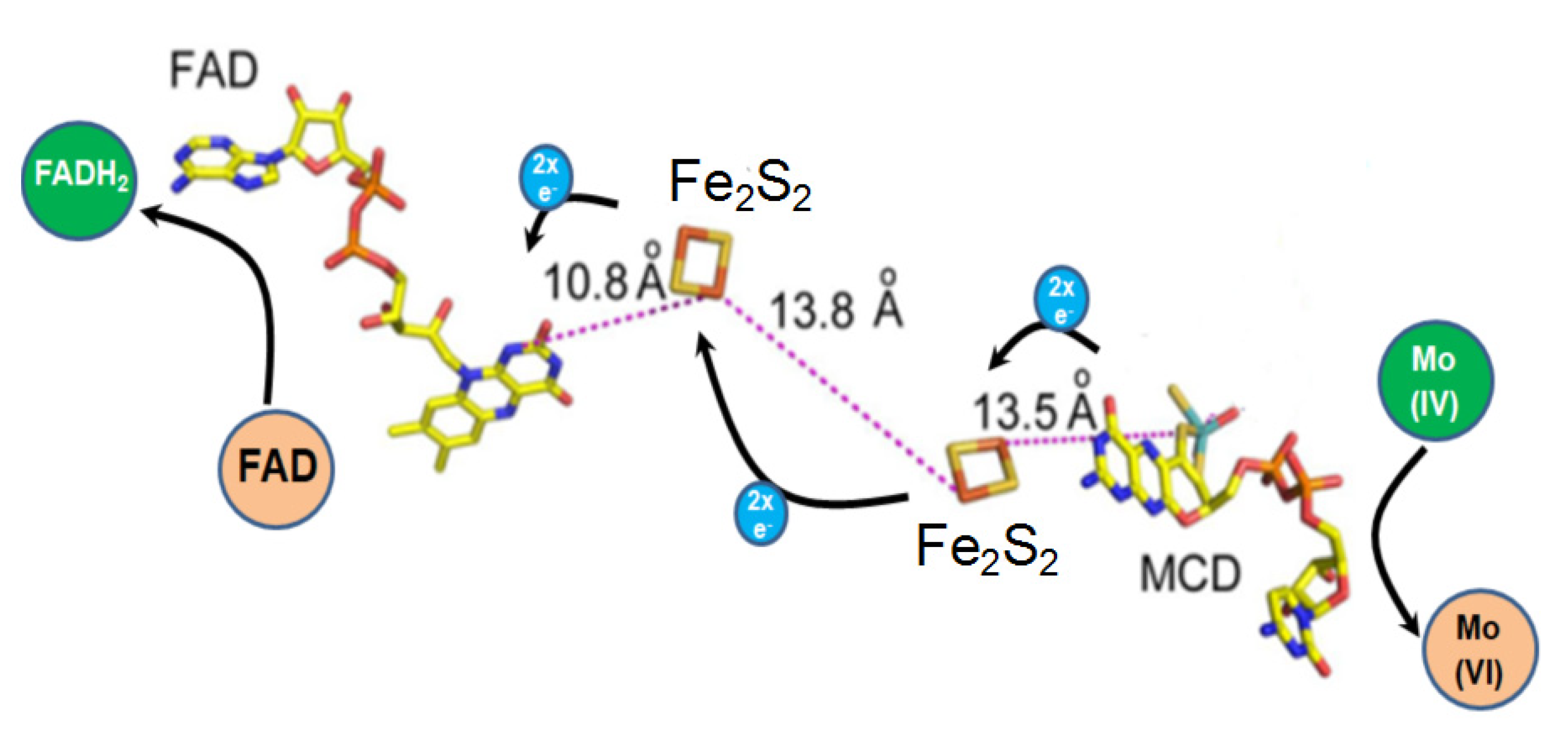
2.3. The Cofactors of Kdh Are Arranged in a Linear Fashion
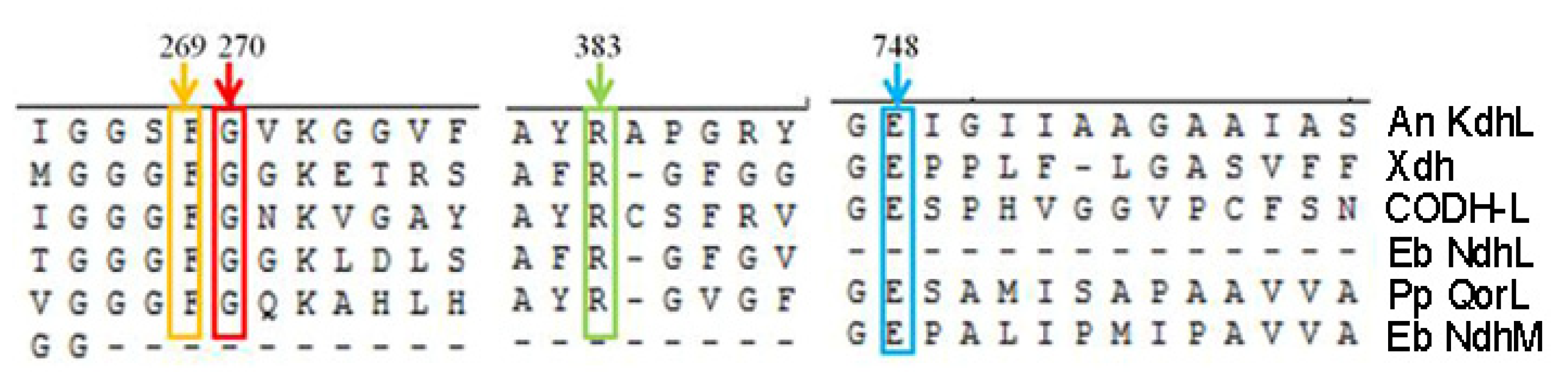
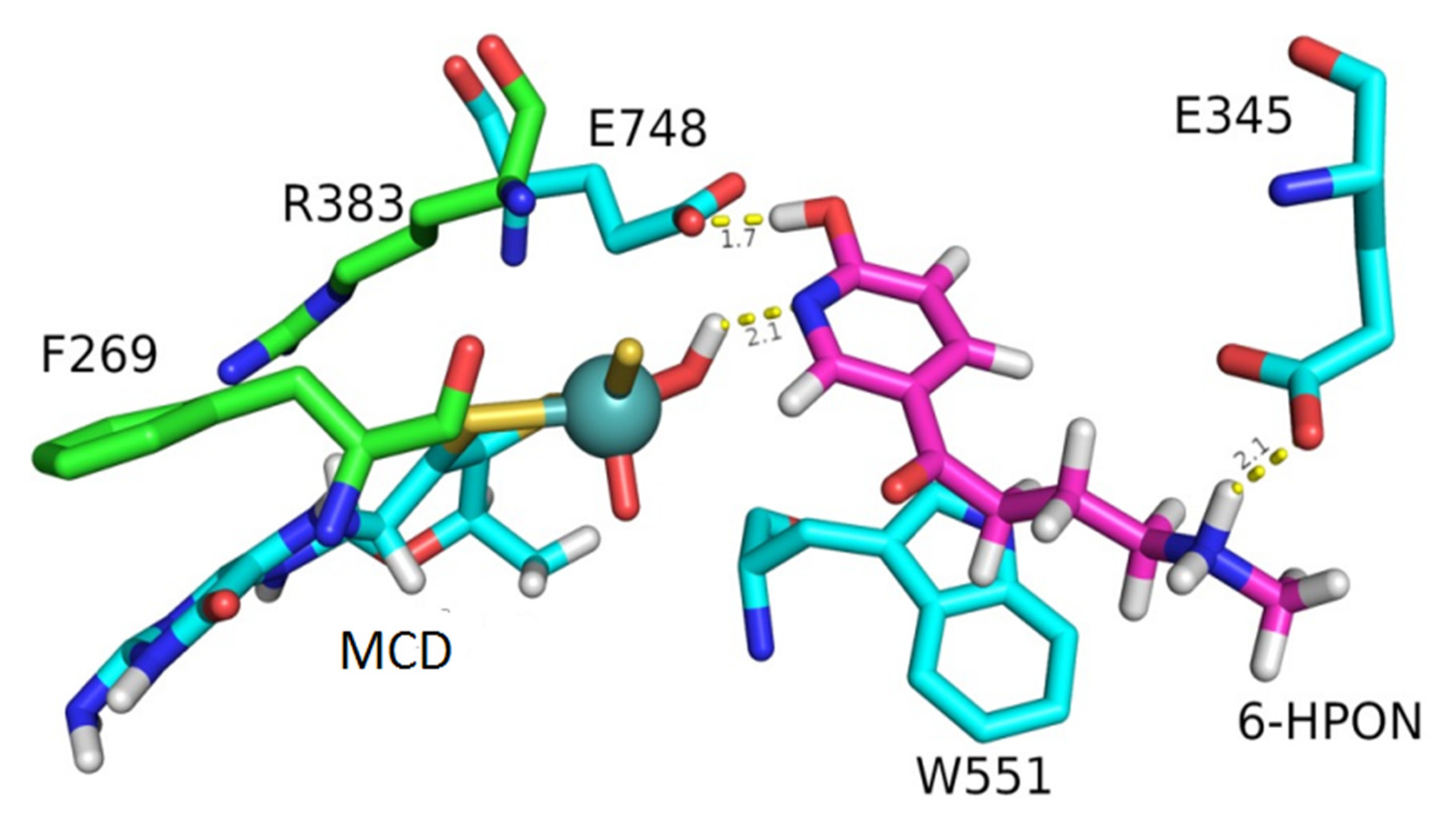
2.4. The Substrate Channel of KdhL
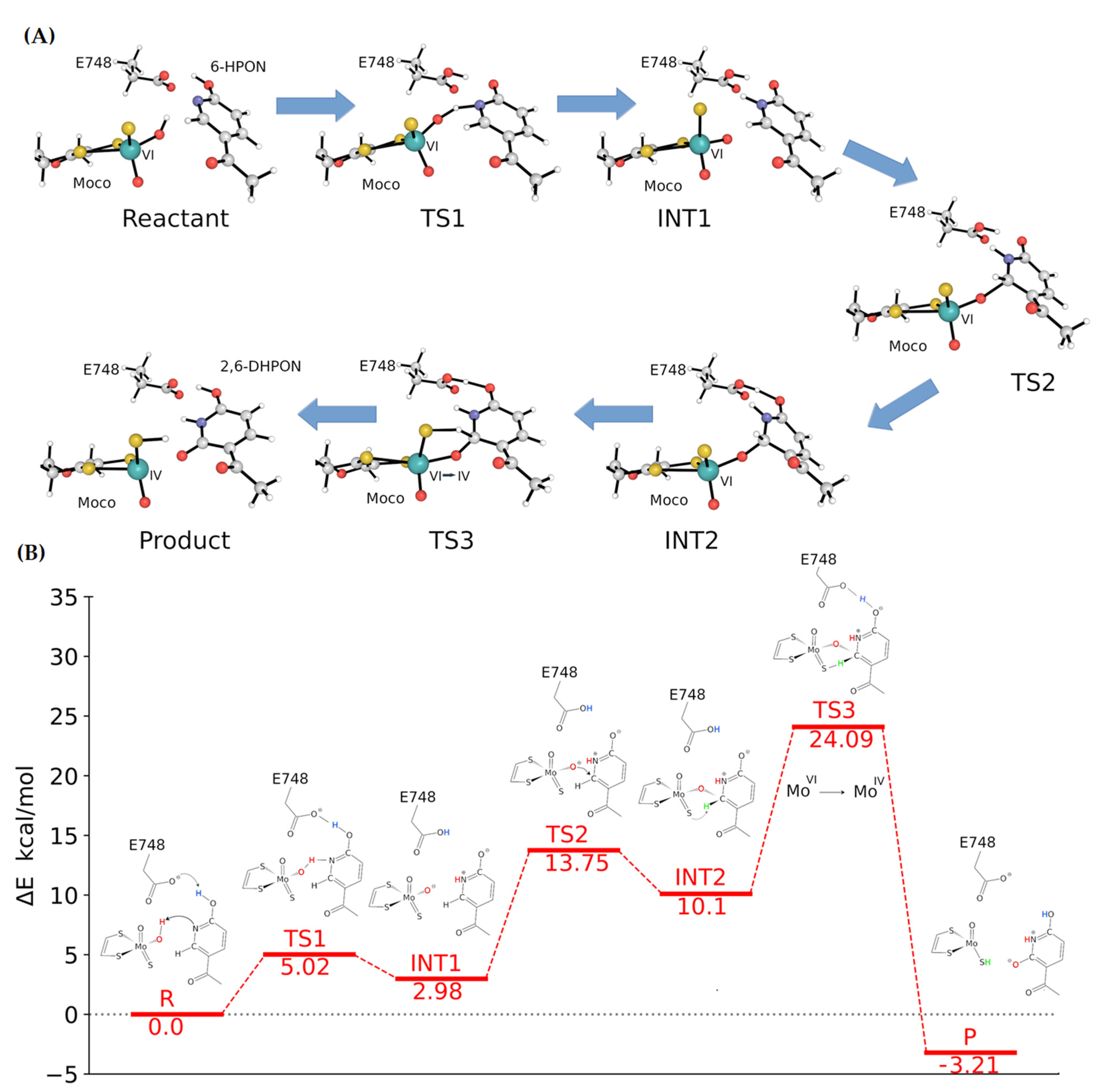
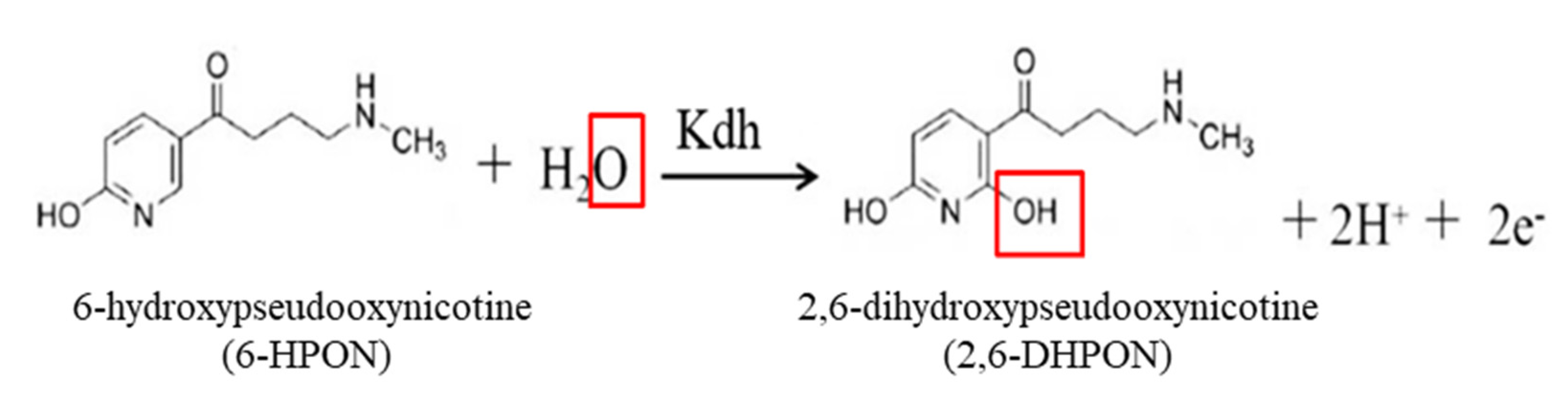
2.5. Catalytic Mechanism of Kdh
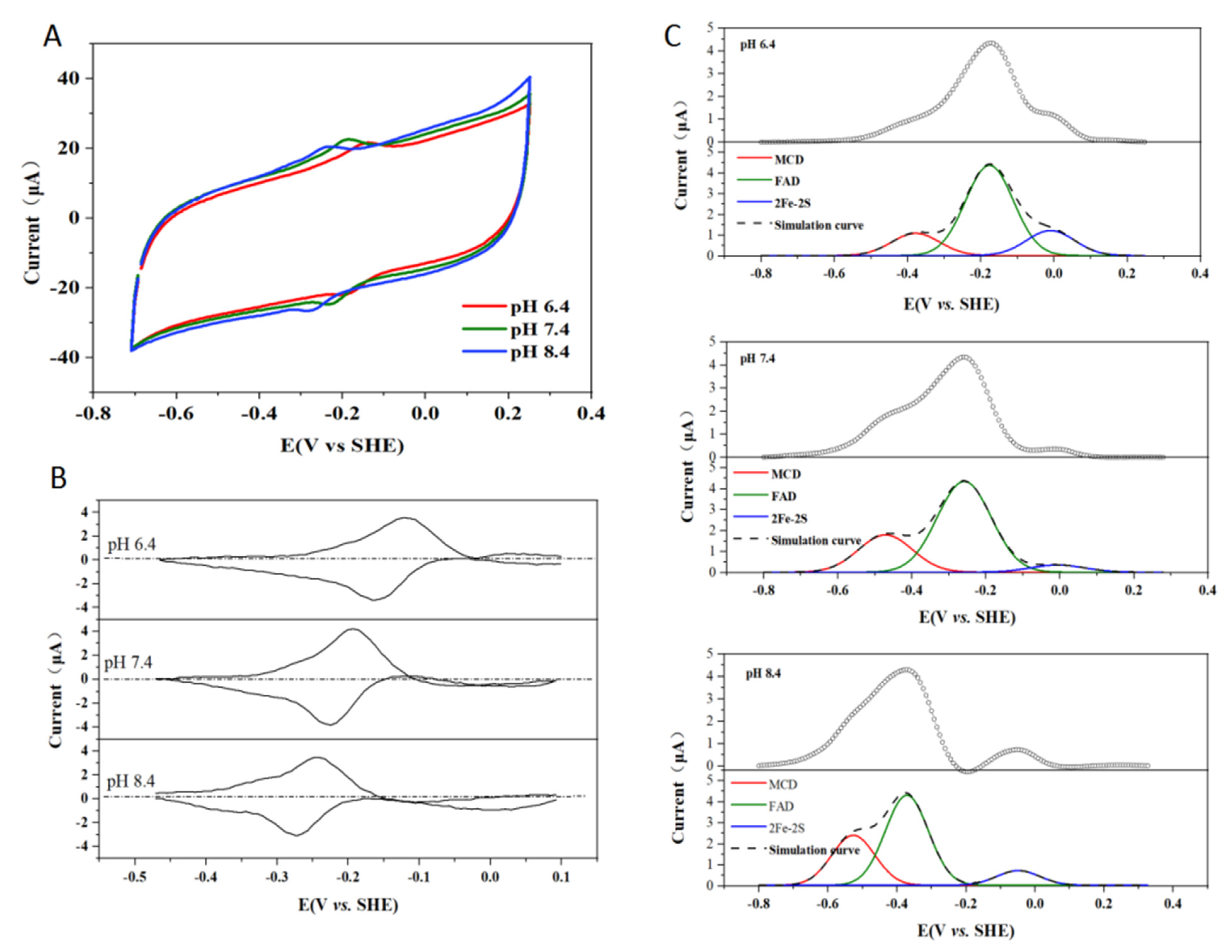
2.6. Effect of pH on the Enzymatic Activity of Kdh
3. Materials and Methods
3.1. Cloning and Purification
3.2. Data Collection and Structure Determination
3.3. Construction of Mutants
3.4. Enzyme Activity Assays
3.5. Electrochemical Measurement
3.6. MD Simulation and QM/MM Calculation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Accession Code
References
- Xu, P.; Yu, B.; Li, F.L.; Cai, X.F.; Ma, C.Q. Microbial degradation of sulfur, nitrogen and oxygen heterocycles. Trends Microbiol. 2006, 14, 398–405. [Google Scholar] [CrossRef]
- Doolittle, D.J.; Winegar, R.; Lee, C.K.; Caldwell, W.S.; Hayes, A.W.; de Bethizy, J.D. The genotoxic potential of nicotine and its major metabolites. Mutat. Res. 1995, 344, 95–102. [Google Scholar] [CrossRef]
- Curwin, B.D.; Hein, M.J.; Sanderson, W.T.; Nishioka, M.G.; Buhler, W. Nicotine exposure and decontamination on tobacco harvesters’ hands. Ann. Occup. Hyg. 2005, 49, 407–413. [Google Scholar]
- Wu, G.; Xu, P.; Chen, D.; Tang, H.; Ren, Y.; Chen, Q.; Lv, Y.; Zhang, Z.; Zhao, Y.L. Structural insights into the specific recognition of N-heterocycle biodenitrogenation-derived substrates by microbial amide hydrolases. Mol. Microbiol. 2014, 91, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Torres Pazmino, D.E.; Winkler, M.; Glieder, A.; Fraaije, M.W. Monooxygenases as biocatalysts: Classification, mechanistic aspects and biotechnological applications. J. Biotechnol. 2010, 146, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, X.; Liu, X.; Wu, W.; Xu, P.; Tang, H. Cloning and characterization the nicotine degradation enzymes 6-hydroxypseudooxynicotine amine oxidase and 6-hydroxy-3-succinoylpyridine hydroxylase in Pseudomonas geniculata N1. Int. Biodeter. Biodegr. 2019, 142, 83–90. [Google Scholar] [CrossRef]
- Liu, G.; Wang, W.; He, F.; Zhang, P.; Xu, P.; Tang, H. Structural insights into 6-hydroxypseudooxynicotine amine oxidase from Pseudomonas geniculata N1: The key enzyme involved in nicotine degradation. Appl. Environ. Microbiol. 2020, 86, e01559-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Yin, B.; Peng, X.X.; Wang, J.Y.; Xie, Z.H.; Gao, J.; Tang, X.K. Biodegradation of nicotine by newly isolated Pseudomonas sp. CS3 and its metabolites. J. Appl. Microbiol. 2012, 112, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Brandsch, R. Microbiology and biochemistry of nicotine degradation. Appl. Microbiol. Biotechnol. 2006, 69, 493–498. [Google Scholar] [CrossRef]
- Igloi, G.L.; Brandsch, R. Sequence of the 165-kilobase catabolic plasmid pAO1 from Arthrobacter nicotinovorans and identification of a pAO1-dependent nicotine uptake system. J. Bacteriol. 2003, 185, 1976–1986. [Google Scholar] [CrossRef] [Green Version]
- Decker, K.; Bleeg, H. Induction and purification of stereospecific nicotine oxidizing enzymes from Arthrobacter oxidans. Biochim. Biophys. Acta. 1965, 105, 313–324. [Google Scholar] [CrossRef]
- Truglio, J.J.; Theis, K.; Leimkuhler, S.; Rappa, R.; Rajagopalan, K.V.; Kisker, C. Crystal structures of the active and alloxanthine-inhibited forms of xanthine dehydrogenase from Rhodobacter capsulatus. Structure 2002, 10, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, K.; Matsumoto, K.; Hille, R.; Eger, B.T.; Pai, E.F.; Nishino, T. The crystal structure of xanthine oxidoreductase during catalysis: Implications for reaction mechanism and enzyme inhibition. Proc. Natl. Acad. Sci. USA 2004, 101, 7931–7936. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.W.; Kirk, M.L. Electronic structure contributions to reactivity in xanthine oxidase family enzymes. J. Biol. Inorg. Chem. 2015, 20, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, I.; Martins, B.M.; Purvanov, V.; Fetzner, S.; Huber, R.; Dobbek, H. Active site geometry and substrate recognition of the molybdenum hydroxylase quinoline 2-oxidoreductase. Structure 2004, 12, 1425–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagener, N.; Pierik, A.J.; Ibdah, A.; Hille, R.; Dobbek, H. The Mo-Se active site of nicotinate dehydrogenase. Proc. Natl. Acad. Sci. USA 2009, 106, 11055–11060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbek, H.; Gremer, L.; Meyer, O.; Huber, R. Crystal structure and mechanism of CO dehydrogenase; a molybdo iron-sulfur flavoprotein containing S-selanylcysteine. Proc. Natl. Acad. Sci. USA 1999, 96, 8884–8889. [Google Scholar] [CrossRef] [Green Version]
- Baitsch, D.; Sandu, C.; Brandsch, R.; Igloi, G.L. Gene cluster on pAO1 of Arthrobacter nicotinovorans involved in degradation of the plant alkaloid nicotine: Cloning; purification; and characterization of 2;6-dihydroxypyridine 3-hydroxylase. J. Bacteriol. 2001, 183, 5262–5267. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, P.; Cerqueira, N.M.F.S.A.; Fernandes, P.A.; Romao, M.J.; Ramos, M.J. Catalytic mechanism of human aldehyde oxidase. Acs. Catal. 2020, 10, 9276–9286. [Google Scholar] [CrossRef]
- Hanzelmann, P.; Dobbek, H.; Gremer, L.; Huber, R.; Meyer, O. The effect of intracellular molybdenum in Hydrogenophaga pseudoflava on the crystallographic structure of the seleno-molybdo-iron-sulfur flavoenzyme carbon monoxide dehydrogenase. J. Mol. Biol. 2000, 301, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Schulz, G.E. Binding of nucleotides by proteins. Curr. Biol. 1992, 2, 61–67. [Google Scholar] [CrossRef]
- Dobbek, H.; Gremer, L.; Kiefersauer, R.; Huber, R.; Meyer, O. Catalysis at a dinuclear [CuSMo(==O)OH] cluster in a CO dehydrogenase resolved at 1.1-A resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 15971–15976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, C.C.; Moser, C.C.; Chen, X.X.; Dutton, P.L. Natural engineering principles of electron tunnelling in biological oxidation-reduction. Nature 1999, 402, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Neese, F.; Wennmohs, F.; Becker, U.; Riplinger, C. The ORCA quantum chemistry program package. J. Chem. Phys. 2020, 152, 224108. [Google Scholar] [CrossRef] [PubMed]
- Hille, R.; Hall, J.; Basu, P. The mononuclear molybdenum enzymes. Chem. Rev. 2014, 114, 3963–4038. [Google Scholar] [CrossRef] [Green Version]
- Vidossich, P.; Lledos, A. The use of localised orbitals for the bonding and mechanistic analysis of organometallic compounds. Dalton Trans. 2014, 43, 11145–11151. [Google Scholar] [CrossRef]
- Hendrik, A.; Joel, H.; Fraser, A. Direct detection and measurement of electron relays in a multicentered enzyme: Voltammetry of electrode-surface films of E. coli fumarate reductase; an iron-sulfur flavoprotein. J. Am. Chem. Soc. 1997, 119, 11628–11638. [Google Scholar]
- Cristinel, S.; Calin-Bogdan, C.; Paula, S.; Roderich, B. Plasmids for nicotine-dependent and -independent gene expression in Arthrobacter nicotinovorans and other Arthrobacter species. Appl. Environ. Microbiol. 2005, 71, 8920–8924. [Google Scholar]
- Paula, S.; Emile, S.; Roderich, B. A Functional Mo bA Gene for molybdopterin cytosine dinucleotide cofactor biosynthesis is required for activity and holoenzyme assembly of the heterotrimeric nicotine dehydrogenases of Arthrobacter nicotinovorans. Appl. Environ. Microbiol. 2006, 72, 5126–5131. [Google Scholar]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [PubMed]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delano, W.L. The PyMOL Molecular Graphic System; Schrodinger: New York, NY, USA, 2002. [Google Scholar]
- Siddiqui, J.A.; Shoeb, S.M.; Takayama, S.; Shimizu, E.; Yorifuji, T. Purification and characterization of histamine dehydrogenase from Nocardioides simplex IFO 12069. FEMS Microbiol. Lett. 2000, 189, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.M.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09 Revision E.01; Gaussian Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Dupradeau, F.-Y.; Cézard, C.; Lelong, R.; Stanislawiak, É.; Pêcher, J.; Delepine, J.C.; Cieplak, P. R.E.DD.B.: A database for RESP and ESP atomic charges; and force field libraries. Nucleic Acids Res. 2007, 36, D360–D367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Merz, K.M. MCPB.py: A python based metal center parameter builder. J. Chem. Inf. Model. 2016, 56, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Yang, W.; Parr, R.G. Development of the Colle-Salvetti correlation-energy formula into a functional of the electron density. Phys. Rev. B Condens. Matter. 1988, 37, 785–789. [Google Scholar] [CrossRef] [Green Version]
- Andrae, D.; Häußermann, U.; Dolg, M.; Stoll, H.; Preuß, H. Energy-adjustedab initio pseudopotentials for the second and third row transition elements. Theor. Chim. Acta 1990, 77, 123–141. [Google Scholar] [CrossRef]
- Weigend, F.; Ahlrichs, R. Balanced basis sets of split valence; triple zeta valence and quadruple zeta valence quality for H to Rn: Design and assessment of accuracy. Phys. Chem. Chem. Phys. 2005, 7, 3297–3305. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | Kdh |
|---|---|
| Beamline | BL17U1 (SSRF) |
| Space group | P212121 |
| Unit cell parameter | |
| a, b, c (Å) | 112.86, 126.79, 294.49 |
| α, β, γ (°) | 90, 90, 90 |
| Resolution (Å) | 50.0–3.4 |
| Rmerge (outer shell) | 0.122 (0.693) |
| I/σI (outer shell) | 11.3 (1.0) |
| CC1/2 | 0.512 |
| Completeness (%) (outer shell) | 100 (100) |
| Redundancy (outer shell) | 12.2 (10.6) |
| Refinement | |
| Resolution (Å) | 147.2–3.4 |
| Number of reflections | 50,560 |
| Rwork/Rfree | 28.7%/32.6% |
| Number of atoms | 18676 |
| B factors (Å2) | 67.88 |
| RMSD bond length (Å) | 0.0097 |
| RMSD bond angles (°) | 1.7528 |
| Ramanchandran plot | |
| Favored (%) | 83.6 |
| Allowed (%) | 12.2 |
| Disallowed (%) | 4.1 |
| WT and Mutants | KM (μM) | kcat (s−1) | kcat/KM (s−1μM−1) |
|---|---|---|---|
| WT | 12.5 ± 0.9 | 9.03 ± 0.12 | 0.72 ± 0.13 |
| E748A | N.D. | N.D. | N.D. |
| F269A | 25.98 ± 2.73 | 2.30 ± 0.07 | 0.09 ± 0.03 |
| W551A | N.D. | N.D. | N.D. |
| R383A | 21.4 ± 2.3 | 6.32 ± 0.16 | 0.30 ± 0.07 |
| E345A | N.D. | N.D. | N.D. |
| Buffer | KM (μM) | kcat (s−1) | kcat/KM (s−1μM−1) |
|---|---|---|---|
| PBS, pH 6.4 | 64.67 ± 11.57 | 5.13 ± 0.41 | 0.08 ± 0.04 |
| PBS, pH 7.4 | 23.41 ± 1.76 | 10.58 ± 0.22 | 0.45 ± 0.13 |
| PBS, pH 8.0 | 12.5 ± 0.91 | 9.03 ± 0.12 | 0.72± 0.13 |
| Tris-HCl, pH 6.8 | 37.71 ± 2.14 | 9.09 ± 0.15 | 0.24 ± 0.07 |
| Tris-HCl, pH 7.4 | 22.29 ± 0.81 | 9.28 ± 0.08 | 0.42 ± 0.10 |
| Tris-HCl, pH 8.0 | 15.88 ± 1.08 | 9.47 ± 0.13 | 0.60 ± 0.12 |
| Tris-HCl, pH 8.5 | 15.09 ± 1.44 | 8.92 ± 0.28 | 0.59 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Mu, X.; Li, W.; Xu, Q.; Xu, P.; Zhang, L.; Zhang, Y.; Wu, G. Structural, Mechanistic, and Functional Insights into an Arthrobacter nicotinovorans Molybdenum Hydroxylase Involved in Nicotine Degradation. Molecules 2021, 26, 4387. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144387
Wang L, Mu X, Li W, Xu Q, Xu P, Zhang L, Zhang Y, Wu G. Structural, Mechanistic, and Functional Insights into an Arthrobacter nicotinovorans Molybdenum Hydroxylase Involved in Nicotine Degradation. Molecules. 2021; 26(14):4387. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144387
Chicago/Turabian StyleWang, Lei, Xia Mu, Wenjin Li, Qin Xu, Ping Xu, Liyun Zhang, Yuebin Zhang, and Geng Wu. 2021. "Structural, Mechanistic, and Functional Insights into an Arthrobacter nicotinovorans Molybdenum Hydroxylase Involved in Nicotine Degradation" Molecules 26, no. 14: 4387. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144387