Long Non-Coding RNAs in Diffuse Large B-Cell Lymphoma
by
 , , and
, , and
Kasper Thystrup Karstensen
1 ,
,
Aleks Schein
1,
Andreas Petri
1,
Martin Bøgsted
2,3,
Karen Dybkær
2,3,
Shizuka Uchida
1,* and
and
Sakari Kauppinen
1,* 1
Center for RNA Medicine, Department of Clinical Medicine, Aalborg University, DK-2450 Copenhagen, Denmark
2
Department of Clinical Medicine, Faculty of Medicine, Aalborg University, DK-9000 Aalborg, Denmark
3
Department of Haematology, Clinical Cancer Research Center, Aalborg University Hospital, DK-9000 Aalborg, Denmark
*
Authors to whom correspondence should be addressed.
Non-Coding RNA 2021, 7(1), 1; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010001
Submission received: 28 October 2020
/
Revised: 21 December 2020
/
Accepted: 24 December 2020
/
Published: 28 December 2020
{kind=link}
{kind=link}
{kind=link}
Abstract
:Diffuse large B-cell lymphoma (DLBCL) is the most common lymphoid malignancy in adults. Although significant progress has been made in recent years to treat DLBCL patients, 30%–40% of the patients eventually relapse or are refractory to first line treatment, calling for better therapeutic strategies for DLBCL. Long non-coding RNAs (lncRNAs) have emerged as a highly diverse group of non-protein coding transcripts with intriguing molecular functions in human disease, including cancer. Here, we review the current understanding of lncRNAs in the pathogenesis and progression of DLBCL to provide an overview of the field. As the current knowledge of lncRNAs in DLBCL is still in its infancy, we provide molecular signatures of lncRNAs in DLBCL cell lines to assist further lncRNA research in DLBCL.
1. Introduction
Recent data implies that at least 80% of the human genome is transcribed, producing about 20,000 messenger RNAs (mRNAs), over 2300 microRNAs (miRNAs), a set of structural RNAs (such as ribosomal RNAs (rRNAs), transfer RNAs (tRNAs), and small nuclear RNAs (snRNAs)), and at least 50,000 long noncoding RNAs (lncRNAs) [1,2,3]. LncRNAs comprise a diverse group of non-protein coding transcripts that are longer than 200 nucleotides in length, lack an open reading frame, and regulate gene expression at many levels by affecting transcription, cellular localization, mRNA stability, translation, and other post-transcriptional events [4,5,6,7]. As a result, lncRNAs regulate a broad range of biological processes, including aging, cell growth and differentiation, hematopoiesis, and immune response [8,9,10,11]. Accordingly, numerous studies have reported highly diverse roles for lncRNAs in cancer [12,13,14], including development and maintenance of resistance to anti-cancer therapy [15,16,17,18]. Most prominent examples involve well-characterized lncRNAs, such as CDKN2B antisense RNA 1 (CDKN2B-AS1, also known as ANRIL) [19], growth arrest specific 5 (GAS5) [20,21], H19 imprinted maternally expressed transcript (H19) [22,23], HOX transcript antisense RNA (HOTAIR) [24,25], metastasis associated lung adenocarcinoma transcript 1 (MALAT1) [26,27], and X inactive specific transcript (XIST) [28,29]. However, the functions of lncRNAs in the pathogenesis of B-cell lymphomas are still poorly understood.
Lymphomas are a collective term used for malignancies of the lymph system, and they are divided into Hodgkin’s lymphomas (10% of lymphomas) and non-Hodgkin lymphomas (NHL), which account for ca. 90% of all lymphomas [30,31,32]. NHLs comprise a diverse group of lymphoid cancers with approx. 85%–90% of NHL cases originating from B cells, while the remaining NHLs derive from T cells or NK cells [33,34]. Diffuse large B-cell lymphoma (DLBCL) is the most common form of NHL, accounting for up to 40% of all lymphomas, with an incidence rate of approximately five to six cases per 100,000 Americans per year [35]. The 5-year survival rate ranges from 50%–80%, depending on risk profile, and the first-line treatment is the R-CHOP regimen (Rituximab plus Cyclophosphamide, Hydroxydaunorubicin, Oncovin, Prednisone combination therapy) [36]. Rituximab, a chimeric anti-CD20 monoclonal antibody [37] was added to the standard CHOP chemotherapy regimen in 2003, which improved the two-year progression-free survival from 51% to 61% and the overall survival rate from 52% to 78% [38,39]. Nevertheless, 30%–40% of the patients eventually relapse or are refractory to R-CHOP, experiencing poor two-year outcomes with overall survival rates of only 20%–40% [40,41], which calls for improved molecular understanding of the disease and new therapeutic strategies for the treatment of DLBCL. To this end, lncRNAs might be a key to further understand the diagnostic and prognostic measures for DLBCL to promote better treatment options for DLBCL patients. In this article, we summarize the current status of lncRNAs in DLBCL and provide a survey for lncRNAs in DLBCL cell lines to assist further research into the biological functions of lncRNAs in the pathogenesis of DLBCL.
2. General Functions of lncRNAs
LncRNAs exert their functions by binding to other macromolecules: DNA, RNA, or proteins. At the level of transcription, a lncRNA can form a scaffold for the epigenetic or transcription factor complex to assist its activation or repression of specific set of genes. The best investigated example is the binding between histone H3 lysine 27 methyltransferase, polycomb repressive complex 2 (PRC2) (i.e., the functional enzymatic component, enhancer of zeste homolog 2 (EZH2)) and lncRNAs such as HOTAIR [42], taurine up-regulated 1 (TUG1) [43], and XIST [44]. However, recent studies suggest that EZH2 binds many RNAs promiscuously, including lncRNAs [45,46,47,48,49,50]. Besides being transcriptional regulators, some lncRNAs can bind other RNAs to exert post-transcriptional regulation. For example, lncRNAs have been suggested to function as competitive endogenous RNAs (ceRNAs) to sequester miRNAs thereby influencing the amount of proteins being produced from the target mRNAs. Such lncRNAs are more commonly known as miRNA sponges, which include the H19 lncRNA sponging miR-675 [51], the hepatocellular carcinoma up-regulated long non-coding RNA (HULC) binding to miR-372 [52], and the ZNFX1 antisense RNA 1 (ZFAS1) sequestering miR-150 [53]. As for the translational control, lncRNAs can bind to RNA-binding proteins, which regulate mRNA translation. For example, in murine cardiomyocytes, the paternally imprinted lncRNA gene, antisense of IGF2R non-protein coding RNA (Airn, also known as Air), binds to the RNA-binding protein insulin like growth factor 2 mRNA binding protein 2 (Igf2bp2) to regulate the translational efficiency of hundreds of mRNAs to control the homeostasis of cardiomyocytes. In addition, lncRNA sequences themselves may be important for genomic imprinting, an epigenetic phenomenon through which genes are expressed in a parent-of-origin dependent manner, as in the case of the paternal imprinting lncRNA Airn [54] and the maternal imprinting lncRNA H19 [55]). Genomic lncRNA sequences may also contain important regulatory sequences, such as enhancers, as in the case of Hand2, opposite strand 1 (Hand2os1, also known as Upperhand [56]) and Hand2 downstream lncRNA (Hdnr, also known as Handsdown [57])].
Because of the broad definition of lncRNAs as non-protein-coding transcripts longer than 200 nucleotides in length, pseudogenes, which arise from the duplication of DNA sequences or retrotransposition and integration into the genome [58], are currently considered as a new category of lncRNAs [59]. Furthermore, circular RNAs (circRNAs) have emerged as an abundant class of covalently closed non-coding RNA molecules in eukaryotes that arise from backsplicing of protein-coding and non-coding exons and/or introns [60,61,62]. Due to the lack of free 5′ and 3′ ends, circRNAs are more resistant to exonuclease-mediated degradation compared to linear RNAs (e.g., mRNAs). Because of their stability, circRNAs accumulate in the blood, saliva, and urine, which have made them attractive candidates for biomarker discovery [63,64]. Finally, some lncRNAs have been shown to encode small peptides, named micropeptides. The examples of micropeptides are myomixer, myoblast fusion factor (MYMX) [65], and myoregulin (MRLN) [66] which regulate muscle formation and activity, respectively. Taken together, lncRNAs comprise a heterogeneous class of non-protein coding transcripts with highly diverse roles in biological processes. However, additional functional studies are required to establish the functions of lncRNAs in human diseases, such as cancer.
3. LncRNAs in DLBCL
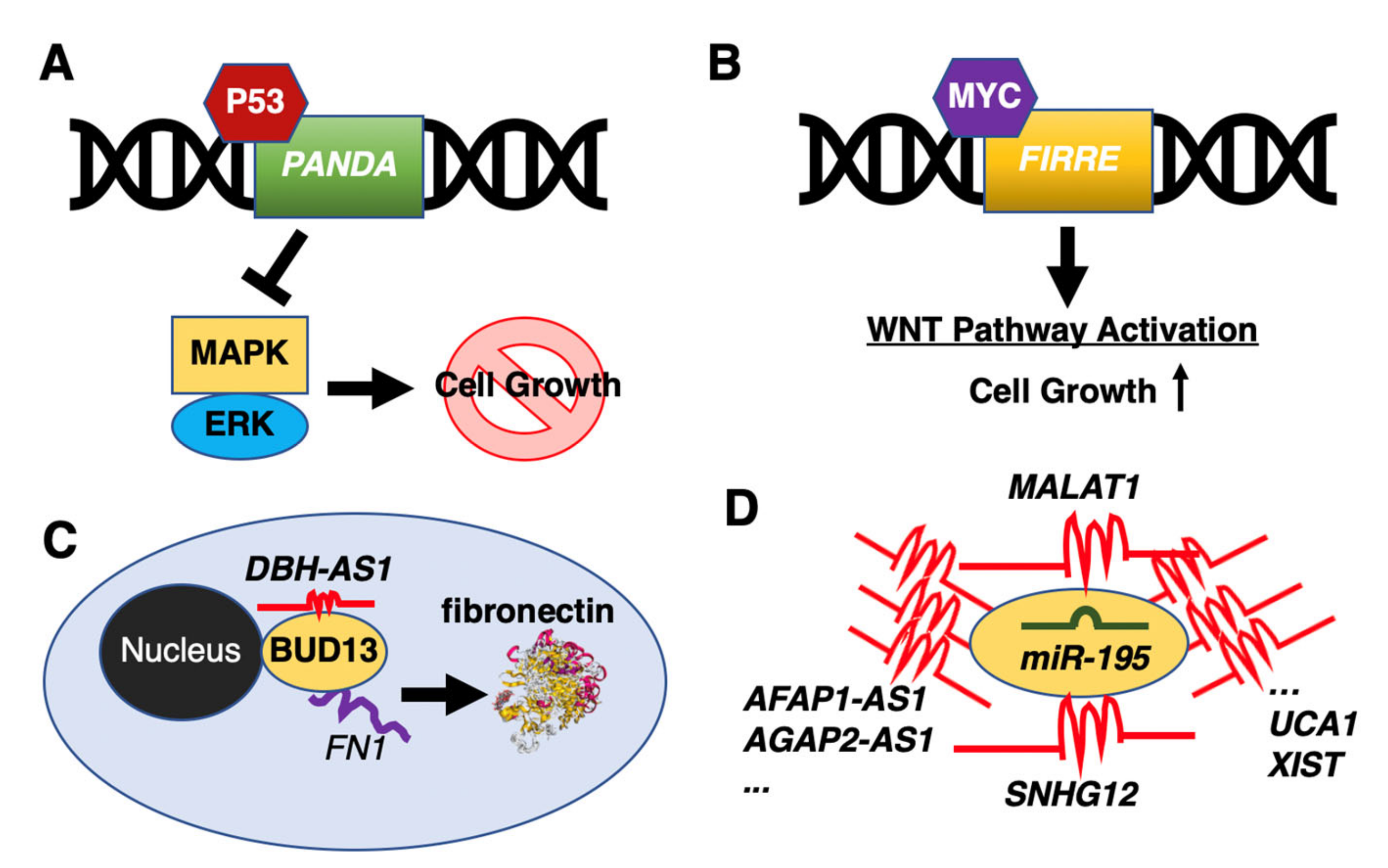
Due to the increased interest to study lncRNAs in recent years, researchers have also identified a number of differentially expressed lncRNAs in DLBCL patients compared to healthy donors or in DLBCL cell lines [67,68,69,70]. For example, the p53-activated lncRNA, promoter of CDKN1A antisense DNA damage activated RNA (PANDA), was reported to inactivate the MAPK/ERK signaling pathway to suppress cell growth by a G0/G1 cell cycle arrest [68] (Figure 1A). By comparison, the firre intergenic repeating RNA element (FIRRE) lncRNA was shown to function as an oncogene by activating Wnt/β-catenin signaling pathway via promoting nuclear translocation of β-catenin [71] (Figure 1B). Other lncRNAs, acting as post-transcriptional regulators, include the DBH antisense RNA 1 (DBH-AS1) binding to the RNA-binding protein, BUD13 homolog (BUD13), controlling the translation of fibronectin 1 (FN1) to positively regulate the proliferation, migration, and invasion of DLBCL cells [72] (Figure 1C). Another example is the EZH2-mediated lncRNA, FAS antisense RNA 1 (FAS-AS1), which binds to the RNA-binding protein, RNA binding motif protein 5 (RBM5), to control alternative splicing of Fas cell surface death receptor (FAS) mRNA [73]. Yet, other lncRNAs were shown to directly control protein modifications to influence tumor growth, as in the case of TUG1, which reduces protein levels of the MET proto-oncogene, receptor tyrosine kinase (MET) by promoting its ubiquitination [74].
Among various functions of lncRNAs, there is an increasing trend to investigate lncRNAs as miRNA sponges by viewing lncRNAs as an extra layer of the post-transcriptional regulatory machinery to fine-tune miRNA-mediated control of protein abundance. For example, the nuclear paraspeckle assembly transcript 1 (NEAT1) was shown to bind miR-34b-5p to affect the proliferation of DLBCL cell lines by targeting the GLI family zinc finger 1 (GLI1) [75]. Two lncRNAs have been implicated in the proliferation of DLBCL cells, the paternally expressed 10 (PEG10) sponging miR-101-3p, which targets kinesin family member 2A (KIF2A) [76], and SMAD5 antisense RNA 1 (SMAD5-AS1) that binds miR-135b-5bp to regulate the translation of the APC regulator of the WNT signaling pathway (APC) [77].
Small nucleolar RNAs (snoRNAs) are a class of small RNA molecules that assist in chemical modifications of other regulatory RNAs (e.g., rRNAs and tRNAs) [78,79]. Similar to miRNAs, snoRNAs are processed from much longer host genes, which are categorized as lncRNAs. In DLBCL, three snoRNA host genes have been proposed to function as miRNA sponges: small nucleolar RNA host gene 12 (SNHG12) sequesters miR-195 to control the cell growth, migration, and invasion of DLBCL cells in vitro [80]; small nucleolar RNA host gene 14 (SNHG14) sponges miR-5590-3p to upregulate Zinc finger E-box binding homeobox 1 (ZEB1); and small nucleolar RNA host gene 16 (SNHG16) sequesters miR-497-5p, to derepress the Pim-1 proto-oncogene, serine/threonine kinase (PIM1).
Since a single miRNA can modulate hundreds of mRNAs, it is not surprising that one miRNA can potentially be sequestered by several lncRNAs. MiR-195, which is a member of the miR-15/107 family, is a known tumor suppressor, whose dysregulation is linked to human diseases, including Alzheimer’s disease, cardiac hypertrophy, and many cancers [81,82]. As mentioned above [80], miR-195 is sequestered by SNHG12. In DLBCL, another lncRNA, MALAT1, was shown to regulate expression of the CD274 molecule (PD-L1) via miR-195 [83]. If the search is extended to outside of DLBCL, more than two dozen lncRNAs are suggested to sponge miR-195, including AFAP1 antisense RNA 1 (AFAP1-AS1) [84], AGAP2 antisense RNA 1 (AGAP2-AS1) [85], cytoskeleton regulator RNA (CYTOR) [86], maternally expressed 3 (MEG3) [87], NEAT1 [88], OIP5 antisense RNA 1 (OIP5-AS1) [89], urothelial cancer associated 1 (UCA1) [90,91], and XIST [92] to name a few (Figure 1D). Thus, it is clear that the studying of lncRNAs as miRNA sponges is far more complex than one would hope for, which suggests that it would be very challenging to use lncRNAs as miRNA sponges for therapeutic purposes.
4. Differential Expression of lncRNAs in Rituximab Sensitive and Resistant DLBCL Cell Lines
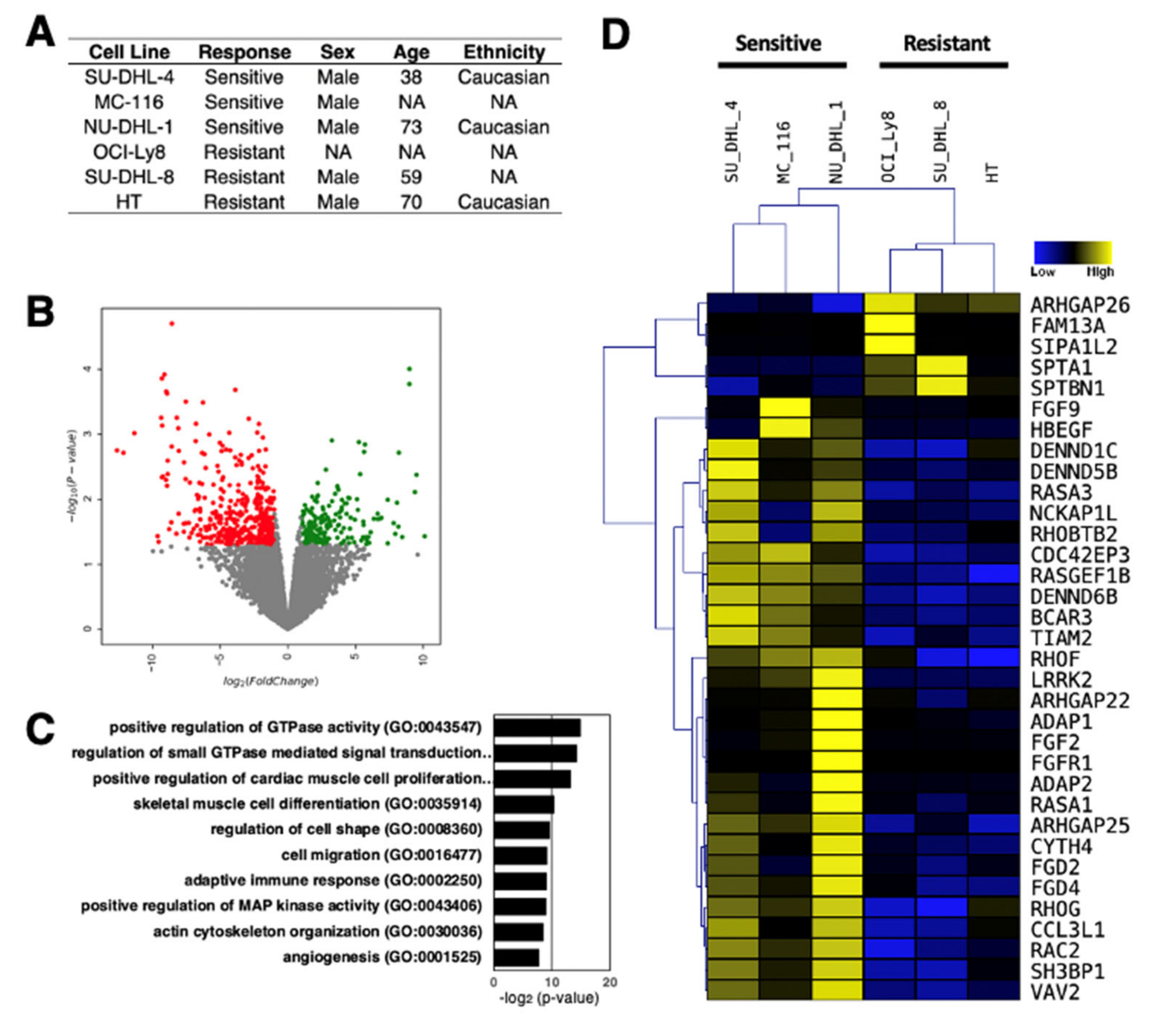
Understanding the role of lncRNAs in rituximab-resistant DLBCL phenotype could potentially guide the development of improved DLBCL therapies, thereby increase the survival rate of DLBCL patients, and cut the treatment costs. As evident from the current findings of lncRNAs in DLBCL, the number of lncRNAs identified and characterized is still limited. To provide further insights into the role of lncRNAs in DLBCL, we analyzed the transcription profiles of DLBCL cell lines, which are either sensitive or resistant to rituximab, by RNA-sequencing (Figure 2A). These line cells are from different individuals and were not treated with rituximab to uncover the intrinsic transcriptomic differences that underlie drug resistance. We uncovered 195 up- and 428 down-regulated genes in rituximab-resistant DLCBL cell lines compared to sensitive cell lines (Figure 2B), suggesting that there are large gene expression differences inherent to rituximab resistance. When Gene Ontology (GO) analysis for differentially expressed genes (both up- and down-regulated genes combined) was performed, several key signaling pathways known to be involved in tumor development were enriched (Figure 2C); most notably the GTPase signaling pathway [93,94] (Figure 2D), confirming the intrinsic differences in DLBCL cell lines compared.
Of the 458 differentially expressed genes, 123 are classified as lncRNAs by the latest annotation provided by the Ensembl database (GRCh38 version 100), comprising 54 up- and 69-down regulated lncRNAs (Supplementary Table S1). To initiate the dissection of the molecular signatures of the differentially expressed lncRNAs, additional bioinformatic analyses were performed. First, the promoter regions of the differentially expressed lncRNAs were examined for potential binding sites of transcription factors to uncover gene regulatory networks of lncRNA expression. Among the 350 transcription factors with potential binding sites in the promoter regions, 10 transcription factors were differentially expressed in rituximab-resistant compared to rituximab-sensitive DLCBL cell lines (p < 0.05) (Figure 3A; Supplementary Table S2). Among the differentially expressed transcription factors, the homeodomain transcription factor Meis homeobox 1 (MEIS1) is predicted to bind most sites in the highest number of promoter regions of the differentially expressed lncRNAs. Although MEIS1 is down-regulated in rituximab-resistant DLCBL cell lines, a recent study reported that the MYC proto-oncogene, bHLH transcription factor (MYC)-dependent down-regulation of MEIS1 is linked to the tumor development and progression via elevated homeobox B13 (HOXB13) expression and androgen receptor (AR) activity in prostate cancers [97], suggesting that the resistance to rituximab may be linked to the down-regulation of MEIS1, which, in turn, could regulate the expression of downstream lncRNAs. Furthermore, the Kruppel like factor 5 (KLF5) is the only up-regulated transcription factor in rituximab-resistant DLCBL cell lines. Interestingly, a previous report found KLF5 to be upregulated in human breast cancer cells treated with the HER2/epidermal growth factor receptor inhibitor, lapatinib [98]. Together with its family member KLF4, KLF5 was shown to induce expression of the anti-apoptotic factor, MCL1 (a BCL2 family member), to coordinate the gene regulatory program in resistance to lapatinib. Thus, further studies are warranted to uncover the importance of KLF5 in DLCBL in relation to regulation of lncRNA expression and rituximab resistance.
Besides lncRNAs functioning as miRNA sponges, increasing evidence suggests that many lncRNAs sequester RNA-binding proteins (RBPs) to influence mRNA degradation and stability. Such lncRNAs are collectively called RBP sponges [101]. Interestingly, the up-regulated lncRNAs in rituximab-resistant DLCBL cell lines harbor many putative RBP binding sites within their sequences (Supplementary Table S3). In particular, the cholesterol-induced regulator of metabolism RNA (CHROMR, also known as CHROME and PRKRA-AS1) has the fourth highest number of RBP binding sites among all up-regulated lncRNAs examined (Supplementary Table S3). A previous study showed that CHROMR expression is increased in the plasma and atherosclerotic plaques of individuals with coronary artery disease [102]. Mechanistically, CHROMR either sequesters or degrades a set of miRNAs (i.e., miR-27b, miR-33a, miR-33b, and miR-128) to repress the expression of genes mediating cholesterol transport in human hepatocytes and macrophages [102]. As cholesterol metabolism has been reported to drive tumor growth and invasion [103], it would be of high interest to study the intertwined link between cholesterol metabolism and DLCBL [104]. Given that many RBPs are involved in cholesterol metabolism and CHROMR is 3.4-fold up-regulated in rituximab-resistant DLCBL cell lines [103], in addition to being miRNA sponge, CHROMR could function as an RBP sponge to modulate the translation of genes mediating cholesterol transport (Figure 3B).
5. Materials and Methods
5.1. Cell Culture and Treatment
In this study, the following six human DLBCL-derived cell lines were used: NU-DHL-1, HT, MC-116, SU-DHL-4 (DSMZ, German Collection of Microorganisms and Cell Cultures); OCI-Ly8, and SU-DHL-8 (provided by Dr. Jose A. Martinez-Climent, Molecular Oncology Laboratory, University of Navarra, Pamplona, Spain). The cell lines were cultured under standard conditions at 37 °C in a humidified atmosphere of 95% air and 5% CO2 with RPMI-1640 medium containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (P/S) for no longer than 20 passages. All cell lines were authenticated by DNA barcoding, as previously described [95,96].
5.2. RNA Isolation and RNA-seq Assay
Total RNA was extracted using a modified protocol combining TRIzol Reagent (Invitrogen, Paisley, UK) and mirVana miRNA Isolation Kit (Ambion/ThermoFisher Scientific, Grand Island, NY, USA), as previously described [105]. The RNA quality and concentration of each sample were determined by Agilent 2100 Bioanalyzer analysis (Agilent Technologies, Santa Clara, CA, USA) and NanoDrop ND-1000 spectrophotometer (ThermoFisher Scientific), respectively. The total RNA of each sample was sent to AROS Applied Biotechnology AS (Aarhus, Denmark) for poly-A selected, pair-end RNA-seq via Illumina HiSeq 2000 platform. The generated and analyzed data were deposited in the Gene Expression Omnibus (accession ID: GSE159852)
5.3. Data Analysis
For RNA-seq data analysis, fastp [106] was used to trim the first seven base pairs, detect paired-end adapters, and analyze overrepresented sequences. After quality control, the trimmed reads were aligned against GRCh38 genome build (version 100 from the Ensembl database) using the Spliced Transcripts Alignment to a Reference (STAR) software [107]. Differential expression analysis was performed using edgeR [108]. The Trimmed Mean of M-values (TMM) method was used to normalize the data to obtain counts per million reads mapped (CPM) values.
A volcano plot was generated using Zenodo [109]. To draw heat maps and run hierarchical clustering, MultiExperiment Viewer (MeV) [110] was used. The bindings of transcription factors were predicted via CiiiDER [111] with its default settings, including the analysis at −1500 bases upstream to +500 bases downstream of the transcription start site (TSS) of each lncRNA. The putative bindings of RBPs were downloaded from the oRNAment database [112]. The network of RBPs was drawn via NetworkAnalyst [100].
6. Concluding Remarks
This review summarizes recent findings of lncRNAs in DLBCL. Although several studies of lncRNAs in DLBCL have been published [67,68,69,70,113,114], the number of functionally studied lncRNAs is still limited, which precludes the defined diagnostic and prognostic importance of lncRNAs in DLBCL patients. To this end, we provide molecular signatures of lncRNAs in rituximab-resistant and -sensitive DLCBL cell lines, respectively. A study published in 2017 identifies 17 lncRNAs that can discriminate between two major molecular subtypes of DLBCL, activated B-cell-like (ABC) and germinal center B-cell-like (GCB), with high specificity, which the authors termed these seventeen lncRNAs as SubSigLnc-17. However, in our RNA-seq data, SubSigLnc-17 were not differentially expressed, which promoted us to further analyze the differentially expressed lncRNAs in our RNA-seq data [115]. We report on initial bioinformatic analyses of the differentially expressed lncRNAs; however, further biological validation and functional studies are required to uncover the biological roles of the identified lncRNAs in DLCBL.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2311-553X/7/1/1/s1, Supplementary Table S1: List of differentially expressed lncRNAs in rituximab-resistant compared to -sensitive DLCBL cell lines (n = 3 biologically independent samples with the threshold values of 2-fold and p < 0.05). Supplementary Table S2: List of differentially expressed transcription factors and their binding to the promoter regions of differentially expressed lncRNAs. Supplementary Table S3: List of expressed RBPs and their binding to the up-regulated lncRNAs. Each number indicates the number of binding sites predicted to bound by each RBP shown.
Author Contributions
Conceptualization, M.B., K.D., and S.K.; methodology, M.B., K.D., and S.K.; resources, K.D. and S.K.; data analysis, K.T.K., A.S., A.P., and S.U.; writing—Original draft preparation, S.U. and S.K.; writing—Review and editing, K.D.; funding acquisition, S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Karen Elise Jensen Foundation and Novo Nordisk Foundation (S.K.).
Institutional Review Board Statement
Ethical review and approval were waived for this study, due to the usage of established human cell lines.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data sets analyzed in this study can be found in the Zenodo repository (https://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.4146742) and in the Gene Expression Omnibus repository (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/) with the reference number GSE159852.
Acknowledgments
We thank the members of Center for RNA Medicine for helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Uchida, S.; Adams, J.C. Physiological roles of non-coding RNAs. Am. J. Physiol. Cell Physiol. 2019, 317, C1–C2. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grasser, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uszczynska-Ratajczak, B.; Lagarde, J.; Frankish, A.; Guigo, R.; Johnson, R. Towards a complete map of the human long non-coding RNA transcriptome. Nat. Rev. Genet. 2018, 19, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Carninci, P. Genome-wide technologies to study RNA-chromatin interactions. Non-Coding RNA 2020, 6. [Google Scholar] [CrossRef]
- Maass, P.G.; Barutcu, A.R.; Rinn, J.L. Interchromosomal interactions: A genomic love story of kissing chromosomes. J. Cell Biol. 2019, 218, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.; Kanduri, C. Understanding long noncoding RNA and chromatin interactions: What we know so far. Non-Coding RNA 2019, 5. [Google Scholar] [CrossRef] [Green Version]
- Lennox, K.A.; Behlke, M.A. Cellular localization of long non-coding RNAs affects silencing by RNAi more than by antisense oligonucleotides. Nucleic Acids Res. 2016, 44, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Ang, C.E.; Trevino, A.E.; Chang, H.Y. Diverse lncRNA mechanisms in brain development and disease. Curr. Opin. Genet. Dev. 2020, 65, 42–46. [Google Scholar] [CrossRef]
- Bar, C.; Chatterjee, S.; Falcao Pires, I.; Rodrigues, P.; Sluijter, J.P.G.; Boon, R.A.; Nevado, R.M.; Andres, V.; Sansonetti, M.; de Windt, L.; et al. Non-coding RNAs: Update on mechanisms and therapeutic targets from the ESC working groups of myocardial function and cellular biology of the heart. Cardiovasc. Res. 2020, 116, 1805–1819. [Google Scholar] [CrossRef]
- Dominguez-Andres, J.; Fanucchi, S.; Joosten, L.A.B.; Mhlanga, M.M.; Netea, M.G. Advances in understanding molecular regulation of innate immune memory. Curr. Opin. Cell. Biol. 2020, 63, 68–75. [Google Scholar] [CrossRef]
- Pereira Fernandes, D.; Bitar, M.; Jacobs, F.M.J.; Barry, G. Long Non-Coding RNAs in Neuronal Aging. Non-Coding RNA 2018, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmeron-Barcenas, E.G.; Mendoza-Catalan, M.A.; Illades-Aguiar, B.; Peralta-Arrieta, I.; Alquisiras-Burgos, I.; Ortiz-Ortiz, J.; Navarro-Tito, N.; Zacapala-Gomez, A.E. Long non-coding RNAs as new players in cervical carcinogenesis: An update. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8314–8328. [Google Scholar] [CrossRef] [PubMed]
- Uchida, S.; Kauppinen, S. Long non-coding RNAs in liver cancer and nonalcoholic steatohepatitis. Non-Coding RNA 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Ginn, L.; Shi, L.; Montagna, M.; Garofalo, M. LncRNAs in non-small-cell lung cancer. Non-Coding RNA 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Smolle, M.A.; Prinz, F.; Calin, G.A.; Pichler, M. Current concepts of non-coding RNA regulation of immune checkpoints in cancer. Mol. Aspects Med. 2019, 70, 117–126. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, J.; Yu, J.; Liu, X.; Yu, C.; Hu, J.; Shi, H.; Ma, X. Long non-coding RNAs: Emerging roles in the Immunosuppressive tumor microenvironment. Front. Oncol. 2020, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Kamal, M.A.; Yuan, C. The regulatory roles of long non-coding RNA in the chemoresistance process of ovarian cancer. Curr. Pharm. Des. 2019, 25, 856–861. [Google Scholar] [CrossRef]
- Bai, H.; Lei, K.; Huang, F.; Jiang, Z.; Zhou, X. Exo-circRNAs: A new paradigm for anticancer therapy. Mol. Cancer 2019, 18, 56. [Google Scholar] [CrossRef]
- Kong, Y.; Hsieh, C.H.; Alonso, L.C. ANRIL: A lncRNA at the CDKN2A/B locus with roles in cancer and metabolic disease. Front. Endocrinol. 2018, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Xie, Z.; Lei, X.; Gan, R. Long non-coding RNA GAS5 in human cancer. Oncol. Lett. 2020, 20, 2587–2594. [Google Scholar] [CrossRef]
- Yu, Y.; Hann, S.S. Novel tumor suppressor lncRNA growth arrest-specific 5 (GAS5) in human cancer. Onco. Targets Ther. 2019, 12, 8421–8436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Shen, A.; Liu, A. Long non-coding RNA H19 and cancer: A competing endogenous RNA. Bull. Cancer 2019, 106, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Tietze, L.; Kessler, S.M. The good, the bad, the question-H19 in hepatocellular carcinoma. Cancers 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Cantile, M.; Di Bonito, M.; Cerrone, M.; Collina, F.; De Laurentiis, M.; Botti, G. Long non-coding RNA HOTAIR in breast cancer therapy. Cancers 2020, 12. [Google Scholar] [CrossRef]
- Qu, X.; Alsager, S.; Zhuo, Y.; Shan, B. HOX transcript antisense RNA (HOTAIR) in cancer. Cancer Lett. 2019, 454, 90–97. [Google Scholar] [CrossRef]
- Amodio, N.; Raimondi, L.; Juli, G.; Stamato, M.A.; Caracciolo, D.; Tagliaferri, P.; Tassone, P. MALAT1: A druggable long non-coding RNA for targeted anti-cancer approaches. J. Hematol. Oncol. 2018, 11, 63. [Google Scholar] [CrossRef] [Green Version]
- Arun, G.; Aggarwal, D.; Spector, D.L. MALAT1 long non-coding RNA: Functional implications. Non-Coding RNA 2020, 6. [Google Scholar] [CrossRef]
- Chen, Y.K.; Yen, Y. The ambivalent role of lncRNA Xist in carcinogenesis. Stem Cell Rev. Rep. 2019, 15, 314–323. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, X.; Jiang, X.; Zhao, H. X-inactive-specific transcript: A long noncoding RNA with complex roles in human cancers. Gene 2018, 679, 28–35. [Google Scholar] [CrossRef]
- Ansell, S.M. Hodgkin lymphoma: A 2020 update on diagnosis, risk-stratification, and management. Am. J. Hematol. 2020, 95, 978–989. [Google Scholar] [CrossRef]
- Shanbhag, S.; Ambinder, R.F. Hodgkin lymphoma: A review and update on recent progress. CA Cancer J. Clin. 2018, 68, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310. [Google Scholar] [CrossRef]
- Types of B-cell Lymphoma. Available online: https://www.cancer.org/cancer/non-hodgkin-lymphoma/about/b-cell-lymphoma.html (accessed on 22 October 2020).
- Salles, G.; Barrett, M.; Foa, R.; Maurer, J.; O’Brien, S.; Valente, N.; Wenger, M.; Maloney, D.G. Rituximab in B-cell hematologic malignancies: A review of 20 years of clinical experience. Adv. Ther. 2017, 34, 2232–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. SEER Cancer Statistics Review, 1975–2017. Available online: https://seer.cancer.gov/csr/1975_2017/ (accessed on 14 September 2020).
- Shaw, J.; Harvey, C.; Richards, C.; Kim, C. Temporal trends in treatment and survival of older adult diffuse large B-cell lymphoma patients in the SEER-medicare linked database. Leuk. Lymphoma 2019, 60, 3235–3243. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, P.; Grillo-Lopez, A.J.; Link, B.K.; Levy, R.; Czuczman, M.S.; Williams, M.E.; Heyman, M.R.; Bence-Bruckler, I.; White, C.A.; Cabanillas, F.; et al. Rituximab chimeric anti-CD20 monoclonal antibody therapy for relapsed indolent lymphoma: Half of patients respond to a four-dose treatment program. J. Clin. Oncol. 1998, 16, 2825–2833. [Google Scholar] [CrossRef]
- Sehn, L.H.; Donaldson, J.; Chhanabhai, M.; Fitzgerald, C.; Gill, K.; Klasa, R.; MacPherson, N.; O’Reilly, S.; Spinelli, J.J.; Sutherland, J.; et al. Introduction of combined CHOP plus rituximab therapy dramatically improved outcome of diffuse large B-cell lymphoma in British Columbia. J. Clin. Oncol. 2005, 23, 5027–5033. [Google Scholar] [CrossRef]
- Vose, J.M.; Link, B.K.; Grossbard, M.L.; Czuczman, M.; Grillo-Lopez, A.; Gilman, P.; Lowe, A.; Kunkel, L.A.; Fisher, R.I. Phase II study of rituximab in combination with chop chemotherapy in patients with previously untreated, aggressive non-Hodgkin’s lymphoma. J. Clin. Oncol. 2001, 19, 389–397. [Google Scholar] [CrossRef]
- Crump, M.; Neelapu, S.S.; Farooq, U.; Van Den Neste, E.; Kuruvilla, J.; Westin, J.; Link, B.K.; Hay, A.; Cerhan, J.R.; Zhu, L.; et al. Outcomes in refractory diffuse large B-cell lymphoma: Results from the international SCHOLAR-1 study. Blood 2017, 130, 1800–1808. [Google Scholar] [CrossRef]
- Rovira, J.; Valera, A.; Colomo, L.; Setoain, X.; Rodriguez, S.; Martinez-Trillos, A.; Gine, E.; Dlouhy, I.; Magnano, L.; Gaya, A.; et al. Prognosis of patients with diffuse large B cell lymphoma not reaching complete response or relapsing after frontline chemotherapy or immunochemotherapy. Ann. Hematol. 2015, 94, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.B.; Yin, D.D.; Sun, M.; Kong, R.; Liu, X.H.; You, L.H.; Han, L.; Xia, R.; Wang, K.M.; Yang, J.S.; et al. P53-regulated long non-coding RNA TUG1 affects cell proliferation in human non-small cell lung cancer, partly through epigenetically regulating HOXB7 expression. Cell Death Dis. 2014, 5, e1243. [Google Scholar] [CrossRef] [PubMed]
- Plath, K.; Fang, J.; Mlynarczyk-Evans, S.K.; Cao, R.; Worringer, K.A.; Wang, H.; de la Cruz, C.C.; Otte, A.P.; Panning, B.; Zhang, Y. Role of histone H3 lysine 27 methylation in X inactivation. Science 2003, 300, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Paucek, R.D.; Gooding, A.R.; Brown, Z.Z.; Ge, E.J.; Muir, T.W.; Cech, T.R. Molecular analysis of PRC2 recruitment to DNA in chromatin and its inhibition by RNA. Nat. Struct. Mol. Biol. 2017, 24, 1028–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Goodrich, K.J.; Gooding, A.R.; Naeem, H.; Archer, S.; Paucek, R.D.; Youmans, D.T.; Cech, T.R.; Davidovich, C. Targeting of polycomb repressive complex 2 to RNA by short repeats of consecutive guanines. Mol. Cell 2017, 65, 1056–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portoso, M.; Ragazzini, R.; Brencic, Z.; Moiani, A.; Michaud, A.; Vassilev, I.; Wassef, M.; Servant, N.; Sargueil, B.; Margueron, R. PRC2 is dispensable for HOTAIR-mediated transcriptional repression. EMBO J. 2017, 36, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Davidovich, C.; Wang, X.; Cifuentes-Rojas, C.; Goodrich, K.J.; Gooding, A.R.; Lee, J.T.; Cech, T.R. Toward a consensus on the binding specificity and promiscuity of PRC2 for RNA. Mol. Cell 2015, 57, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Rojas, C.; Hernandez, A.J.; Sarma, K.; Lee, J.T. Regulatory interactions between RNA and polycomb repressive complex 2. Mol. Cell 2014, 55, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Davidovich, C.; Zheng, L.; Goodrich, K.J.; Cech, T.R. Promiscuous RNA binding by polycomb repressive complex 2. Nat. Struct. Mol. Biol. 2013, 20, 1250–1257. [Google Scholar] [CrossRef] [Green Version]
- Tsang, W.P.; Ng, E.K.; Ng, S.S.; Jin, H.; Yu, J.; Sung, J.J.; Kwok, T.T. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis 2010, 31, 350–358. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Wu, H.; Ni, P.; Gu, Z.; Qiao, Y.; Chen, N.; Sun, F.; Fan, Q. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucleic Acids Res. 2010, 38, 5366–5383. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Xie, J.; Shen, C.; Cheng, D.; Shi, Y.; Wu, Z.; Deng, X.; Chen, H.; Shen, B.; Peng, C.; et al. Amplification of long noncoding RNA ZFAS1 promotes metastasis in hepatocellular carcinoma. Cancer Res. 2015, 75, 3181–3191. [Google Scholar] [CrossRef] [Green Version]
- Latos, P.A.; Pauler, F.M.; Koerner, M.V.; Senergin, H.B.; Hudson, Q.J.; Stocsits, R.R.; Allhoff, W.; Stricker, S.H.; Klement, R.M.; Warczok, K.E.; et al. Airn transcriptional overlap, but not its lncRNA products, induces imprinted Igf2r silencing. Science 2012, 338, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Monnier, P.; Martinet, C.; Pontis, J.; Stancheva, I.; Ait-Si-Ali, S.; Dandolo, L. H19 lncRNA controls gene expression of the imprinted gene network by recruiting MBD1. Proc. Natl. Acad. Sci. USA 2013, 110, 20693–20698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.M.; Anderson, D.M.; McAnally, J.R.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Transcription of the non-coding RNA upperhand controls Hand2 expression and heart development. Nature 2016, 539, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, N.; Ali, T.; Kopitchinski, N.; Schuster, P.; Beisaw, A.; Hendrix, D.A.; Schulz, M.H.; Muller-McNicoll, M.; Dimmeler, S.; Grote, P. The lncRNA locus handsdown regulates cardiac gene programs and is essential for early mouse development. Dev. Cell 2019, 50, 644–657. [Google Scholar] [CrossRef]
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Carter, D.R. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 2011, 17, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Grander, D.; Johnsson, P. Pseudogene-expressed RNAs: Emerging roles in gene regulation and disease. Curr. Top. Microbiol. Immunol. 2016, 394, 111–126. [Google Scholar] [CrossRef]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in cancer. Nat. Rev. Cancer 2020, 21, 10–1038. [Google Scholar] [CrossRef]
- Li, F.; Yang, Q.; He, A.T.; Yang, B.B. Circular RNAs in cancer: Limitations in functional studies and diagnostic potential. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Lei, B.; Tian, Z.; Fan, W.; Ni, B. Circular RNA: A novel biomarker and therapeutic target for human cancers. Int. J. Med. Sci. 2019, 16, 292–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising biomarkers for human diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, P.; Ramirez-Martinez, A.; Li, H.; Cannavino, J.; McAnally, J.R.; Shelton, J.M.; Sanchez-Ortiz, E.; Bassel-Duby, R.; Olson, E.N. Control of muscle formation by the fusogenic micropeptide myomixer. Science 2017, 356, 323–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Anderson, K.M.; Chang, C.L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; et al. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; He, Y.; Zhang, H.; Wu, Z.; Li, D.; Zheng, C. Comprehensive analysis of differentially expressed profiles of lncRNAs and mRNAs reveals ceRNA networks in the transformation of diffuse large B-cell lymphoma. Oncol. Lett. 2018, 16, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, M.; Xu, H.; Wang, Y.; Li, Z.; Chang, Y.; Wang, X.; Fu, X.; Zhou, Z.; Yang, S.; et al. Discovery and validation of the tumor-suppressive function of long noncoding RNA PANDA in human diffuse large B-cell lymphoma through the inactivation of MAPK/ERK signaling pathway. Oncotarget 2017, 8, 72182–72196. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.Y.; Wu, B.; Yan, W.; Gong, Z.M.; Sun, Q.; Wang, H.H.; Yang, W. Microarray expression profiles of long non-coding RNAs in germinal center-like diffuse large B-cell lymphoma. Oncol Rep. 2017, 38, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Fang, C.; Li, X.; Geng, Y.; Li, R.; Wu, C.; Jiang, J.; Wu, C. Predictive analysis of long non-coding RNA expression profiles in diffuse large B-cell lymphoma. Oncotarget 2017, 8, 23228–23236. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Cui, Z.; Liu, X.; Wu, S.; Wu, Y.; Fang, F.; Zhao, H. LncRNA FIRRE is activated by MYC and promotes the development of diffuse large B-cell lymphoma via Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2019, 510, 594–600. [Google Scholar] [CrossRef]
- Song, Y.; Gao, F.; Peng, Y.; Yang, X. Long non-coding RNA DBH-AS1 promotes cancer progression in diffuse large B-cell lymphoma by targeting FN1 via RNA-binding protein BUD13. Cell. Biol. Int. 2020, 44, 1331–1340. [Google Scholar] [CrossRef]
- Sehgal, L.; Mathur, R.; Braun, F.K.; Wise, J.F.; Berkova, Z.; Neelapu, S.; Kwak, L.W.; Samaniego, F. FAS-antisense 1 lncRNA and production of soluble versus membrane Fas in B-cell lymphoma. Leukemia 2014, 28, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Yan, Z.; Wang, X.; Cao, J.; Chen, W.; Qi, K.; Zhou, D.; Xia, J.; Qi, N.; Li, Z.; et al. Downregulation of long non-coding RNA TUG1 suppresses tumor growth by promoting ubiquitination of MET in diffuse large B-cell lymphoma. Mol. Cell Biochem. 2019, 461, 47–56. [Google Scholar] [CrossRef]
- Qian, C.S.; Li, L.J.; Huang, H.W.; Yang, H.F.; Wu, D.P. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. 2020, 20, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Su, L.; Jiang, J. Long non-coding RNA paternally expressed imprinted gene 10 (PEG10) elevates diffuse large B-cell lymphoma progression by regulating kinesin family member 2A (KIF2A) via Targeting MiR-101-3p. Med. Sci. Monit. 2020, 26, e922810. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.C.; Jiao, Y.; Zhang, Y.Y.; Ning, J.; Zhang, Y.R.; Xu, J.; Wei, W.; Kang-Sheng, G. Lnc SMAD5-AS1 as ceRNA inhibit proliferation of diffuse large B cell lymphoma via Wnt/ β -catenin pathway by sponging miR-135b-5p to elevate expression of APC. Cell Death Dis. 2019, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.; Malla, S.; Lyons, S.M. snoRNPs: Functions in ribosome biogenesis. Biomolecules 2020, 10, 783. [Google Scholar] [CrossRef]
- Yang, H.; Jiang, Z.; Wang, S.; Zhao, Y.; Song, X.; Xiao, Y.; Yang, S. Long non-coding small nucleolar RNA host genes in digestive cancers. Cancer Med. 2019, 8, 7693–7704. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Zhang, X.M.; Han, B.Q.; Dai, H.B. Long noncoding RNA SNHG12 indicates the prognosis and accelerates tumorigenesis of diffuse large B-cell lymphoma through sponging microR-195. Onco. Targets Ther. 2020, 13, 5563–5574. [Google Scholar] [CrossRef]
- Loh, H.Y.; Norman, B.P.; Lai, K.S.; Rahman, N.; Alitheen, N.B.M.; Osman, M.A. The regulatory role of microRNAs in breast cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Liang, X.; Li, X.; Zhang, Y.; Sun, Z.; Liu, Y.; Wang, J. MicroRNA-195: A review of its role in cancers. Onco. Targets Ther. 2018, 11, 7109–7123. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; Lian, G.Y.; Song, Y.; Huang, Y.F.; Gong, Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. 2019, 231, 116335. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Wu, A.; Yu, H.; Yu, Q.; Zheng, B.; Yang, W.; Tian, D.; Li, P.; Gao, Y. Involvement of the lncRNA AFAP1-AS1/microRNA-195/E2F3 axis in proliferation and migration of enteric neural crest stem cells of Hirschsprung’s disease. Exp. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, K.; Liu, Y.; Liu, X.; Liu, B.; Ba, Y.; Xing, W. Silencing lncRNA AGAP2-AS1 Upregulates miR-195-5p to repress migration and invasion of EC cells via the decrease of FOSL1 expression. Mol. Ther. Nucleic Acids 2020, 20, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, W. Long noncoding RNA CYTOR sponges miR-195 to modulate proliferation, migration, invasion and radiosensitivity in nonsmall cell lung cancer cells. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Song, D. LncRNA MEG3 Participates in caerulein-induced inflammatory injury in human pancreatic cells via regulating miR-195-5p/FGFR2 axis and inactivating NF- κB pathway. Inflammation 2020. [Google Scholar] [CrossRef]
- Tang, N.; Dong, Y.; Liu, J.; Zhao, H. Silencing of long non-coding RNA NEAT1 upregulates miR-195a to attenuate intervertebral disk degeneration via the BAX/BAK pathway. Front. Mol. Biosci. 2020, 7, 147. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, T.; Tian, L.; Li, Y. LncRNA OIP5-AS1 promotes the proliferation of hemangioma vascular endothelial cells via regulating miR-195-5p/NOB1 axis. Front. Pharmacol. 2019, 10, 449. [Google Scholar] [CrossRef]
- Li, H.J.; Sun, X.M.; Li, Z.K.; Yin, Q.W.; Pang, H.; Pan, J.J.; Li, X.; Chen, W. LncRNA UCA1 promotes mitochondrial function of bladder cancer via the MiR-195/ARL2 signaling pathway. Cell Physiol. Biochem. 2017, 43, 2548–2561. [Google Scholar] [CrossRef]
- Yin, D.; Fu, C.; Sun, D. Silence of lncRNA UCA1 represses the growth and tube formation of human microvascular endothelial cells through miR-195. Cell Physiol. Biochem. 2018, 49, 1499–1511. [Google Scholar] [CrossRef]
- Yang, C.; Wu, K.; Wang, S.; Wei, G. Long non-coding RNA XIST promotes osteosarcoma progression by targeting YAP via miR-195-5p. J. Cell Biochem. 2018, 119, 5646–5656. [Google Scholar] [CrossRef]
- Chang, K.C.; Chang, W.C.; Chang, Y.; Hung, L.Y.; Lai, C.H.; Yeh, Y.M.; Chou, Y.W.; Chen, C.H. Ran GTPase-activating protein 1 is a therapeutic target in diffuse large B-cell lymphoma. PLoS ONE 2013, 8, e79863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouw, L.G.; Reading, N.S.; Jenson, S.D.; Lim, M.S.; Elenitoba-Johnson, K.S. Expression of the Rho-family GTPase gene RHOF in lymphocyte subsets and malignant lymphomas. Br. J. Haematol. 2005, 129, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.B.; Reinholdt, L.; Schonherz, A.A.; Due, H.; Jespersen, D.S.; Grubach, L.; Ettrup, M.S.; Roge, R.; Falgreen, S.; Sorensen, S.; et al. High CXCR4 expression impairs rituximab response and the prognosis of R-CHOP-treated diffuse large B-cell lymphoma patients. Oncotarget 2019, 10, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Due, H.; Brondum, R.F.; Young, K.H.; Bogsted, M.; Dybkaer, K. MicroRNAs associated to single drug components of R-CHOP identifies diffuse large B-cell lymphoma patients with poor outcome and adds prognostic value to the international prognostic index. BMC Cancer 2020, 20, 237. [Google Scholar] [CrossRef]
- Whitlock, N.C.; Trostel, S.Y.; Wilkinson, S.; Terrigino, N.T.; Hennigan, S.T.; Lake, R.; Carrabba, N.V.; Atway, R.; Walton, E.D.; Gryder, B.E.; et al. MEIS1 down-regulation by MYC mediates prostate cancer development through elevated HOXB13 expression and AR activity. Oncogene 2020, 39, 5663–5674. [Google Scholar] [CrossRef]
- Farrugia, M.K.; Sharma, S.B.; Lin, C.C.; McLaughlin, S.L.; Vanderbilt, D.B.; Ammer, A.G.; Salkeni, M.A.; Stoilov, P.; Agazie, Y.M.; Creighton, C.J.; et al. Regulation of anti-apoptotic signaling by Kruppel-like factors 4 and 5 mediates lapatinib resistance in breast cancer. Cell Death Dis. 2015, 6, e1699. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]
- HafezQorani, S.; Houdjedj, A.; Arici, M.; Said, A.; Kazan, H. RBPSponge: Genome-wide identification of lncRNAs that sponge RBPs. Bioinformatics 2019, 35, 4760–4763. [Google Scholar] [CrossRef]
- Hennessy, E.J.; van Solingen, C.; Scacalossi, K.R.; Ouimet, M.; Afonso, M.S.; Prins, J.; Koelwyn, G.J.; Sharma, M.; Ramkhelawon, B.; Carpenter, S.; et al. The long noncoding RNA CHROME regulates cholesterol homeostasis in primate. Nat. Metab. 2019, 1, 98–110. [Google Scholar] [CrossRef]
- Huang, B.; Song, B.L.; Xu, C. Cholesterol metabolism in cancer: Mechanisms and therapeutic opportunities. Nat. Metab. 2020, 2, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Rink, J.S.; Yang, S.; Cen, O.; Taxter, T.; McMahon, K.M.; Misener, S.; Behdad, A.; Longnecker, R.; Gordon, L.I.; Thaxton, C.S. rational targeting of cellular cholesterol in diffuse large B-cell lymphoma (DLBCL) enabled by functional lipoprotein nanoparticles: A therapeutic strategy dependent on cell of origin. Mol. Pharm. 2017, 14, 4042–4051. [Google Scholar] [CrossRef] [PubMed]
- Dybkaer, K.; Bogsted, M.; Falgreen, S.; Bodker, J.S.; Kjeldsen, M.K.; Schmitz, A.; Bilgrau, A.E.; Xu-Monette, Z.Y.; Li, L.; Bergkvist, K.S.; et al. Diffuse large B-cell lymphoma classification system that associates normal B-cell subset phenotypes with prognosis. J. Clin. Oncol. 2015, 33, 1379–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedre, R. reneshbedre/bioinfokit: Bioinformatics Data Analysis and Visualization Toolkit. Available online: https://zenodo.org/record/3747737#.X-SnxthKhPY (accessed on 13 September 2020).
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gearing, L.J.; Cumming, H.E.; Chapman, R.; Finkel, A.M.; Woodhouse, I.B.; Luu, K.; Gould, J.A.; Forster, S.C.; Hertzog, P.J. CiiiDER: A tool for predicting and analysing transcription factor binding sites. PLoS ONE 2019, 14, e0215495. [Google Scholar] [CrossRef] [Green Version]
- Benoit Bouvrette, L.P.; Bovaird, S.; Blanchette, M.; Lecuyer, E. oRNAment: A database of putative RNA binding protein target sites in the transcriptomes of model species. Nucleic Acids Res. 2020, 48, D166–D173. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Cheng, L.; Shi, H.; Zhang, Z.; Zhao, H.; Wang, Z.; Zhou, M. A potential panel of six-long non-coding RNA signature to improve survival prediction of diffuse large-B-cell lymphoma. Sci. Rep. 2016, 6, 27842. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Jiang, Y.; Du, W.; Fairchild, L.; Melnick, A.; Elemento, O. Transcriptome sequencing reveals thousands of novel long non-coding RNAs in B cell lymphoma. Genome Med. 2015, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Zhao, H.; Xu, W.; Bao, S.; Cheng, L.; Sun, J. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol. Cancer 2017, 16, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Representative long non-coding RNAs (lncRNAs) in diffuse large B-cell lymphoma (DLBCL). (A) The p53-activated PANDA inhibits cell growth through inactivation of the MAPK/ERK pathway. (B) The MYC-activated FIRRE activates the Wnt/β-catenin signaling pathway to facilitate DLBCL cell growth. (C) The lncRNA DBH-AS1 controls the translation of fibronectin 1 (FN1) through binding to the RNA-binding protein, BUD13. (D) Many lncRNAs have been suggested to function as miRNA sponges to sequester miR-195.
Figure 1.
Representative long non-coding RNAs (lncRNAs) in diffuse large B-cell lymphoma (DLBCL). (A) The p53-activated PANDA inhibits cell growth through inactivation of the MAPK/ERK pathway. (B) The MYC-activated FIRRE activates the Wnt/β-catenin signaling pathway to facilitate DLBCL cell growth. (C) The lncRNA DBH-AS1 controls the translation of fibronectin 1 (FN1) through binding to the RNA-binding protein, BUD13. (D) Many lncRNAs have been suggested to function as miRNA sponges to sequester miR-195.

Figure 2.
RNA-seq analysis of three rituximab-resistant and three rituximab-sensitive DLBCL cell lines. (A) Sample information for the DLBCL cell lines used in this study [95,96]. (B) Volcano plot of RNA-seq data. With the threshold values of 2-fold and p < 0.05, there are 195 up- and 428 down-regulated genes in rituximab-resistant compared to rituximab-sensitive DLCBL cell lines (n = 3 biologically independent samples). (C) Gene ontology (GO) of differentially expressed genes (both up- and down-regulated genes combined). (D) Hierarchical clustering of differentially expressed genes involved in GTPase signaling pathway.
Figure 2.
RNA-seq analysis of three rituximab-resistant and three rituximab-sensitive DLBCL cell lines. (A) Sample information for the DLBCL cell lines used in this study [95,96]. (B) Volcano plot of RNA-seq data. With the threshold values of 2-fold and p < 0.05, there are 195 up- and 428 down-regulated genes in rituximab-resistant compared to rituximab-sensitive DLCBL cell lines (n = 3 biologically independent samples). (C) Gene ontology (GO) of differentially expressed genes (both up- and down-regulated genes combined). (D) Hierarchical clustering of differentially expressed genes involved in GTPase signaling pathway.

Figure 3.
Molecular characteristics of differentially expressed lncRNAs in rituximab-resistant DLBCL cell lines compared to rituximab-sensitive cell lines. (A) Enrichment of transcription factor binding in the promoter regions of differentially expressed lncRNAs. The following 10 transcription factors with the highest number of binding sites in these promoter regions are shown in the image along with the log-transformed fold change (logFC) values in rituximab-resistant compared to rituximab-sensitive DLCBL cell lines: cut like homeobox 2 (CUX2); forkhead box N3 (FOXN3); HIC ZBTB transcriptional repressor 1 (HIC1); Kruppel like factor 5 (KLF5); myocyte enhancer factor 2C (MEF2C); Meis homeobox 1 (MEIS1); POU class 5 homeobox 1 (POU5F1); REL proto-oncogene, NF-kB subunit (REL); SRY-box transcription factor 5 (SOX5); and transcription factor 7 like 2 (TCF7L2). To allow for the visual inspection, 39 of 123 differentially expressed lncRNAs are shown along with the logFC values in rituximab-resistant compared to -sensitive DLCBL cell lines. (B) Protein-protein interactions (PPIs) of RNA-binding proteins (RBPs) predicted to bind CHROMR. PPIs are based on the information provided by the STRING database [99] visualized through NetworkAnalyst [100].
Figure 3.
Molecular characteristics of differentially expressed lncRNAs in rituximab-resistant DLBCL cell lines compared to rituximab-sensitive cell lines. (A) Enrichment of transcription factor binding in the promoter regions of differentially expressed lncRNAs. The following 10 transcription factors with the highest number of binding sites in these promoter regions are shown in the image along with the log-transformed fold change (logFC) values in rituximab-resistant compared to rituximab-sensitive DLCBL cell lines: cut like homeobox 2 (CUX2); forkhead box N3 (FOXN3); HIC ZBTB transcriptional repressor 1 (HIC1); Kruppel like factor 5 (KLF5); myocyte enhancer factor 2C (MEF2C); Meis homeobox 1 (MEIS1); POU class 5 homeobox 1 (POU5F1); REL proto-oncogene, NF-kB subunit (REL); SRY-box transcription factor 5 (SOX5); and transcription factor 7 like 2 (TCF7L2). To allow for the visual inspection, 39 of 123 differentially expressed lncRNAs are shown along with the logFC values in rituximab-resistant compared to -sensitive DLCBL cell lines. (B) Protein-protein interactions (PPIs) of RNA-binding proteins (RBPs) predicted to bind CHROMR. PPIs are based on the information provided by the STRING database [99] visualized through NetworkAnalyst [100].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Karstensen, K.T.; Schein, A.; Petri, A.; Bøgsted, M.; Dybkær, K.; Uchida, S.; Kauppinen, S. Long Non-Coding RNAs in Diffuse Large B-Cell Lymphoma. Non-Coding RNA 2021, 7, 1. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010001
AMA Style
Karstensen KT, Schein A, Petri A, Bøgsted M, Dybkær K, Uchida S, Kauppinen S. Long Non-Coding RNAs in Diffuse Large B-Cell Lymphoma. Non-Coding RNA. 2021; 7(1):1. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010001
Chicago/Turabian StyleKarstensen, Kasper Thystrup, Aleks Schein, Andreas Petri, Martin Bøgsted, Karen Dybkær, Shizuka Uchida, and Sakari Kauppinen. 2021. "Long Non-Coding RNAs in Diffuse Large B-Cell Lymphoma" Non-Coding RNA 7, no. 1: 1. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010001
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.