Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice

Abstract
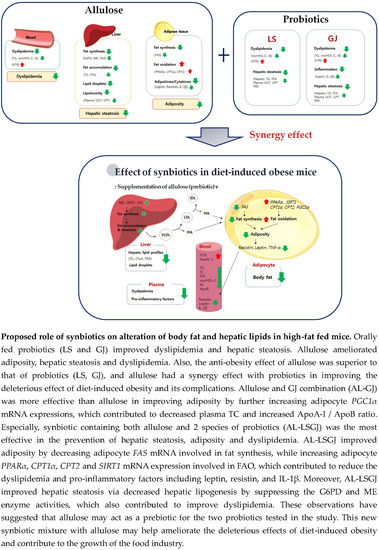
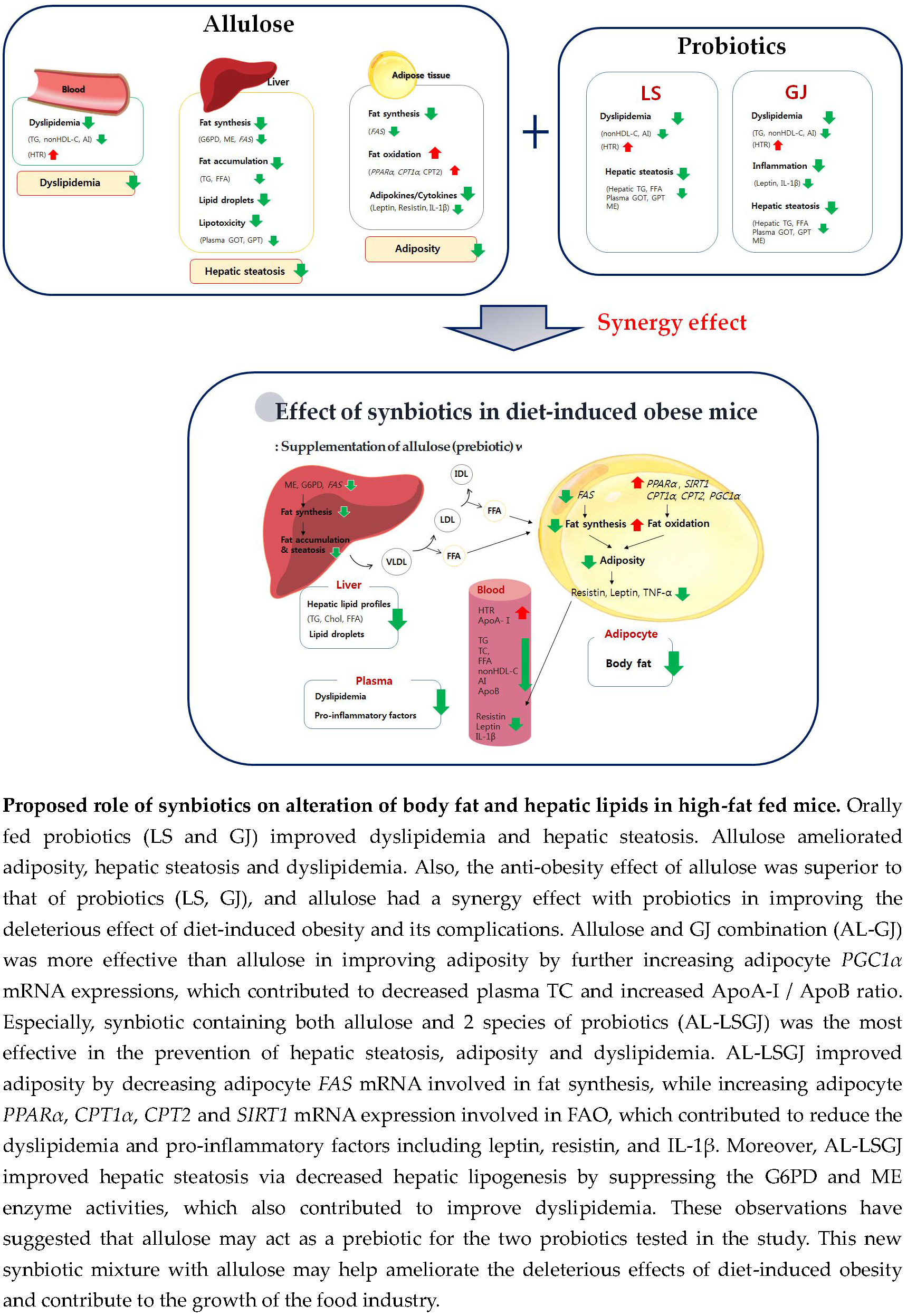
:
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Plasma Biomarkers
2.3. Hepatic Lipid Contents
2.4. Hepatic Lipid-Regulating Enzymes Activities
2.5. Real-Time qPCR Analysis
2.6. Histopathological Analysis
2.7. Statistical Analysis
3. Results
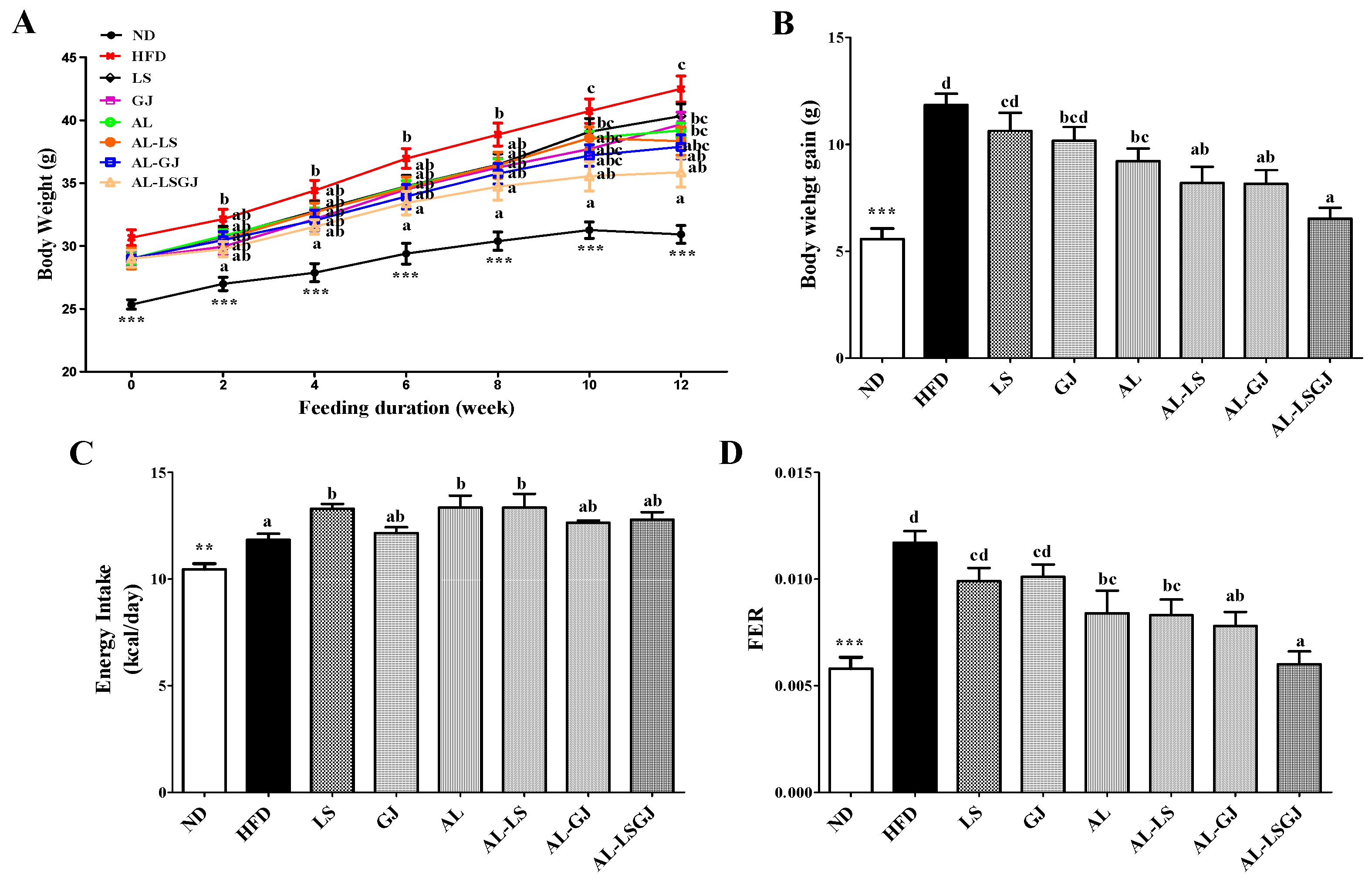
3.1. Body Weights and Food Efficiency Ratio
3.2. Adipose Tissue Weights
3.3. Plasma Lipid Profiles
3.4. Plasma Adipokine and Cytokine Levels
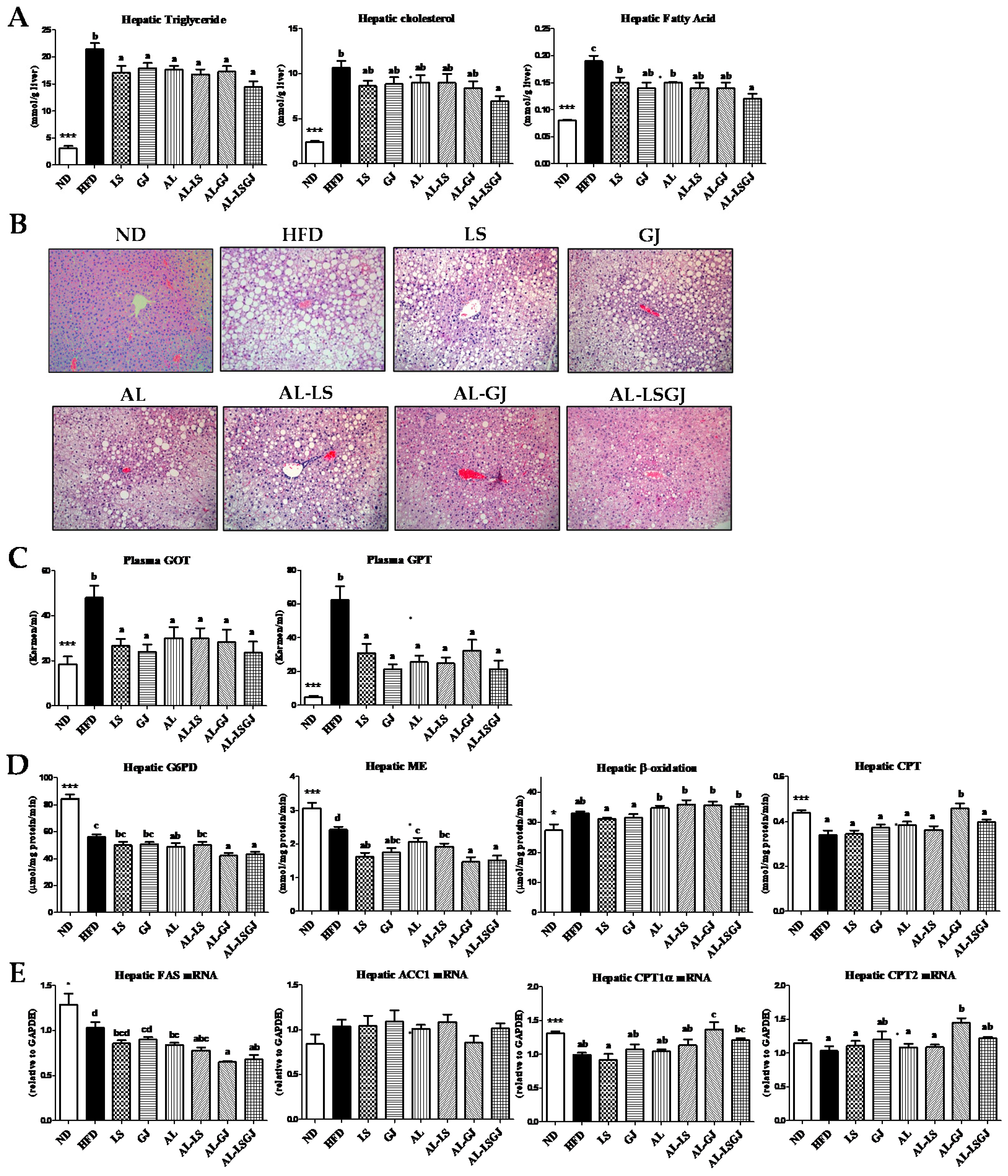
3.5. Hepatic Lipids Levels, Enzyme Activity, and mRNA Expression
3.6. mRNA Expression in Epididymal White Adipose Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sáez-Lara, M.J.; Robles-Sanchez, C.; Ruiz-Ojeda, F.J.; Plaza-Diaz, J.; Gil, A. Effects of Probiotics and Synbiotics on Obesity, Insulin Resistance Syndrome, Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease: A Review of Human Clinical Trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.; Moudgal, V. Probiotics: A review. JCOM 2012, 19, 76–84. [Google Scholar]
- Yoo, J.Y.; Kim, S.S. Probiotics and Prebiotics: Present Status and Future Perspectives on Metabolic Disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Umeki, M.; Oue, K.; Mochizuki, S.; Shirai, Y.; Sakai, K. Effect of Lactobacillus rhamnosus KY-3 and cellobiose as synbiotics on lipid metabolism in rats. J. Nutr. Sci. Vitaminol. 2004, 50, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Boutron-Ruault, M.C. Nutritional advantages of probiotics and prebiotics. Br. J. Nutr. 2002, 87, S153–S157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Singh, A.; Mathur, N. Review Synbiotics “Combinationof Probiotics and Prebiotics”. IOSR-JBB 2017, 3, 19–24. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of Leuconostoc mesenteroides starter cultures on microbial communities and metabolites during kimchi fermentation. Int. J. Food. Microbiol. 2012, 153, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.; Jeong, J.J.; Woo, K.H.; Han, M.J.; Kim, D.H. Lactobacillus sakei OK67 ameliorates high-fat diet-induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.Y.; Choi, E.A.; Lee, J.J.; Chang, H.C. Characterization of starter kimchi fermented with Leuconostoc kimchii GJ2 and its cholesterol-lowering effects in rats fed a high-fat and high-cholesterol diet. J. Sci. Food Agric. 2015, 95, 2750–2756. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Matsuo, T.; Tsukamoto, I.; Toyoda, Y.; Ogawa, M.; Nagata, Y.; Tokuda, M. Rare sugar d-allulose: Potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2015, 155, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Han, H.J.; Kim, A.H.; Choi, J.Y.; Cho, S.J.; Park, Y.B.; Jung, U.J.; Choi, M.S. d-Allulose supplementation normalized the body weight and fat-pad mass in diet-induced obese mice via the regulation of lipid metabolism under isocaloric fed condition. Mol. Nutr. Food Res. 2016, 60, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Kimoto-Nira, H.; Moriya, N.; Hayakawa, S.; Kuramasu, K.; Ohmori, H.; Yamasaki, S.; Ogawa, M. Effects of rare sugar d-allulose on acid production and probiotic activities of dairy lactic acid bacteria. J. Dairy Sci. 2017, 100, 5936–5944. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Hulcher, F.H.; Oleson, W.H. Simplified spectrophotometric assay for microsomal 3-hydroxy-3-methylglutaryl CoA reductase by measurement of coenzyme A. J. Lipid Res. 1973, 14, 625–631. [Google Scholar] [PubMed]
- Rudack, D.; Chisholm, E.M.; Holten, D. Rat liver glucose 6-phosphate dehydrogenase. Regulation by carbohydrate diet and insulin. J. Biol. Chem. 1971, 246, 1249–1254. [Google Scholar] [PubMed]
- Ochoa, S. Malic enzyme: Malic enzymes from pigeon and wheat germ. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; pp. 323–326. [Google Scholar]
- Markwell, M.A.; Mc Groarty, E.J.; Bieber, L.L.; Tolbert, N.E. The subcellular distribution of carnitine acyltransferases in mammalian liver and kidney. A new peroxisomal enzyme. J. Biol. Chem. 1973, 248, 3426–3432. [Google Scholar] [PubMed]
- Lazarow, P.B. Assay of peroxisomal beta-oxidation of fatty acids. Method Enzymol. 1981, 72, 315–319. [Google Scholar]
- Patterson, J.A.; Burkholder, K.M. Application of prebiotics and probiotics in poultry production. Poult. Sci. 2003, 82, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolida, S.; Gibson, G.R. Synbiotics in health and disease. Ann. Rev. Food Sci. Technol. 2011, 2, 373–393. [Google Scholar] [CrossRef] [PubMed]
- Rastall, R.A.; Maitin, V. Prebiotics and synbiotics: Towards the next generation. Curr. Opin. Biotechnol. 2002, 13, 490–496. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N.; Naruse, M.; Usui, T.; Tagami, T.; Suganami, T.; Yamada, K.; Kuzuya, H.; Shimatsu, A.; Ogawa, Y. Leptin-to-adiponectin ratio as a potential atherogenic index in obese type 2 diabetic patients. Diabetes Care 2004, 27, 2488–2490. [Google Scholar] [CrossRef] [PubMed]
- Vendrell, J.; Broch, M.; Vilarrasa, N.; Molina, A.; Gómez, J.M.; Gutiérrez, C.; Simón, I.; Soler, J.; Richart, C. Resistin, adiponectin, ghrelin, leptin, and proinflammatory cytokines: Relationships in obesity. Obes. Res. 2004, 12, 962–971. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, L.l.; Siersbæk, M.; Mandrup, S. PPARs: Fatty acid sensors controlling metabolism. Semin. Cell Dev. Biol. 2012, 23, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Gerhart-Hines, Z.; Rodgers, J.T.; Bare, O.; Lerin, C.; Kim, S.H.; Mostoslavsky, R.; Alt, F.W.; Wu, Z.; Puigserver, P. Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1alpha. EMBO J. 2007, 26, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Rottiers, V.; Näär, A.M. MicroRNAs in metabolism and metabolic disorders. Nat. Rev. Mol. Cell Biol. 2012, 13, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrisett, J.D.; Abdel-Fattah, G.; Hoogeveen, R.; Mitchell, E.; Ballantyne, C.M.; Pownall, H.J.; Opekun, A.R.; Jaffe, J.S.; Oppermann, S.; Kahan, B.D. Effects of sirolimus on plasma lipids, lipoprotein levels, and fatty acid metabolism in renal transplant patients. J. Lipid Res. 2002, 43, 1170–1180. [Google Scholar] [PubMed]
- Egusa, G.; Beltz, W.F.; Grundy, S.M.; Howard, B.V. Influence of obesity on the metabolism of apolipoprotein B in humans. J. Clin. Investig. 1985, 76, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | AL | |
|---|---|---|---|
| Casein | 20 | 20 | 20 |
| D, L-Methionine | 0.3 | 0.3 | 0.3 |
| Corn starch | 15 | 11.1 | 11.1 |
| Sucrose | 50 | 37 | 34 |
| Cellulose | 5 | 5 | 5 |
| Corn oil | 5 | 3 | 3 |
| Lard | 17 | 17 | |
| Mineral mix 1 | 3.5 | 4.2 | 4.2 |
| Vitamin mix 2 | 1 | 1.2 | 1.2 |
| Choline bitartrate | 0.2 | 0.2 | 0.2 |
| Cholesterol | 1 | 1 | |
| Tert-Butylhydroquinone | 0.001 | 0.004 | 0.004 |
| Allulose | 3 | ||
| Total (%) | 100 | 100 | 100 |
| Gene | Primer Direction | Primer Sequence |
|---|---|---|
| GAPDH | Forward | 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse | 5′-TGTAGACCATGTAGTTGAGGTCA-3′ | |
| FAS | Forward | 5′-GCTGCGGAAACTTCAGGAAAT-3′ |
| Reverse | 5′-AGAGACGTGTCACTCCTGGACTT-3′ | |
| ACC1 | Forward | 5′-GCCTCTTCCTGACAAACGAG-3′ |
| Reverse | 5′-TGACTGCCGAAACATCTCTG-3′ | |
| CPT1α | Forward | 5′-ATCTGGATGGCTATGGTCAAGGTC-3′ |
| Reverse | 5′-GTGCTGTCATGCGTTGGAAGTC-3′ | |
| CPT2 | Forward | 5′-GCCTGCTGTTGCGTGACTG-3′ |
| Reverse | 5′-TGGTGGGTACGATGCTGTGC-3′ | |
| PPARα | Forward | 5′-GGCACCCTCACATCATCAAACTG-3′ |
| Reverse | 5′-TGGAACAGACGGCGGCTTTC-3′ | |
| SIRT1 | Forward | 5′-TGTGAAGTTACTGCAGGAGTGTAAA-3′ |
| Reverse | 5′-GCATAGATACCGTCTCTTGATCTGAA-3′ | |
| PGC1α | Forward | 5′-AAGTGTGGAACTCTCTGGAACTG-3′ |
| Reverse | 5′-GGGTTATCTTGGTTGGCTTTATG-3′ |
| ND | HFD | LS | GJ | AL | AL-LS | AL-GJ | AL-LSGJ | |
|---|---|---|---|---|---|---|---|---|
| Liver | 3.27 ± 0.05 *** | 4.67 ± 0.20 | 4.18 ± 0.16 | 4.27 ± 0.25 | 4.56 ± 0.12 | 4.49 ± 0.10 | 4.31 ± 0.19 | 4.18 ± 0.11 |
| Kidney | 0.92 ± 0.02 *** | 0.72 ± 0.01 a | 0.76 ± 0.03 ab | 0.76 ± 0.03 ab | 0.83 ± 0.03 bc | 0.85 ± 0.02 c | 0.90 ± 0.03 c | 0.89 ± 0.05 c |
| Epididymal WAT | 3.11 ± 0.21 *** | 6.05 ± 0.11 c | 6.03 ± 0.27 c | 5.98 ± 0.12 bc | 5.89 ± 0.25 bc | 5.43 ± 0.18 abc | 5.18 ± 0.34 ab | 4.84 ± 0.25 a |
| Perirenal WAT | 0.40 ± 0.03 *** | 0.94 ± 0.04 d | 0.79 ± 0.07 bcd | 0.90 ± 0.11 cd | 0.71 ± 0.06 abc | 0.66 ± 0.05 ab | 0.57 ± 0.07 ab | 0.54 ± 0.04 a |
| Mesenteric WAT | 0.88 ± 0.07 *** | 1.97 ± 0.12 e | 1.77 ± 0.19 de | 1.60 ± 0.14 cd | 1.26 ± 0.13 bc | 1.19 ± 0.08 abc | 1.07 ± 0.10 ab | 0.86 ± 0.06 a |
| Subcutaneous WAT | 2.67 ± 0.24 *** | 6.10 ± 0.22 b | 5.59 ± 0.38 b | 5.65 ± 0.32 b | 5.68 ± 0.50 b | 5.17 ± 0.27ab | 4.37 ± 0.41 a | 4.35 ± 0.31 a |
| Retroperitoneal WAT | 0.75 ± 0.06 *** | 1.44 ± 0.03 b | 1.34 ± 0.05 ab | 1.28 ± 0.11 ab | 1.38 ± 0.07 ab | 1.30 ± 0.06 ab | 1.19 ± 0.09 a | 1.21 ± 0.08 a |
| Interscapular WAT | 1.53 ± 0.13 *** | 3.27 ± 0.08 c | 2.77 ± 0.20 b | 2.70 ± 0.13 b | 2.57 ± 0.24 b | 2.33 ± 0.14 ab | 2.02 ± 0.19 a | 2.03 ± 0.16 a |
| Total WAT | 9.34 ± 0.68 *** | 19.63 ± 0.40 d | 19.29 ± 0.65 d | 18.11 ± 0.64 cd | 17.49 ± 1.14 cd | 16.49 ± 0.44 bc | 14.69 ± 1.00 ab | 14.09 ± 0.63 a |
| Visceral WAT | 5.14 ± 0.35 *** | 10.39 ± 0.18 d | 9.93 ± 0.51 cd | 9.76 ± 0.22 cd | 9.23 ± 0.43 bc | 8.58 ± 0.32 ab | 8.01 ± 0.56 ab | 7.45 ± 0.36 a |
| ND | HFD | LS | GJ | AL | AL-LS | AL-GJ | AL-LSGJ | |
|---|---|---|---|---|---|---|---|---|
| TG (mmol/L) | 0.80 ± 0.05 ** | 0.98 ± 0.05 b | 0.87 ± 0.05 ab | 0.83 ± 0.05 a | 0.78 ± 0.05 a | 0.75 ± 0.06 a | 0.82 ± 0.03 a | 0.73 ± 0.03 a |
| Total-C (mmol/L) | 3.37 ± 0.14 *** | 5.29 ± 0.22 c | 4.62 ± 0.26 bc | 4.56 ± 0.18 bc | 4.81 ± 0.29 bc | 4.85 ± 0.26 bc | 4.32 ± 0.30 a | 4.32 ± 0.15 a |
| FFA (mmol/L) | 0.71 ± 0.01 * | 0.77 ± 0.02 b | 0.74 ± 0.02 ab | 0.72 ± 0.02 ab | 0.73 ± 0.02 ab | 0.71 ± 0.02 ab | 0.75 ± 0.02 ab | 0.69 ± 0.02 a |
| HDL-C (mmol/L) | 1.03 ± 0.06 *** | 1.62 ± 0.04 a | 1.64 ± 0.11 a | 1.76 ± 0.07 ab | 1.75 ± 0.09 ab | 1.71 ± 0.06 ab | 1.58 ± 0.08 a | 1.88 ± 0.06 b |
| nonHDL-C (mmol/L) | 2.45 ± 0.12 *** | 3.79 ± 0.19 b | 2.96 ± 0.17 a | 2.78 ± 0.16 a | 3.10 ± 0.21 a | 3.15 ± 0.14 a | 2.59 ± 0.20 a | 2.69 ± 0.13 a |
| HTR (%) | 30.63 ± 0.63 | 29.29 ± 0.73 a | 35.46 ± 0.64 b | 36.80 ± 1.17 b | 35.80 ± 1.15 b | 35.04 ± 1.51 b | 37.03 ± 1.03 b | 37.90 ± 1.07 b |
| AI | 2.24 ± 0.07 | 2.46 ± 0.10 b | 1.80 ± 0.04 a | 1.69 ± 0.10 a | 1.87 ± 0.09 a | 1.82 ± 0.11 a | 1.76 ± 0.08 a | 1.72 ± 0.07 a |
| ApoA-I (mg/dL) | 44.72 ± 0.43 | 44.87 ± 0.42 b | 43.39 ± 0.28 a | 44.51 ± 0.32 ab | 44.61 ± 0.75 ab | 43.91 ± 0.51 ab | 45.20 ± 0.42 b | 49.05 ± 0.60 c |
| ApoB 100 (mg/dL) | 2.74 ± 0.24 *** | 5.91 ± 0.61 b | 5.10 ± 0.38 ab | 4.62 ± 0.82 ab | 5.10 ± 0.25 ab | 5.07 ± 0.59 ab | 3.78 ± 0.35 a | 3.37 ± 0.37 a |
| ApoA-I/ApoB | 19.01 ± 0.97 *** | 8.56 ± 0.72 a | 8.85 ± 0.51 a | 11.97 ± 1.43 ab | 8.89 ± 0.39 a | 10.13 ± 1.43 ab | 13.15 ± 1.47 bc | 16.30 ± 2.09 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.-R.; Kwon, E.-Y.; Kim, H.-J.; Choi, M.-S. Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice. Nutrients 2018, 10, 1797. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111797
Choi B-R, Kwon E-Y, Kim H-J, Choi M-S. Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice. Nutrients. 2018; 10(11):1797. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111797
Chicago/Turabian StyleChoi, Bo-Ra, Eun-Young Kwon, Hye-Jin Kim, and Myung-Sook Choi. 2018. "Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice" Nutrients 10, no. 11: 1797. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111797