
Aged Brains Express Less Melanocortin Receptors, Which Correlates with Age-Related Decline of Cognitive Functions

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
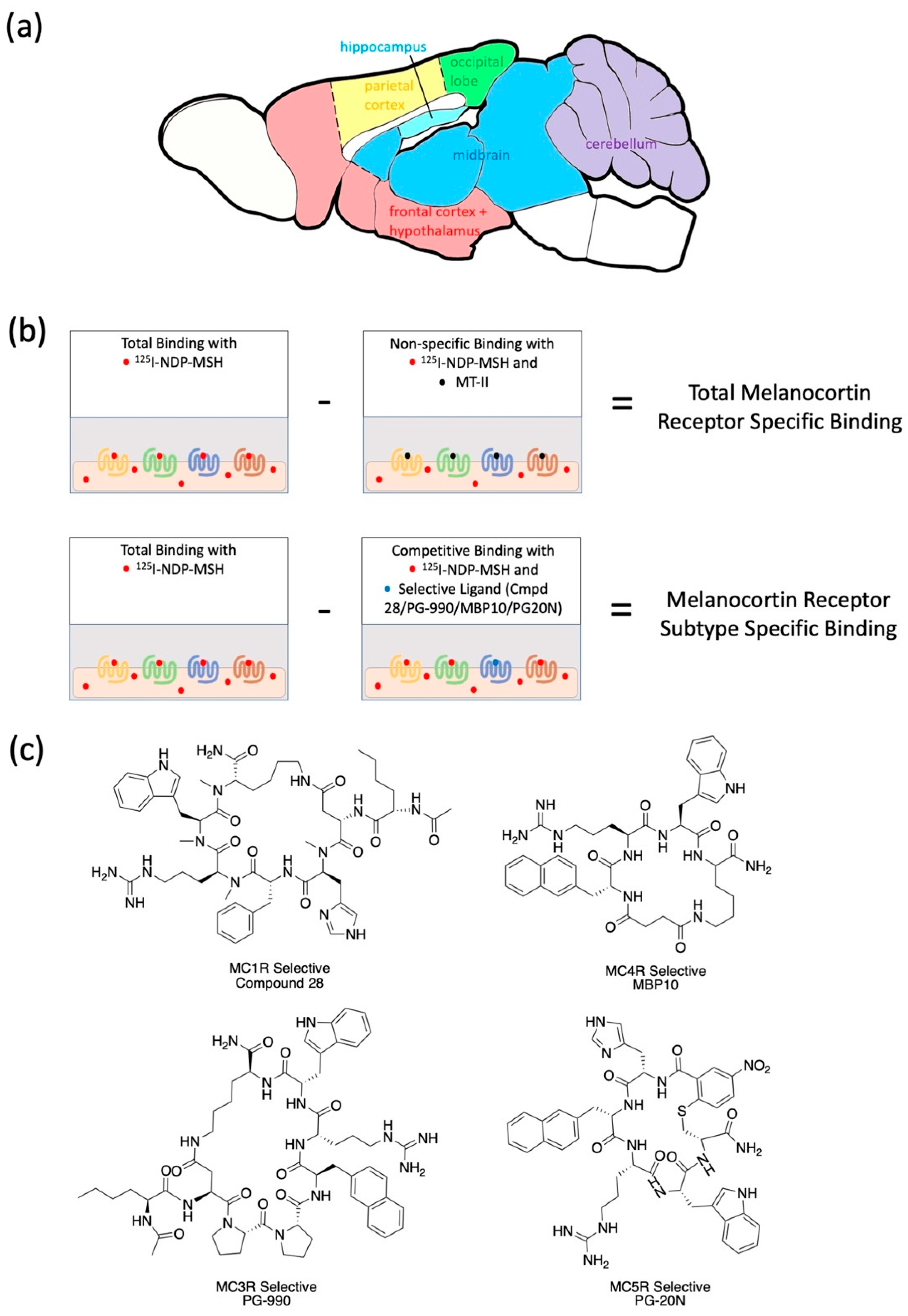
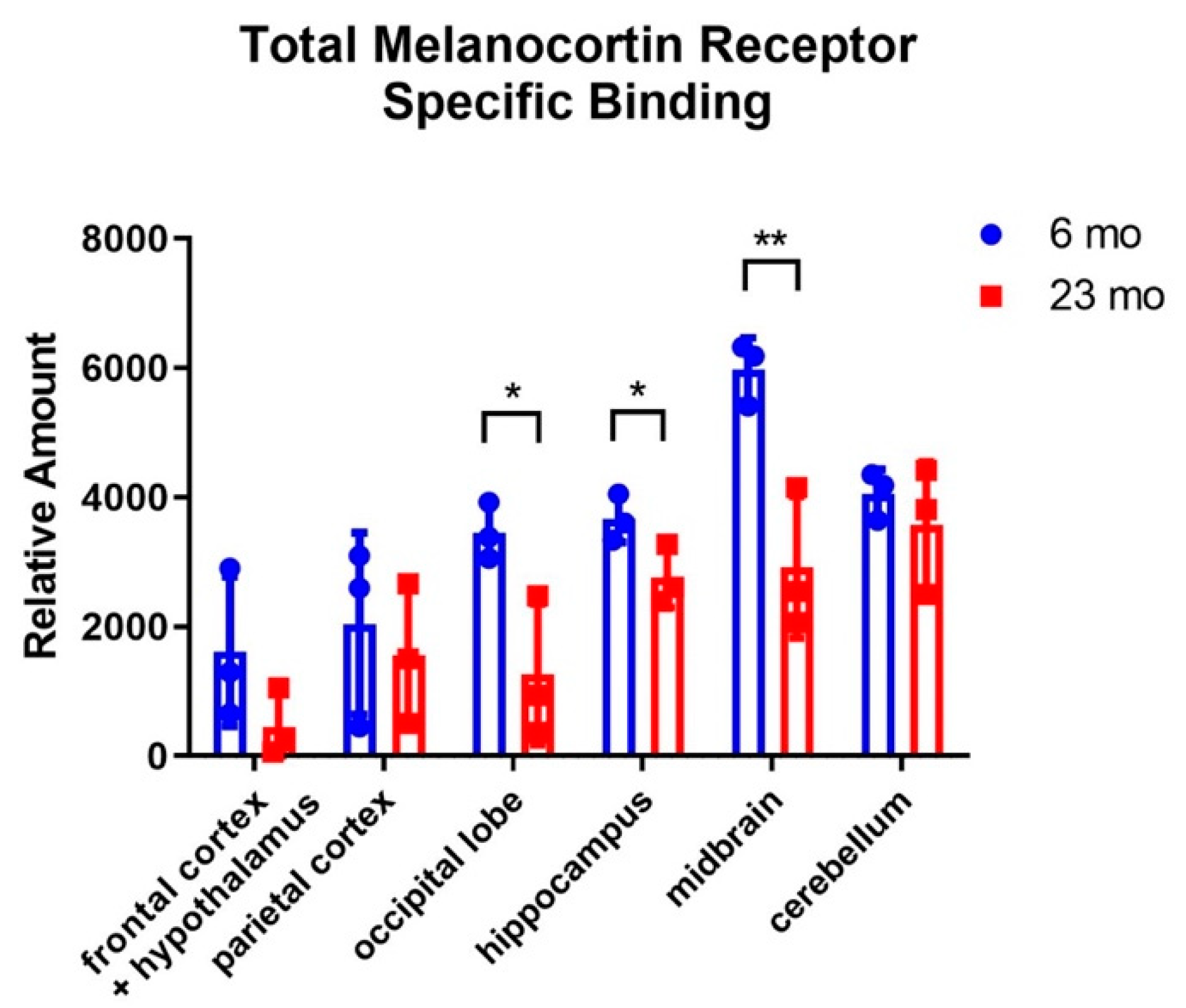
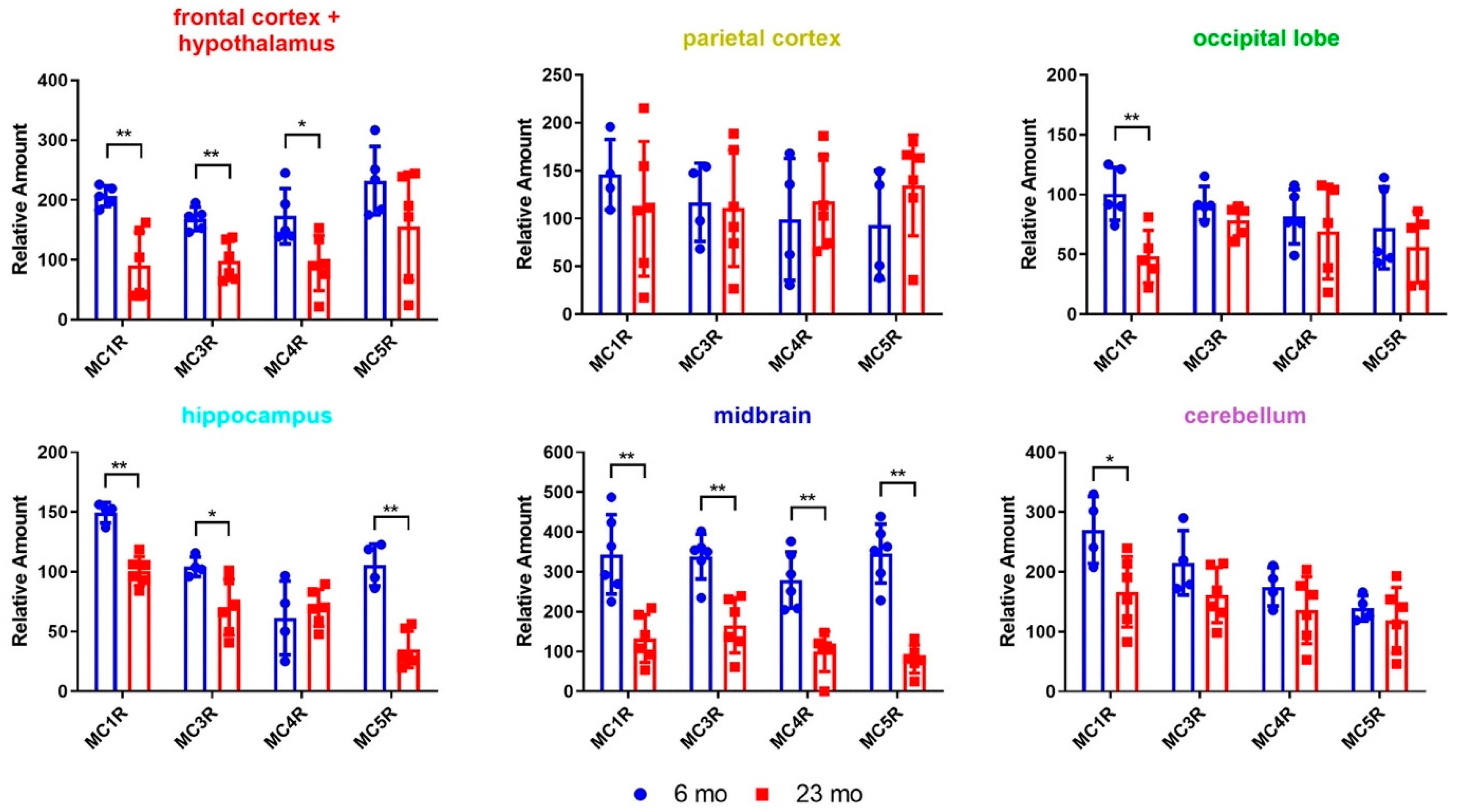
2.1. Radioligand Binding Assays to Determine Brain Region-Specific MCR Expression in Young (6 Mon) and Aged (23 Mon) Rats
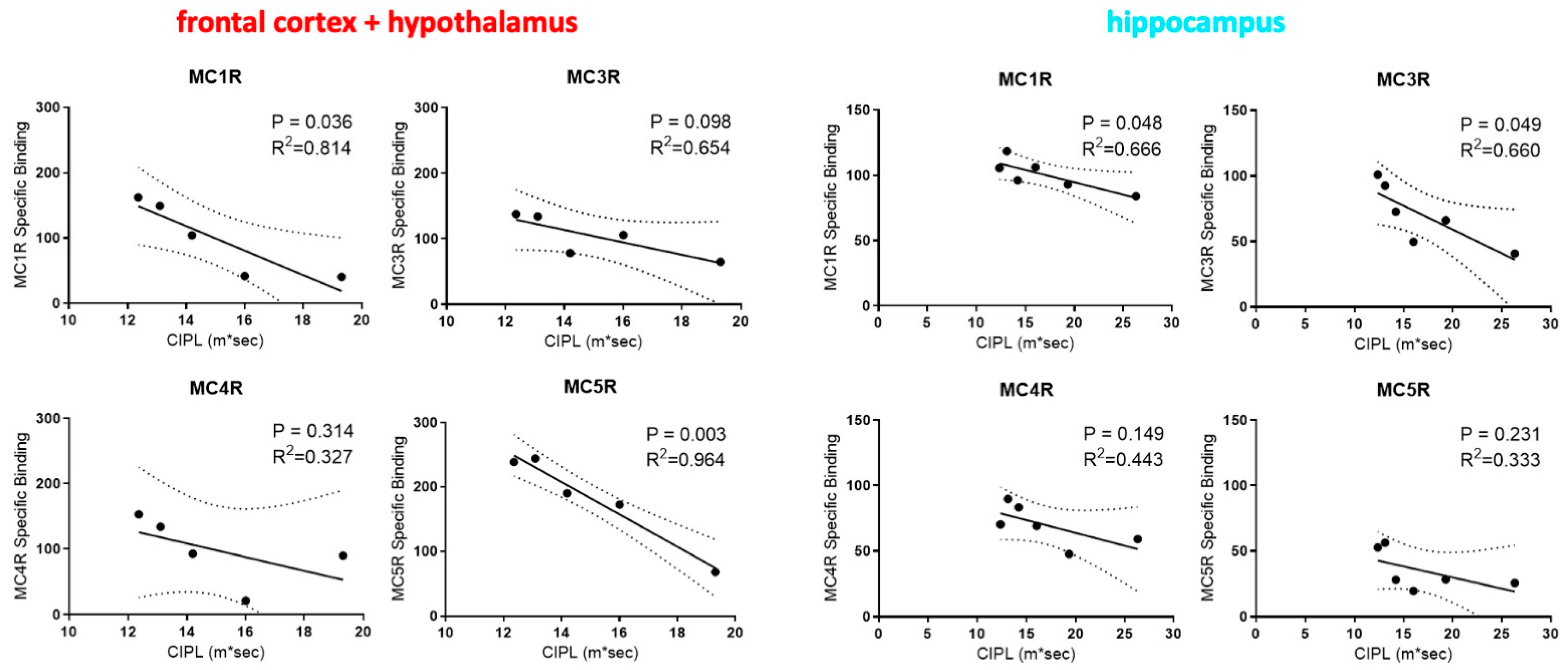
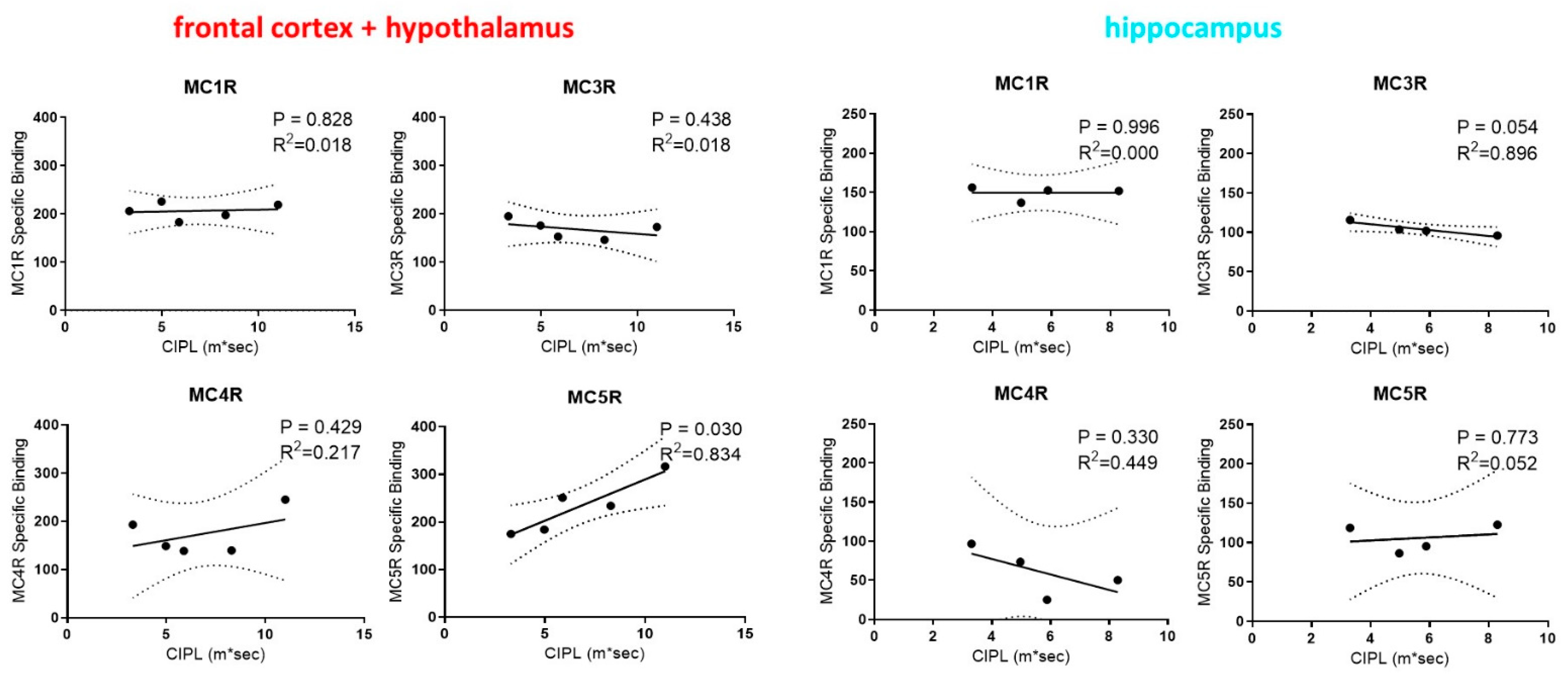
2.2. Correlations between Rats’ Performance in Spatial Learning and Memory and the Expression of MCRs in Their Brain
3. Discussion
4. Materials and Methods
4.1. Animals and Morris Water Maze Testing
4.2. Brain Extraction Procedures
4.3. Membrane Preparation and Protein Quantification
4.4. Membrane Binding Assays
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Holtzman, D.M.; Morris, J.C.; Goate, A.M. Alzheimer’s disease: The challenge of the second century. Sci. Transl. Med. 2011, 3, 77sr71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, P.J.; Watson, J.; Lund, J.; Davies, C.H.; Peters, G.; Dodds, C.M.; Swirski, B.; Lawrence, P.; Bentley, G.D.; O’Neill, B.V.; et al. The potent M1 receptor allosteric agonist GSK1034702 improves episodic memory in humans in the nicotine abstinence model of cognitive dysfunction. Int. J. Neuropsychopharmacol. 2013, 16, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Bradley, S.J.; Bourgognon, J.M.; Sanger, H.E.; Verity, N.; Mogg, A.J.; White, D.J.; Butcher, A.J.; Moreno, J.A.; Molloy, C.; Macedo-Hatch, T.; et al. M1 muscarinic allosteric modulators slow prion neurodegeneration and restore memory loss. J. Clin. Investig. 2017, 127, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.Y.; Watson, M.L.; Gallagher, M.; Nicolle, M.M. Muscarinic receptor-mediated GTP-Eu binding in the hippocampus and prefrontal cortex is correlated with spatial memory impairment in aged rats. Neurobiol. Aging 2007, 28, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.D.; Ferrari-DiLeo, G.; Mash, D.C.; Levey, A.I. Differential regulation of molecular subtypes of muscarinic receptors in Alzheimer’s disease. J. Neurochem. 1995, 64, 1888–1891. [Google Scholar] [CrossRef] [PubMed]
- Mulugeta, E.; Karlsson, E.; Islam, A.; Kalaria, R.; Mangat, H.; Winblad, B.; Adem, A. Loss of muscarinic M4 receptors in hippocampus of Alzheimer patients. Brain Res. 2003, 960, 259–262. [Google Scholar] [CrossRef]
- Piggott, M.; Owens, J.; O’Brien, J.; Paling, S.; Wyper, D.; Fenwick, J.; Johnson, M.; Perry, R.; Perry, E. Comparative distribution of binding of the muscarinic receptor ligands pirenzepine, AF-DX 384, (R,R)-I-QNB and (R,S)-I-QNB to human brain. J. Chem. Neuroanat. 2002, 24, 211–223. [Google Scholar] [CrossRef]
- Lebois, E.P.; Thorn, C.; Edgerton, J.R.; Popiolek, M.; Xi, S. Muscarinic receptor subtype distribution in the central nervous system and relevance to aging and Alzheimer’s disease. Neuropharmacology 2018, 136, 362–373. [Google Scholar] [CrossRef]
- Banuelos, C.; Beas, B.S.; McQuail, J.A.; Gilbert, R.J.; Frazier, C.J.; Setlow, B.; Bizon, J.L. Prefrontal cortical GABAergic dysfunction contributes to age-related working memory impairment. J. Neurosci. 2014, 34, 3457–3466. [Google Scholar] [CrossRef]
- McQuail, J.A.; Frazier, C.J.; Bizon, J.L. Molecular aspects of age-related cognitive decline: The role of GABA signaling. Trends Mol. Med. 2015, 21, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Simonyi, A.; Ngomba, R.T.; Storto, M.; Catania, M.V.; Miller, L.A.; Youngs, B.; DiGiorgi-Gerevini, V.; Nicoletti, F.; Sun, G.Y. Expression of groups I and II metabotropic glutamate receptors in the rat brain during aging. Brain Res. 2005, 1043, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Nabulsi, N.; Gaiser, E.; Gallezot, J.D.; Henry, S.; Planeta, B.; Lin, S.F.; Ropchan, J.; Williams, W.; Morris, E.; et al. Age-related change in 5-HT6 receptor availability in healthy male volunteers measured with (11)C-GSK215083 PET. J. Nucl. Med. 2018, 59, 1445–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froestl, W. Chemistry and pharmacology of GABAB receptor ligands. Adv. Pharmacol. 2010, 58, 19–62. [Google Scholar] [CrossRef] [PubMed]
- Quiedeville, A.; Boulouard, M.; Da Silva Costa-Aze, V.; Dauphin, F.; Bouet, V.; Freret, T. 5-HT6 receptor antagonists as treatment for age-related cognitive decline. Rev. Neurosci. 2014, 25, 417–427. [Google Scholar] [CrossRef]
- Jiang, S.; Li, Y.; Zhang, C.; Zhao, Y.; Bu, G.; Xu, H.; Zhang, Y.W. M1 muscarinic acetylcholine receptor in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Bruno, V.; Caraci, F.; Copani, A.; Matrisciano, F.; Nicoletti, F.; Battaglia, G. The impact of metabotropic glutamate receptors into active neurodegenerative processes: A “dark side” in the development of new symptomatic treatments for neurologic and psychiatric disorders. Neuropharmacology 2017, 115, 180–192. [Google Scholar] [CrossRef]
- Zhou, Y.; Cai, M. Novel approaches to the design of bioavailable melanotropins. Expert Opin. Drug Discov. 2017, 12, 1023–1030. [Google Scholar] [CrossRef]
- Roselli-Rehfuss, L.; Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Low, M.J.; Tatro, J.B.; Entwistle, M.L.; Simerly, R.B.; Cone, R.D. Identification of a receptor for gamma melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proc. Natl. Acad. Sci. USA 1993, 90, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Gantz, I.; Konda, Y.; Tashiro, T.; Shimoto, Y.; Miwa, H.; Munzert, G.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning of a novel melanocortin receptor. J. Biol. Chem. 1993, 268, 8246–8250. [Google Scholar] [CrossRef]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 2007, 445, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Hawrylycz, M.J.; Lein, E.S.; Guillozet-Bongaarts, A.L.; Shen, E.H.; Ng, L.; Miller, J.A.; van de Lagemaat, L.N.; Smith, K.A.; Ebbert, A.; Riley, Z.L.; et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 2012, 489, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.C.; King, M.G. Alpha-melanocyte-stimulating hormone and behavior. Neurosci. Biobehav. Rev. 1982, 6, 297–310. [Google Scholar] [CrossRef]
- McLay, R.N.; Pan, W.; Kastin, A.J. Effects of peptides on animal and human behavior: A review of studies published in the first twenty years of the journal Peptides. Peptides 2001, 22, 2181–2255. [Google Scholar] [CrossRef]
- Beckwith, B.E.; Sandman, C.A.; Hothersall, D.; Kastin, A.J. Influence of neonatal injections of alpha-MSH on learning, memory and attention in rats. Physiol. Behav. 1977, 18, 63–71. [Google Scholar] [CrossRef]
- Shen, Y.; Tian, M.; Zheng, Y.; Gong, F.; Fu, A.K.Y.; Ip, N.Y. Stimulation of the hippocampal POMC/MC4R circuit alleviates synaptic plasticity impairment in an Alzheimer’s disease model. Cell Rep. 2016, 17, 1819–1831. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, D.; Bitto, A.; Galantucci, M.; Zaffe, D.; Ottani, A.; Irrera, N.; Neri, L.; Cavallini, G.M.; Altavilla, D.; Botticelli, A.R.; et al. Melanocortins protect against progression of Alzheimer’s disease in triple-transgenic mice by targeting multiple pathophysiological pathways. Neurobiol. Aging 2014, 35, 537–547. [Google Scholar] [CrossRef]
- Giuliani, D.; Galantucci, M.; Neri, L.; Canalini, F.; Calevro, A.; Bitto, A.; Ottani, A.; Vandini, E.; Sena, P.; Sandrini, M.; et al. Melanocortins protect against brain damage and counteract cognitive decline in a transgenic mouse model of moderate Alzheimer’s disease. Eur. J. Pharmacol. 2014, 740, 144–150. [Google Scholar] [CrossRef]
- Giuliani, D.; Neri, L.; Canalini, F.; Calevro, A.; Ottani, A.; Vandini, E.; Sena, P.; Zaffe, D.; Guarini, S. NDP-α-MSH induces intense neurogenesis and cognitive recovery in Alzheimer transgenic mice through activation of melanocortin MC4 receptors. Mol. Cell Neurosci. 2015, 67, 13–21. [Google Scholar] [CrossRef]
- Al-Obeidi, F.; Castrucci, A.M.; Hadley, M.E.; Hruby, V.J. Potent and prolonged acting cyclic lactam analogues of alpha-melanotropin: Design based on molecular dynamics. J. Med. Chem. 1989, 32, 2555–2561. [Google Scholar] [CrossRef]
- Doedens, L.; Opperer, F.; Cai, M.; Beck, J.G.; Dedek, M.; Palmer, E.; Hruby, V.J.; Kessler, H. Multiple N-methylation of MT-II backbone amide bonds leads to melanocortin receptor subtype hMC1R selectivity: Pharmacological and conformational studies. J. Am. Chem. Soc. 2010, 132, 8115–8128. [Google Scholar] [CrossRef] [Green Version]
- Carotenuto, A.; Merlino, F.; Cai, M.; Brancaccio, D.; Yousif, A.M.; Novellino, E.; Hruby, V.J.; Grieco, P. Discovery of Novel Potent and Selective Agonists at the Melanocortin-3 Receptor. J. Med. Chem. 2015, 58, 9773–9778. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, M.A.; MacNeil, T.; Kalyani, R.N.; Tang, R.; Van der Ploeg, L.H.; Weinberg, D.H. Selective, high affinity peptide antagonists of alpha-melanotropin action at human melanocortin receptor 4: Their synthesis and biological evaluation in vitro. J. Med. Chem. 2001, 44, 3665–3672. [Google Scholar] [CrossRef]
- Grieco, P.; Cai, M.; Liu, L.; Mayorov, A.; Chandler, K.; Trivedi, D.; Lin, G.; Campiglia, P.; Novellino, E.; Hruby, V.J. Design and microwave-assisted synthesis of novel macrocyclic peptides active at melanocortin receptors: Discovery of potent and selective hMC5R receptor antagonists. J. Med. Chem. 2008, 51, 2701–2707. [Google Scholar] [CrossRef] [Green Version]
- D’Hooge, R.; De Deyn, P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res. Brain Res. Rev. 2001, 36, 60–90. [Google Scholar] [CrossRef]
- Cai, M.; Hruby, V.J. Design of cyclized selective melanotropins. Biopolymers 2016, 106, 876–883. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Beyer, A.; Aebersold, R. On the dependency of cellular protein levels on mRNA abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Fu, W.Y.; Cheng, E.Y.; Fu, A.K.; Ip, N.Y. Melanocortin-4 receptor regulates hippocampal synaptic plasticity through a protein kinase A-dependent mechanism. J. Neurosci. 2013, 33, 464–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, D.; Minutoli, L.; Ottani, A.; Spaccapelo, L.; Bitto, A.; Galantucci, M.; Altavilla, D.; Squadrito, F.; Guarini, S. Melanocortins as potential therapeutic agents in severe hypoxic conditions. Front. Neuroendocrinol. 2012, 33, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Mykicki, N.; Herrmann, A.M.; Schwab, N.; Deenen, R.; Sparwasser, T.; Limmer, A.; Wachsmuth, L.; Klotz, L.; Kohrer, K.; Faber, C.; et al. Melanocortin-1 receptor activation is neuroprotective in mouse models of neuroinflammatory disease. Sci. Transl. Med. 2016, 8, 362ra146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Doycheva, D.M.; Gamdzyk, M.; Yang, Y.; Lenahan, C.; Li, G.; Li, D.; Lian, L.; Tang, J.; Lu, J.; et al. Activation of MC1R with BMS-470539 attenuates neuroinflammation via cAMP/PKA/Nurr1 pathway after neonatal hypoxic-ischemic brain injury in rats. J. Neuroinflam. 2021, 18, 26. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, P.B. Role of inflammation in cognitive impairment: Results of observational epidemiological studies and clinical trials. Ann. N. Y. Acad. Sci. 2010, 1207, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Barnes, C.A. Age-related decrease in cholinergic synaptic transmission in three hippocampal subfields. Neurobiol. Aging 1996, 17, 439–451. [Google Scholar] [CrossRef]
- Gallagher, M.; Nicolle, M.M. Animal models of normal aging: Relationship between cognitive decline and markers in hippocampal circuitry. Behav. Brain Res. 1993, 57, 155–162. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Chawla, M.K.; Rios-Monterrosa, J.L.; Wang, L.; Zempare, M.A.; Hruby, V.J.; Barnes, C.A.; Cai, M. Aged Brains Express Less Melanocortin Receptors, Which Correlates with Age-Related Decline of Cognitive Functions. Molecules 2021, 26, 6266. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206266
Zhou Y, Chawla MK, Rios-Monterrosa JL, Wang L, Zempare MA, Hruby VJ, Barnes CA, Cai M. Aged Brains Express Less Melanocortin Receptors, Which Correlates with Age-Related Decline of Cognitive Functions. Molecules. 2021; 26(20):6266. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206266
Chicago/Turabian StyleZhou, Yang, Monica K. Chawla, Jose L. Rios-Monterrosa, Lingzhi Wang, Marc A. Zempare, Victor J. Hruby, Carol A. Barnes, and Minying Cai. 2021. "Aged Brains Express Less Melanocortin Receptors, Which Correlates with Age-Related Decline of Cognitive Functions" Molecules 26, no. 20: 6266. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206266