
Magnolol and Honokiol Inhibited the Function and Expression of BCRP with Mechanism Exploration

1
School of Pharmacy, College of Pharmacy, China Medical University, Taichung 406040, Taiwan
2
Department of Pharmacy, China Medical University Hospital, Taichung 404332, Taiwan
3
College of Medical and Health Science, Asia University, Taichung 41354, Taiwan
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to the study.
Molecules 2021, 26(23), 7390; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237390
Submission received: 1 November 2021
/
Revised: 25 November 2021
/
Accepted: 4 December 2021
/
Published: 6 December 2021
Abstract
:Breast cancer resistance protein (BCRP), one of the ATP-binding cassette (ABC) transporters, was associated with the multidrug resistance (MDR) of chemotherapy. Magnolol (MN) and honokiol (HK) are major bioactive polyphenols of Magnolia officinalis. This study investigated the effects of MN and HK on the function and expression of BCRP for the purpose of developing BCRP inhibitor to overcome MDR. Cell lines including MDCKII-BCRP and MDCKII-WT were used for evaluating the function and expression of BCRP. The results showed that MN (100–12.5 µM) and HK (100–12.5 µM) significantly decreased the function of BCRP by 80~12% and 67~14%, respectively. In addition, MN and HK were verified as substrates of BCRP. Furthermore, MN and HK reduced the protein expression of BCRP, and inhibited the phosphorylation of epidermal growth factor receptor (EGFR) and phosphatidylinositol 3-kinase (PI3K). In conclusion, both MN and HK decreased the function and expression of BCRP via EGFR/PI3K signaling pathway. Therefore, both compounds were promising candidates for reversing the MDR of chemotherapy.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
In recent decades, it is well recognized that overexpression of ATP-binding cassette (ABC) transporters, such as P-glycoprotein (P-gp), multidrug resistance-associated proteins (MRPs) and breast cancer resistance protein (BCRP), is one of the causative mechanisms of multidrug resistance (MDR) [1]. The inhibition on ABC transporter-mediated efflux from cancer cells was believed to be one of the feasible strategies to overcome MDR [2]. Many phytochemicals such as flavonoids, curcuminoids, taccalonolides, and terpenes have been reported to show inhibitions on ABC transporters. Therefore, there are growing interests to search for natural products as inhibitors of P-gp, MRPs and BCRP [3,4,5].
BCRP is the most recently reported ABC transporter and its tissue distribution highly resembles P-gp in tumors and various organs, and shared broad specificity of substrates [6]. The substrates of BCRP included a variety of therapeutic agents such as anticancer drugs (mitoxantrone, irinotecan, methotrexate, gefitinib), antiviral drugs (lamivudine, zidovudine), antihyperlipidemic drugs (rosuvastatin), antidiabetic drugs (glyburide) and anti-inflammatory drugs (sulfasalazine) as well as many acidic physiological substances such as estrone-3-sulfate, 17β-estradiol 17-(β-D-glucuronide), and uric acid [7]. Thus, BCRP has now been recognized by the USFDA to be one of the key ABC transporters involved in clinically relevant drug disposition, drug interactions [7] and might also play important roles in MDR [8].
A variety of natural compounds have been investigated in the efforts of developing MDR-reversing agents owing to their low toxicities [9]. Magnolol (MN) and honokiol (HK) (chemical structures shown in Figure 1), major bioactive polyphenols of Magnolia officinalis, have been shown to be safe and without troublesome adverse effects [10]. In pharmacological aspect, MN exhibited numerous beneficial pharmacological activities, such as anti-inflammatory [11,12], neuroprotection [13], antitumor [14,15], cardiovascular protection [16], anti-angiogenesis [17], antioxidation [18] and antibacterial activities [19]. Being a structural isomer of MN, HK also showed various beneficial biological effects, including neuroprotection [20], antitumor [21], anti-angiogenesis [22], antioxidation and antibacterial activities [23]. Concerning the associations with MDR, MN and HK were promising agents for reducing MDR via down-regulation of P-gp expression [24]. Moreover, MN and HK were inhibitors of nuclear factor-kappa B (NF-κB) and reversed the MDR of vinblastine, a substrate of P-gp [25].
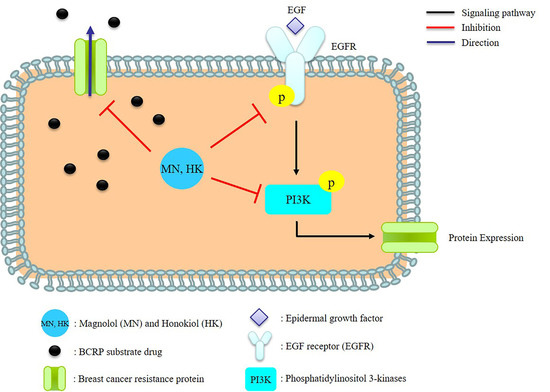
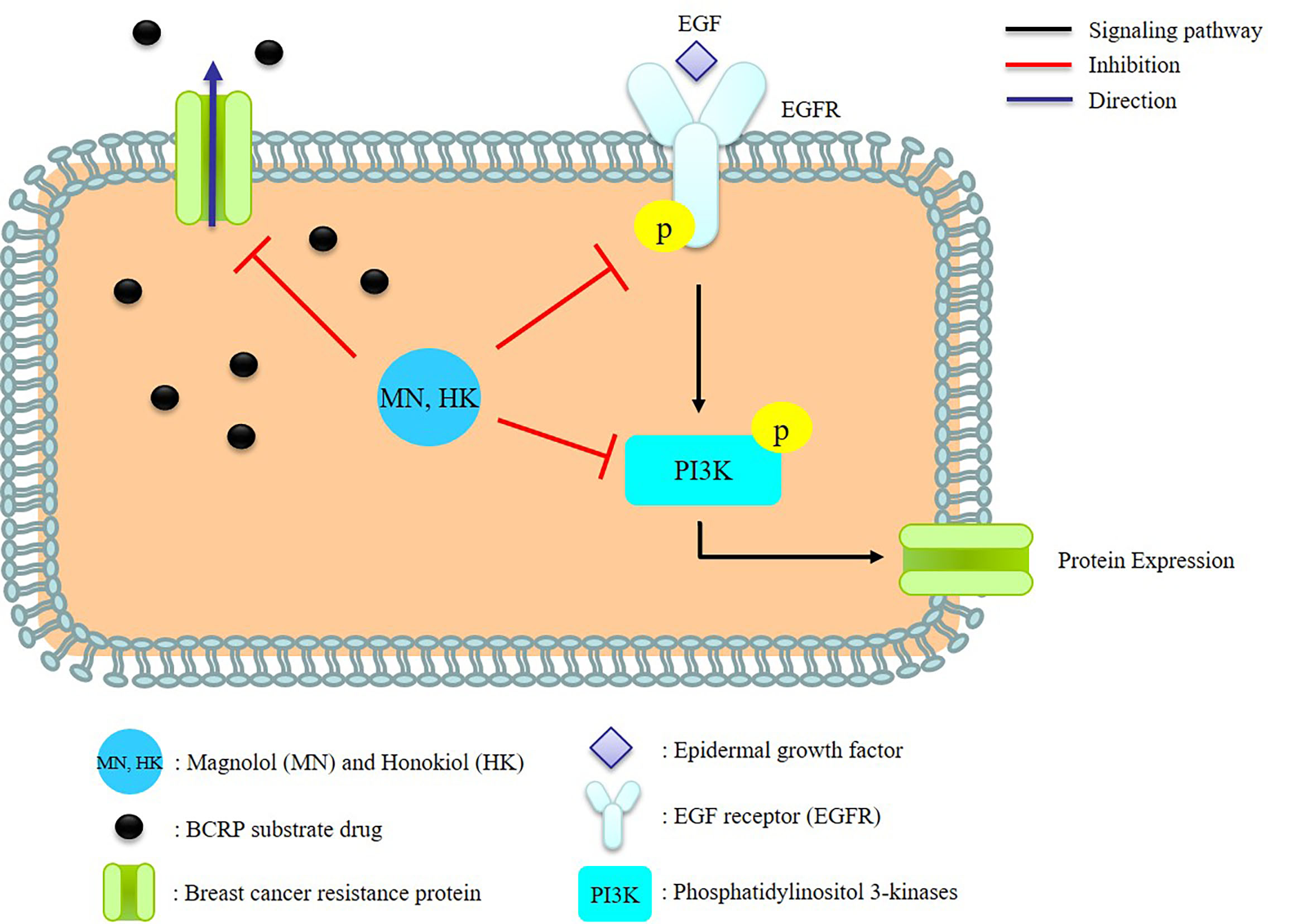
It has been recognized that epidermal growth factor receptor (EGFR) was overexpressed during the progression of a number of tumor types [26], and thus, suppressing the EGFR signaling pathways was an effective therapeutic target in treating cancers [27,28]. On the other hand, the induction of BCRP expression by EGF treatment has been demonstrated in several cell lines, including human BCRP-transfected Madin-Darby canine kidney cells (MDCKII-BCRP) [9,10]. Therefore, we speculated that BCRP expression was associated with EGFR/PI3K signaling pathway. Till now, the modulations of MN and HK on BCRP have not been reported yet. This study investigated the influences of MN and HK on the function and expression of BCRP. Furthermore, the modulation mechanisms were explored.
2. Results
2.1. Cell Viability Assay
MTT assay indicated that various concentrations of tested drugs and Ko143, a potent BCRP inhibitor used as positive control, at 0.25 μM all exerted no toxic influences on the viability of MDCKII-WT and MDCKII-BCRP cells (Supplementary Materials).
2.2. Effects of MN and HK on the Function of BCRP
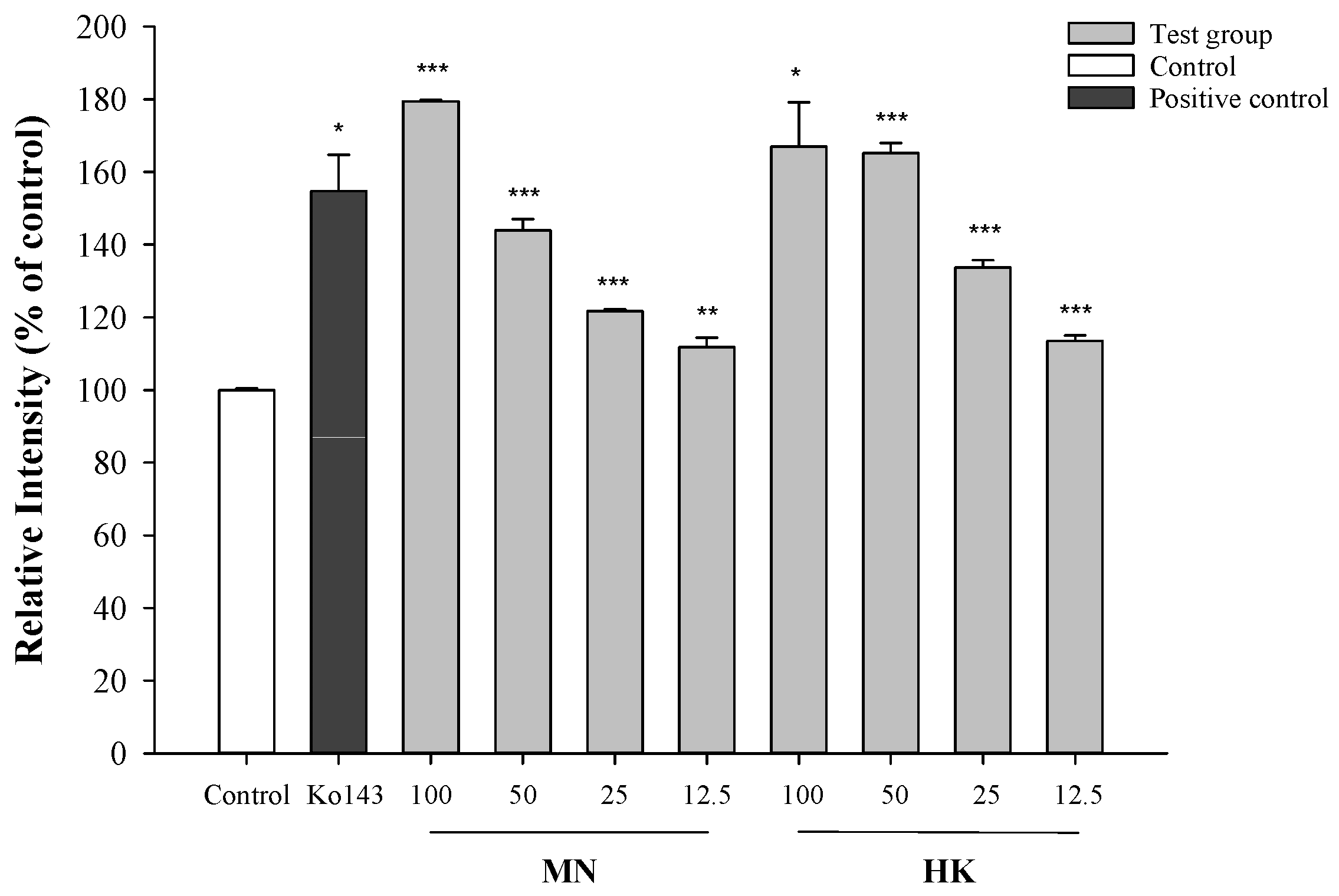
The effects of MN and HK on the intracellular accumulation of mitoxantrone (MXR), a typical BCRP substrate, in MDCKII-BCRP cells by using flow cytometry method are shown in Figure 2. The results showed that MN and HK at 100, 50, 25 and 12.5 μM significantly increased the intracellular accumulations of MXR by 80%, 44%, 22%, 12% (MN), and 67%, 65%, 34%, 14% (HK), respectively. Ko143 at 0.25 μM significantly increased the intracellular accumulations of MXR by 55%.
2.3. Intracellular Accumulations of MN and HK in MDCKII-WT and MDCKII-BCRP Cells
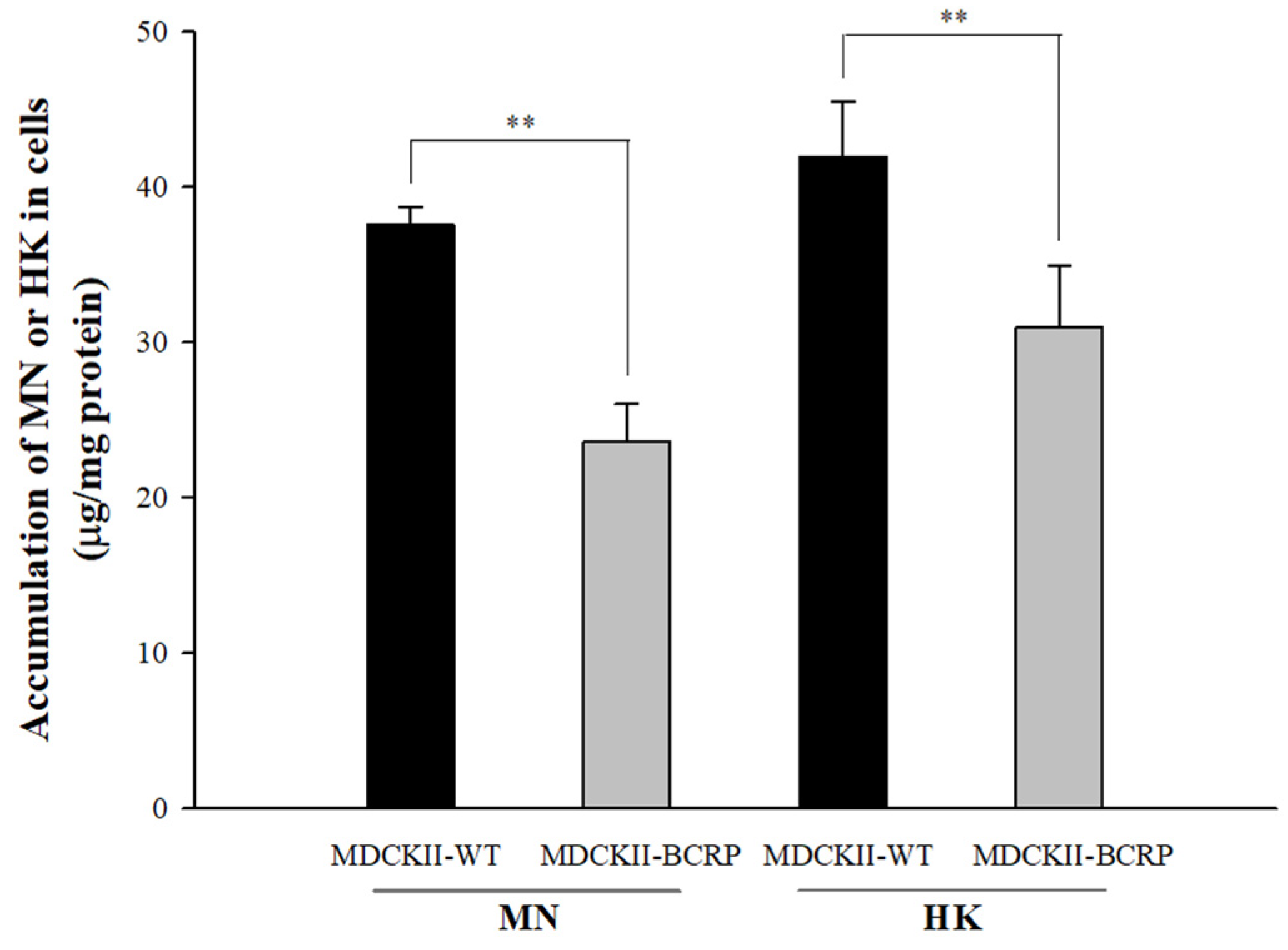
An HPLC-UV analytical method was established and validated in this study for the determination of MN and HK in cell lysates. The calibration ranges of MN (2.5–80 μg/mL) and HK (1.3–40 μg/mL) in cell lysates were with good linearity (the correlation coefficients of MN and HK were 0.9998 and 0.9996, respectively). The intracellular accumulations of MN and HK in MDCKII-WT and MDCKII-BCRP cells after incubations with MN and HK are shown in Figure 3. The intracellular concentrations of MN in MDCKII-BCRP cells after 30-min incubations were significantly lower than the corresponding concentrations in MDCKII-WT cells by 38%. Likewise, the intracellular concentration of HK in MDCKII-BCRP cells was significantly lower than the corresponding concentrations in MDCKII-WT cells by 27%. The results indicated that MN and HK are substrates of BCRP.
2.4. Effects of MN and HK on the Expressions of BCRP, EGFR, and PI3K
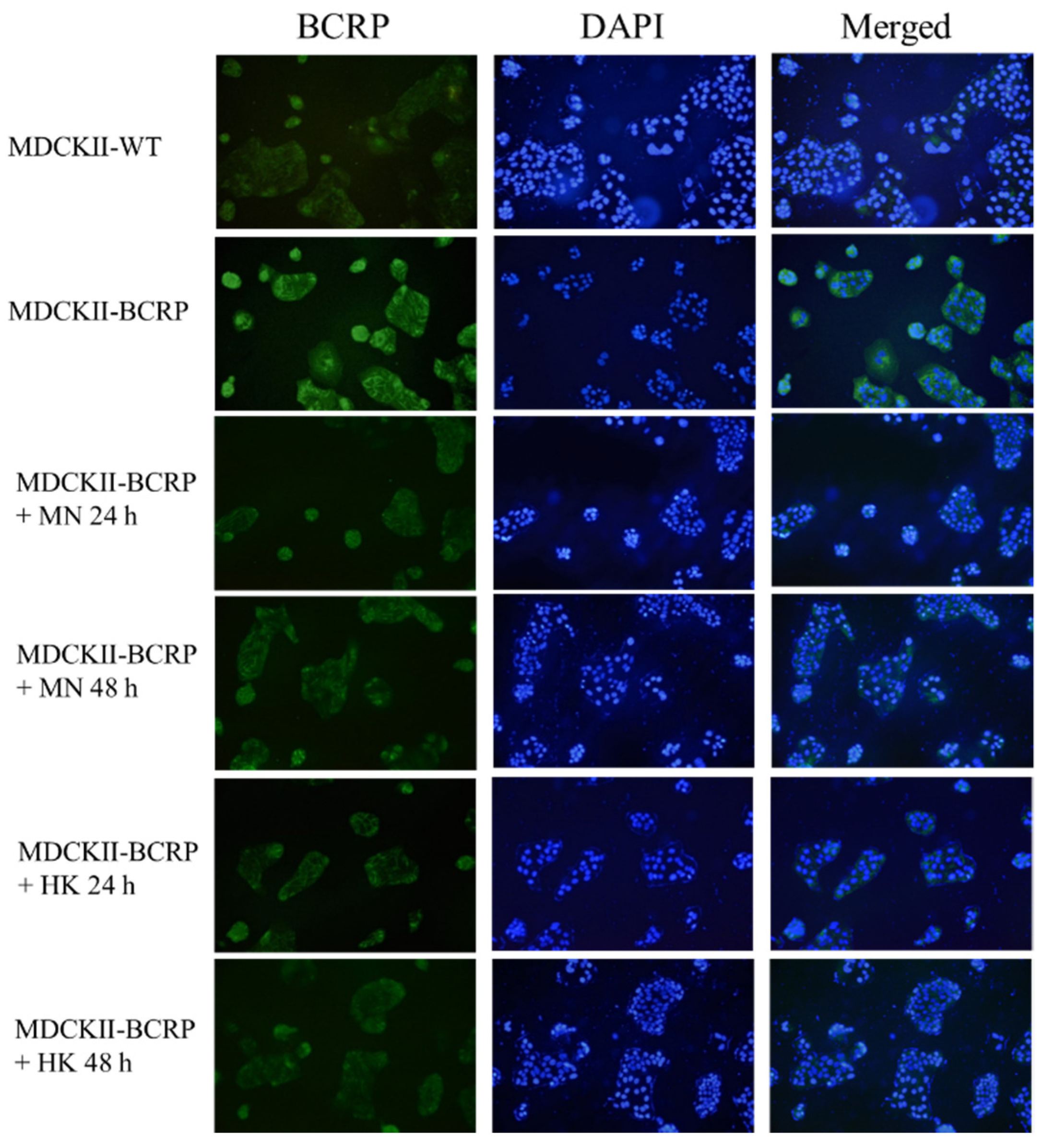
By using immunofluorescence assay, the expressions of BCRP in MDCKII-WT and MDCKII-BCRP cells after treating with MN and HK, individually, for 24 h and 48 h, are shown in Figure 4. The results indicated that MN and HK decreased the expression of BCRP in MDCKII-BCRP cells.
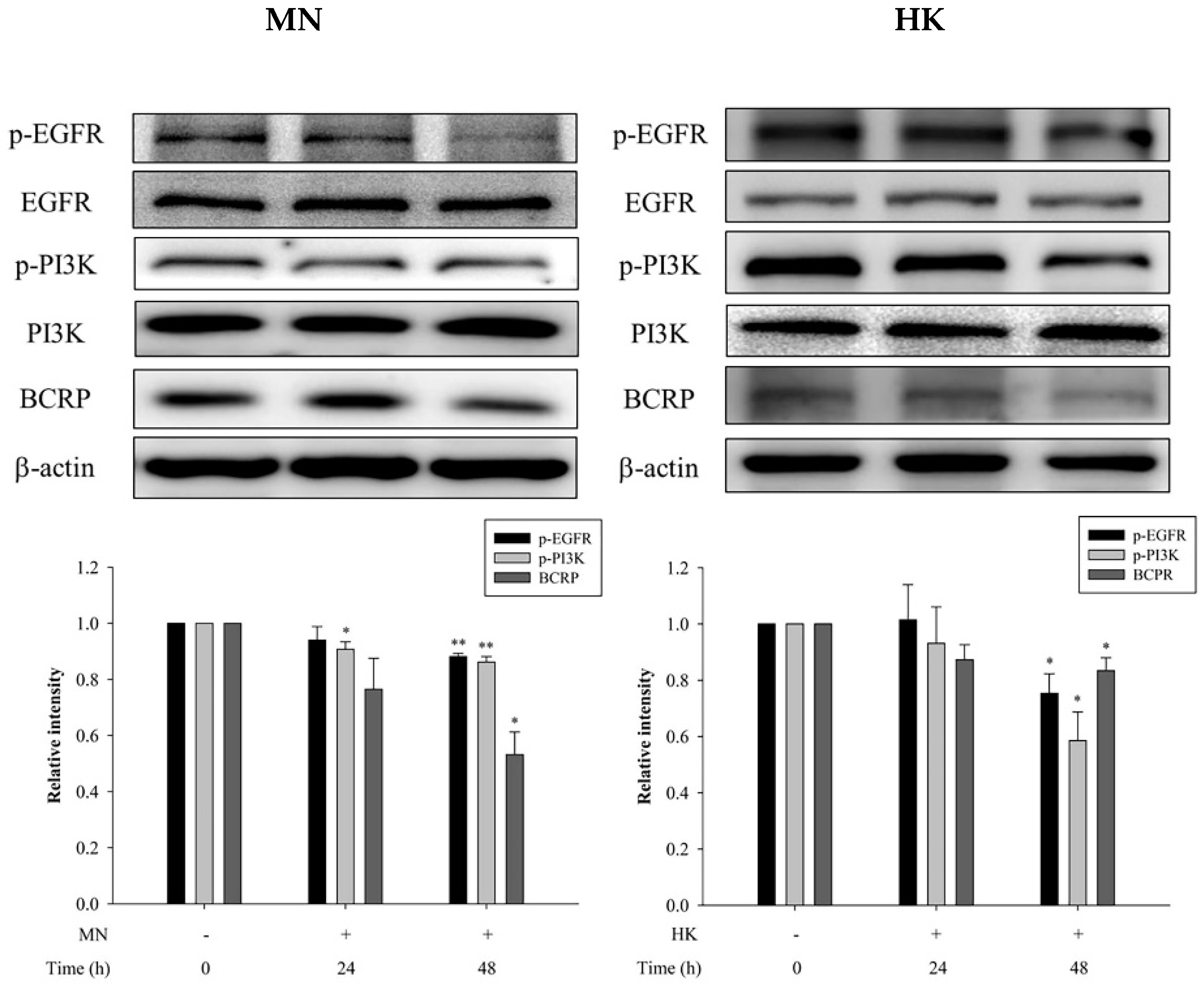
On other hand, through western blot analysis, the protein expressions of p-EGFR, p-PI3K and BCRP in MDCKII-BCRP cells after treating with MN and HK, individually, for 24 h and 48 h are shown in Figure 5. The results showed that after treatment with MN and HK for 48 h, the protein expressions of p-EGFR were significantly reduced by 12% and 25%, the protein expressions of p-PI3K were significantly decreased by 14% and 41%, and the protein expressions of BCRP were significantly reduced by 47% and 17%, respectively, indicaing that MN and HK decreased the expressions of p-EGFR, p-PI3K and BCRP. Moreover, Figure 5 also showed that the modulation effects were in a time-dependent manner.
3. Discussion
This study employed cell model to investigate the involvement of MN and HK in the BCRP-mediated transport and their modulation mechanism on BCRP. MXR was used as a substrate of BCRP. The transport assays showed that both MN and HK increased the intracellular accumulation of MXR, proving that MN and HK acted as inhibitors of BCRP. In order to verify whether MN and HK were substrates of BCRP, the intracellular accumulations of MN or HK in MDCKII-WT and MDCKII-BCRP cells were compared. The results showed that the intracellular accumulations of MN and HK in MDCKII-BCRP cells were significantly lower than those in MDCKII-WT cells, indicating that the efflux transports of MN and HK were mediated by BCRP. Taken together, MN and HK were substrates and inhibitors of BCRP.
In order to understand the modulation mechanism of MN and HK on BCRP, the influences on the EGFR/PI3K signaling pathway cells were investigated by using MDCKII-BCRP cells. The present results showed that MN and HK inhibited the protein expressions of p-EGFR and p-PI3K, thereby inhibiting BCRP expression, indicating that MN and HK down-regulated BCRP via the EGFR/PI3K signaling pathway. Recently, MN and HK have been reported as promising anticarcinogenic and anticancer agents through inhibiting PI3K/AKT/mammalian target of rapamycin (mTOR) signaling pathway [29]. In addition, MN triggered the apoptosis of human prostate cancer cells by suppressing the EGFR/PI3K/AKT signaling pathway [30]. Similarly, HK inhibited proliferation, invasion, and induced apoptosis through EGFR/PI3K/AKT signaling pathway [31]. Besides, MN and HK were shown to inhibit the protein expression of NF-κB, which was one of the major downstream targets of EGFR/PI3K and involved in a variety of processes, such as inflammatory, immune responses and MDR [32,33,34,35]. Therefore, our present results indicating that MN and HK inhibited EGFR/PI3K signaling pathway was in good agreements with these previous findings [29,30,31].
The PI3K/AKT/mTOR pathway was a key link modulating the MDR of cancers [34,36,37]. Moreover, if this signaling pathway was blocked, BCRP expression would be inhibited, and the MDR was probably reversed [27]. The present study revealing that MN and HK decreased the function and expression of BCRP through inhibiting the phosphorylation of EGFR and PI3K suggested that MN and HK were promising candidates for overcoming the MDR of chemotherapy using BCRP substrate drugs. In brief, this was the first study to demonstrate that MN and HK were substrates/inhibitors of BCRP and worthy of further investigations as a single anticancer agent or in combined therapeutics with other anticancer drugs such as mitoxantrone, topotecan and gefitinib etc. [38,39].
4. Materials and Methods
4.1. Chemicals and Reagents
MN (purity > 98%) and HK (purity > 98%) were purchased from ChemFaces (Wuhan, PRC). Ko143 (purity 96%) was obtained from Enzo Life Sciences, Inc. (Farmingdale, NY, USA). MXR (purity 99%) was obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Dimethyl sulfoxide (DMSO), formic acid, sodium dodecyl sulfate (SDS), 3-(4′,5′-dimethylthiazol-2′-yl)-2,5-diphenyltetrazolium bromide (MTT), butylparaben and triton X-100 were supplied by Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS) was obtained from Biological Industries Inc. (Kibbutz, Beit Haemek, Israel). Penicillin-Streptomycin-Glutamine, Dulbecco’s modified Eagle medium (DMEM), 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, Hank’s buffered salt solution (HBSS)and trypsin/EDTA were purchased from Invitrogen (Grand Island, NY, USA). Antibodies against ABCG2 and GAPDH were purchased from GeneTex (San Antonio, TX, USA). Polyvinylidene fluoride transfer membranes (Immobilon P) and chemiluminescence (ECL) were obtained from Millipore Corp. (Bedford, MA, USA). 4′,6-diamidino-2-phenylindole (DAPI) staining solution and goat anti-rabbit IgG H&L (DyLight™ 488) antibody were purchased from Abcam (Cambridge, UK). Acetonitrile (ACN) and methanol with LC grade were obtained from Mallinckrodt Baker (Phillipsburg, NJ, USA). Milli-Q plus water (Bedford, MA, USA) was used throughout this study.
4.2. Cell Lines and Culture Conditions
MDCKII-WT and MDCKII-BCRP cells were kindly provided by Prof. Dr. Piet Borst (Netherlands Cancer Institute, Amsterdam, Netherlands). Cells were grown in Dulbecco’s modified Eagle medium supplemented with 10% fetal bovine serum, 100 units/mL of penicillin, 100 μg/mL of streptomycin and 292 μg/mL of glutamine at 37 °C in a humidified incubator containing 5% CO2. The medium was changed every other day and cells were subcultured when 80% to 90% confluency was reached.
4.3. Cell Viability Assay
The effects of MN, HK and Ko143 on the viability of MDCKII-WT and MDCKII-BCRP cells were evaluated by MTT assay, modified from a previous study [40]. After seeding the cells into a 96-well plate for overnight incubation, these tested drugs were added and incubated for suitable time depending on each experimental design, then 100 μL of MTT (5 mg/mL) was added and incubated for additional 3 h. DMSO solution was added to lyze the cell and then the cell viability was measured at 595 nm by a microplate reader (BioTek instruments lnc., Winooski, VT, USA).
4.4. Effects of MN and HK on the Function of BCRP
MDCKII-BCRP cells were used to evaluate the effects of MN and HK on the efflux transport of MXR, a fluorescent typical substrate of BCRP [41]. Briefly, cell suspension (5 × 105 in each reaction tube) was pre-incubated with a series concentration of MN and HK, and Ko143 (0.25 μM, an inhibitor of BCRP) at 37 °C for 15 min. MXR (5 μM) was then added and co-incubated for another 30 min. After incubation, cells were washed and re-suspended in ice-cold phosphate buffered saline. The intracellular fluorescence of MXR was measured by a FACScan flow cytometer. The transport studies were performed in triplicates.
4.5. The Intracellular Accumulations of MN and HK in MDCKII-WT and MDCKII-BCRP Cells
In order to verify whether MN and HK are substrates of BCRP, MDCKII-WT and MDCKII-BCRP cells were used for comparison. The cells were seeded onto 12-well plates at a density of 3 × 105 cells per well. After 3-day culturing, the medium was removed and washed with ice-cold phosphate-buffered saline. Four hundred microliters of MN or HK (100 μM in pH 7.4 HBSS) was added into each well and incubated at 37 °C for 30 min. After washing, cell lysates were obtained after trypsinization with 300 μL of 0.5% Trypsin-EDTA and lyzed by liquid nitrogen. The cell lysate (100 μL) was added to 100 μL of pH 5.0 buffer, 50 μL of ascorbic acid, 50 μL of 0.1 N HCl and partitioned with 300 μL of ethyl acetate containing 40 μg/mL of butylparaben as internal standard. After centrifugation, the ethyl acetate layer was dried by nitrogen gas and reconstituted with 50 μL of acetonitrile, and 20 μL was subject to HPLC-UV analysis. For calibrator preparation, cell lysate (100 μL) was spiked with various concentrations of MN (2.5–80.0 μg/mL) or HK (1.3–40.0 μg/mL) and then mixed with 100 μL of pH 5.0 buffer, 50 μL of ascorbic acid, 50 μL of 0.1 N HCl, then partitioned with 300 μL of ethyl acetate containing 40 μg/mL of butylparaben. The later procedure was identical to that described above for cell lysates. The intracellular concentrations of MN and HK were calculated after correction with protein contents.
4.6. Effects of MN and HK on the Expression of BCRP
Cells were seeded into 12-wells plates at a density of 1 × 104/wells and incubated with MN (12.5 μM) and HK (12.5 μM), individually, at 37 °C for 24 h and 48 h [42]. After the supernatant was removed, methanol was added and stood for 10 min. After washing, 0.1% Triton X-100 was incubated for 10 min, washed 3 times with PBS, and incubated with 1% bovine serum albumin (BSA) for 1 h. Cells were then washed 3 times in PBS, and then incubated with primary antibody overnight at 4 °C. After washing, cells were incubated with the DyLight™ 488-conjugated goat anti-rabbit IgG antibody (Jackson ImmunoResearch, West Grove, PA, USA) at room temperature for 2 h. After washing, cells were incubated with the 4′,6-diamidino-2-phenylindole (DAPI) for 10 min in the dark. The imaging of cells was acquired by using confocal laser scanning microscopy (Nikon, TE2000-U, Tokyo, Japan).
4.7. Effects of MN and HK on BCRP Expression and EGFR/PI3K Signaling Pathway
After treatment with MN (12.5 μM) and HK (12.5 μM) for 24 h and 48 h, individually, the cells were lyzed with radioimmune precipitation (RIPA) buffer (Merck, Darmstadt, Germany) and collected. The samples were separated by 10% SDS-polyacrylamide gel electrophoresis and then transfered onto polyvinylidenedifluoride membranes (Immobilon, Millipore, Bedford, MA, USA). The membranes were blocked at room temperature for 1 min in the blocking buffer (Goal Bio, Taipei, TW), and then washed 3 times with 0.1% TBST (Tris-buffered saline with 0.1% Tween® 20 detergent). After washing, the blots were incubated with p-EGFR, EGFR, p-PI3K, PI3K, BCRP and β-actin primary antibodies, individually, at 4 °C overnight. Then the blots were washed 3 times with 0.1% TBST, and reacted with secondary antibodies at room temperature for 1 h. The bands were detected by using the ECL kit (Advansta Inc., San Jose, CA, USA).
4.8. Data Analysis
The statistical software SPSS was used for analyzing the differences among treatments by using unpaired Student’s t-test, taking p < 0.05 as significant.
5. Conclusions
MN and HK decreased the function and expression of BCRP through inhibition on the EGFR/PI3K signaling pathway. This is the first study revealing that MN and HK were substrates/inhibitors of BCRP and they are potential candidates to overcome MDR of BCRP substrate drugs.
Supplementary Materials
The following are available online. Figure S1: Effects of MN (μM) and HK (μM) on the cell viability of MDCKII-WT and MDCKII-BCRP at 1 h. (Mean ± S.D.) Control: 0.01% DMSO in reaction buffer, Figure S2: Effects of MN (μM) and HK (μM) on the cell viability of MDCKII-BCRP at 24 and 48 h. (Mean ± S.D.) Control: 0.01% DMSO in reaction buffer.
Author Contributions
Conceptualization, Y.-C.H. and S.-P.L.; methodology, Y.-C.H. and S.-P.L.; software, C.-P.Y., P.-Y.L. and S.-Y.C.; validation, S.-Y.C. and P.-Y.L.; formal analysis, S.-Y.C. and P.-Y.L.; investigation, C.-P.Y., P.-Y.L. and S.-Y.C.; resources, Y.-C.H. and S.-P.L.; data curation, S.-Y.C. and P.-Y.L.; writing—original draft preparation, C.-P.Y. and P.-Y.L.; writing—review and editing, Y.-C.H., S.-P.L. and C.-P.Y.; supervision, Y.-C.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Technology, Taiwan (MOST 108-2320-B-039-041-MY3), China Medical University, Taiwan (CMU109-MF-20) and Medical University Hospital, Taiwan (DMR-108-146).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are not available.
Conflicts of Interest
The authors have declared no conflict of interest.
Sample Availability
Samples of the compounds in this study are available from the authors.
Abbreviations
ABC, ATP-binding cassette; ACN, acetonitrile; AKT, protein kinase B; BCRP, breast cancer resistance protein; DAPI, 4′,6-diamidino-2-phenylindole; DMEM, Dulbecco’s modified Eagle medium; DMSO, dimethyl sulfoxide; ECL, chemiluminescence; EGF, epidermal growth factor; EGFR, EGF receptor; ERK, extracellular signal regulated kinase; FBS, fetal bovine serum; HBSS, Hank’s buffered salt solution; HK, honokiol; MAPK, mitogen-activated protein kinase; MDCKII, Madin-Darby canine kidney type II cells; MDR, multidrug resistance; MEK, mitogen-activated protein kinase; MN, magnolol; MRPs, multidrug resistance-associated proteins; mTOR, mammalian target of rapamycin; MTT, 3-(4′,5′-dimethylthiazol-2′-yl)-2,5-diphenyltetrazolium bromide; MXR, mitoxantrone; NF-κB, nuclear factor-kappa B; P-gp, P-glycoprotein; PI3K, phosphatidylinositol 3-kinase; SDS, sodium dodecyl sulfate; TKIs, tyrosine kinase inhibitors.
References
- Gillet, J.P.; Gottesman, M.M. Mechanisms of multidrug resistance in cancer. Methods Mol. Biol. 2010, 596, 47–76. [Google Scholar] [PubMed]
- Wu, C.P.; Calcagno, A.M.; Ambudkar, S.V. Reversal of ABC drug transporter-mediated multidrug resistance in cancer cells: Evaluation of current strategies. Curr. Mol. Pharmacol. 2008, 1, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Tinoush, B.; Shirdel, I.; Wink, M. Phytochemicals: Potential Lead Molecules for MDR Reversal. Front. Pharmacol. 2020, 11, 832. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.M.F.; Cardoso, D.S.P.; Ferreira, M.-J.U. Overcoming Multidrug Resistance: Flavonoid and Terpenoid Nitrogen-Containing Derivatives as ABC Transporter Modulators. Molecules 2020, 25, 3364. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Silvia, D.; Laura, B.; Maria, N.R.; Elisabetta, T. Recent advances in the search of BCRP- and dual P-gp/BCRP-based multidrug resistance modulators. Cancer Drug Resist. 2019, 2, 710–743. [Google Scholar]
- Mao, Q.; Unadkat, J.D. Role of the breast cancer resistance protein (BCRP/ABCG2) in drug transport—An update. AAPS J. 2015, 17, 65–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, T.; Ross, D.D. Breast cancer resistance protein (BCRP/ABCG2): Its role in multidrug resistance and regulation of its gene expression. Chin. J. Cancer 2012, 31, 73–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaoglu, M.; Güllüce, M.; Karradayi, M. Plant-Derived Natural Products as Multidrug Resistance Modulators in Cancer Therapy. Anatol. J. Biol. 2020, 1, 1–51. [Google Scholar]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and Toxicology of Magnolol and Honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, J.; Xu, W.; Sun, Y.; You, J.; Lu, H.; Song, Y.; Wei, J.; Li, L. Magnolol inhibits Streptococcus suis-induced inflammation and ROS formation via TLR2/MAPK/NF-kappaB signaling in RAW264.7 cells. Pol. J. Vet. Sci. 2018, 21, 111–118. [Google Scholar] [PubMed]
- Chen, H.; Fu, W.; Chen, H.; You, S.; Liu, X.; Yang, Y.; Wei, Y.; Huang, J.; Rui, W. Magnolol attenuates the inflammation and enhances phagocytosis through the activation of MAPK, NF-kappaB signal pathways in vitro and in vivo. Mol. Immunol. 2019, 105, 96–106. [Google Scholar] [CrossRef]
- Xie, Z.; Zhao, J.; Wang, H.; Jiang, Y.; Yang, Q.; Fu, Y.; Zeng, H.; Holscher, C.; Xu, J.; Zhang, Z. Magnolol alleviates Alzheimer’s disease-like pathology in transgenic C. elegans by promoting microglia phagocytosis and the degradation of beta-amyloid through activation of PPAR-gamma. Biomed. Pharmacother. 2020, 124, 109886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.H.; Ren, H.Y.; Shen, J.X.; Zhang, X.Y.; Ye, H.M.; Shen, D.Y. Magnolol suppresses the proliferation and invasion of cholangiocarcinoma cells via inhibiting the NF-kappaB signaling pathway. Biomed. Pharmacother. 2017, 94, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Ma, H.; Zhang, T.; Liu, H.; Yu, L.; Li, G.; Li, H.; Hu, M. Magnolol Inhibits the Growth of Non-Small Cell Lung Cancer via Inhibiting Microtubule Polymerization. Cell Physiol. Biochem. 2017, 42, 1789–1801. [Google Scholar] [CrossRef]
- Chang, H.; Chang, C.Y.; Lee, H.J.; Chou, C.Y.; Chou, T.C. Magnolol ameliorates pneumonectomy and monocrotaline-induced pulmonary arterial hypertension in rats through inhibition of angiotensin II and endothelin-1 expression. Phytomedicine 2018, 51, 205–213. [Google Scholar] [CrossRef]
- Chen, M.C.; Lee, C.F.; Huang, W.H.; Chou, T.C. Magnolol suppresses hypoxia-induced angiogenesis via inhibition of HIF-1alpha/VEGF signaling pathway in human bladder cancer cells. Biochem. Pharmacol. 2013, 85, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Baschieri, A.; Pulvirenti, L.; Muccilli, V.; Amorati, R.; Tringali, C. Chain-breaking antioxidant activity of hydroxylated and methoxylated magnolol derivatives: The role of H-bonds. Org. Biomol. Chem. 2017, 15, 6177–6184. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Ding, H.; Liu, Y.; Yang, Q.; Xu, N.; Yang, Y.; Ai, X. Magnolol protects channel catfish from Aeromonas hydrophila infection via inhibiting the expression of aerolysin. Vet. Microbiol. 2017, 211, 119–123. [Google Scholar] [CrossRef]
- Wang, H.; Liao, Z.; Sun, X.; Shi, Q.; Huo, G.; Xie, Y.; Tang, X.; Zhi, X.; Tang, Z. Intravenous administration of Honokiol provides neuroprotection and improves functional recovery after traumatic brain injury through cell cycle inhibition. Neuropharmacology 2014, 86, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.P.; Lee, W.L.; Tang, Y.Q.; Yap, W.H. Honokiol: A Review of Its Anticancer Potential and Mechanisms. Cancers 2020, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, L.E.; Arbiser, J.L. Honokiol, a multifunctional antiangiogenic and antitumor agent. Antioxid. Redox Signal. 2009, 11, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.L.; Man, K.M.; Huang, P.H.; Chen, W.C.; Chen, D.C.; Cheng, Y.W.; Liu, P.L.; Chou, M.C.; Chen, Y.H. Honokiol and magnolol as multifunctional antioxidative molecules for dermatologic disorders. Molecules 2010, 15, 6452–6465. [Google Scholar] [CrossRef] [PubMed]
- Han, H.K.; Anh, L.T.V. Modulation of P-Glycoprotein Expression by Honokiol, Magnolol and 4-O-Methylhonokiol, the Bioactive Components of Magnolia officinalis. Anticancer Res. 2012, 32, 4445–4452. [Google Scholar]
- Nabekura, T.; Hiroi, T.; Kawasaki, T.; Uwai, Y. Effects of natural nuclear factor-kappa B inhibitors on anticancer drug efflux transporter human P-glycoprotein. Biomed. Pharmacother. 2015, 70, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Levitzki, A.; Klein, S. My journey from tyrosine phosphorylation inhibitors to targeted immune therapy as strategies to combat cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 11579–11586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pick, A.; Wiese, M. Tyrosine kinase inhibitors influence ABCG2 expression in EGFR-positive MDCK BCRP cells via the PI3K/Akt signaling pathway. ChemMedChem 2012, 7, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Chen, Y.-J.; Hung, M.-C. Implication of nuclear EGFR in the development of resistance to anticancer therapies. BioMedicine 2011, 1, 2–10. [Google Scholar] [CrossRef]
- Mottaghi, S.; Abbaszadeh, H. Natural Lignans Honokiol and Magnolol as Potential Anticarcinogenic and Anticancer Agents. A Comprehensive Mechanistic Review. Nutr. Cancer 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Szczepanski, M.J.; Lee, Y.J. Magnolol induces apoptosis via inhibiting the EGFR/PI3K/Akt signaling pathway in human prostate cancer cells. J. Cell. Biochem. 2009, 106, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, R.Z.; Jiang, Z.B.; Wei, C.L.; Luo, L.X.; Yao, X.J.; Li, G.P.; Leung, E.L. Honokiol Inhibits Proliferation, Invasion and Induces Apoptosis Through Targeting Lyn Kinase in Human Lung Adenocarcinoma Cells. Front. Pharmacol. 2018, 9, 558. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, L.; Liu, R.; Zhang, D.; Lan, X.; Huang, C.; Xin, W.; Wang, C.; Zhang, D.; Du, G. A novel naturally occurring salicylic acid analogue acts as an anti-inflammatory agent by inhibiting nuclear factor-kappaB activity in RAW264.7 macrophages. Mol. Pharm. 2012, 9, 671–677. [Google Scholar] [CrossRef]
- Labbozzetta, M.; Notarbartolo, M.; Poma, P. Can NF-kappaB Be Considered a Valid Drug Target in Neoplastic Diseases? Our Point of View. Int. J. Mol. Sci. 2020, 21, 3070. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, Y.; Liu, G.; Li, C.; Song, Y.; Cao, Z.; Li, W.; Hu, J.; Lu, C.; Liu, Y. PI3K/AKT pathway as a key link modulates the multidrug resistance of cancers. Cell Death Dis. 2020, 11, 797. [Google Scholar] [CrossRef]
- Nabekura, T.; Kawasaki, T.; Furuta, M.; Kaneko, T.; Uwai, Y. Effects of Natural Polyphenols on the Expression of Drug Efflux Transporter P-Glycoprotein in Human Intestinal Cells. ACS Omega 2018, 3, 1621–1626. [Google Scholar] [CrossRef]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/mTOR Pathway Causes Drug Resistance in Breast Cancer. Front. Pharmacol. 2021, 12, 628690. [Google Scholar] [CrossRef] [PubMed]
- Akinleye, A.; Avvaru, P.; Furqan, M.; Song, Y.; Liu, D. Phosphatidylinositol 3-kinase (PI3K) inhibitors as cancer therapeutics. J. Hematol. Oncol. 2013, 6, 88. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, I.; Nishikawa, S.; Yamamoto, A.; Kono, Y.; Fujita, T. The Impact of Breast Cancer Resistance Protein (BCRP/ABCG2) on Drug Transport Across Caco-2 Cell Monolayers. Drug Metab. Dispos. 2020, 48, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.J.; Huang, W.C.; Wei, Y.L.; Hsu, S.C.; Yuan, P.; Lin, H.Y.; Wistuba, I.I.; Lee, J.J.; Yen, C.J.; Su, W.C.; et al. Elevated BCRP/ABCG2 expression confers acquired resistance to gefitinib in wild-type EGFR-expressing cells. PLoS ONE 2011, 6, e21428. [Google Scholar] [CrossRef]
- Twentyman, P.R.; Luscombe, M. A Study of Some Variables in a Tetrazolium Dye (Mtt) Based Assay for Cell-Growth and Chemosensitivity. Br. J. Cancer 1987, 56, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.P.; Hsieh, Y.C.; Shia, C.S.; Hsu, P.W.; Chen, J.Y.; Hou, Y.C.; Hsieh, Y.W. Increased systemic exposure of methotrexate by a polyphenol-rich herb via modulation on efflux transporters multidrug resistance-associated protein 2 and breast cancer resistance protein. J. Pharm. Sci. 2016, 105, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Li, P.Y.; Liang, Y.C.; Sheu, M.J.; Huang, S.S.; Chao, C.Y.; Kuo, Y.H.; Huang, G.J. Alpinumisoflavone attenuates lipopolysaccharide-induced acute lung injury by regulating the effects of anti-oxidation and anti-inflammation both in vitro and in vivo. RSC Adv. 2018, 8, 31515–31528. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Chemical structures of MN (left) and HK (right).

Figure 2.
Effects of MN (μM), HK (μM) and Ko143 (0.25 μM, a positive control of BCRP inhibitor) on the intracellular accumulations of MXR in MDCKII-BCRP cells by using flow cytometry method. * p < 0.05, ** p < 0.01, *** p < 0.001. Control: 0.01% DMSO in reaction buffer.
Figure 2.
Effects of MN (μM), HK (μM) and Ko143 (0.25 μM, a positive control of BCRP inhibitor) on the intracellular accumulations of MXR in MDCKII-BCRP cells by using flow cytometry method. * p < 0.05, ** p < 0.01, *** p < 0.001. Control: 0.01% DMSO in reaction buffer.

Figure 3.
Intracellular accumulations of MN (100 μM) and HK (100 μM) in MDCKII-WT and MDCKII-BCRP cells after incubation for 30 min determined by HPLC-UV analysis and corrected with protein contents. ** p < 0.01.
Figure 3.
Intracellular accumulations of MN (100 μM) and HK (100 μM) in MDCKII-WT and MDCKII-BCRP cells after incubation for 30 min determined by HPLC-UV analysis and corrected with protein contents. ** p < 0.01.

Figure 4.
Effects of MN (12.5 μM) and HK (12.5 μM) on the protein expression of breast cancer resistance protein (BCRP) in MDCKII-WT and MDCKII-BCRP cells at 24 h and 48 h after incubation.
Figure 4.
Effects of MN (12.5 μM) and HK (12.5 μM) on the protein expression of breast cancer resistance protein (BCRP) in MDCKII-WT and MDCKII-BCRP cells at 24 h and 48 h after incubation.

Figure 5.
Effects of MN (12.5 μM) and HK (12.5 μM) on the protein expressions of p-EGFR, EGFR, p-PI3K, PI3K and BCRP, in MDCKII-BCRP cells at 24 h and 48 h after incubation. * p < 0.05, ** p < 0.01.
Figure 5.
Effects of MN (12.5 μM) and HK (12.5 μM) on the protein expressions of p-EGFR, EGFR, p-PI3K, PI3K and BCRP, in MDCKII-BCRP cells at 24 h and 48 h after incubation. * p < 0.05, ** p < 0.01.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, C.-P.; Li, P.-Y.; Chen, S.-Y.; Lin, S.-P.; Hou, Y.-C. Magnolol and Honokiol Inhibited the Function and Expression of BCRP with Mechanism Exploration. Molecules 2021, 26, 7390. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237390
AMA Style
Yu C-P, Li P-Y, Chen S-Y, Lin S-P, Hou Y-C. Magnolol and Honokiol Inhibited the Function and Expression of BCRP with Mechanism Exploration. Molecules. 2021; 26(23):7390. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237390
Chicago/Turabian StyleYu, Chung-Ping, Pei-Ying Li, Szu-Yu Chen, Shiuan-Pey Lin, and Yu-Chi Hou. 2021. "Magnolol and Honokiol Inhibited the Function and Expression of BCRP with Mechanism Exploration" Molecules 26, no. 23: 7390. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26237390