Molecular Fingerprints of Iron Parameters among a Population-Based Sample

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Laboratory Measurements and Phenotypic Characterization
2.3. Metabolomics Measurements
2.4. Statistical Analysis
3. Results
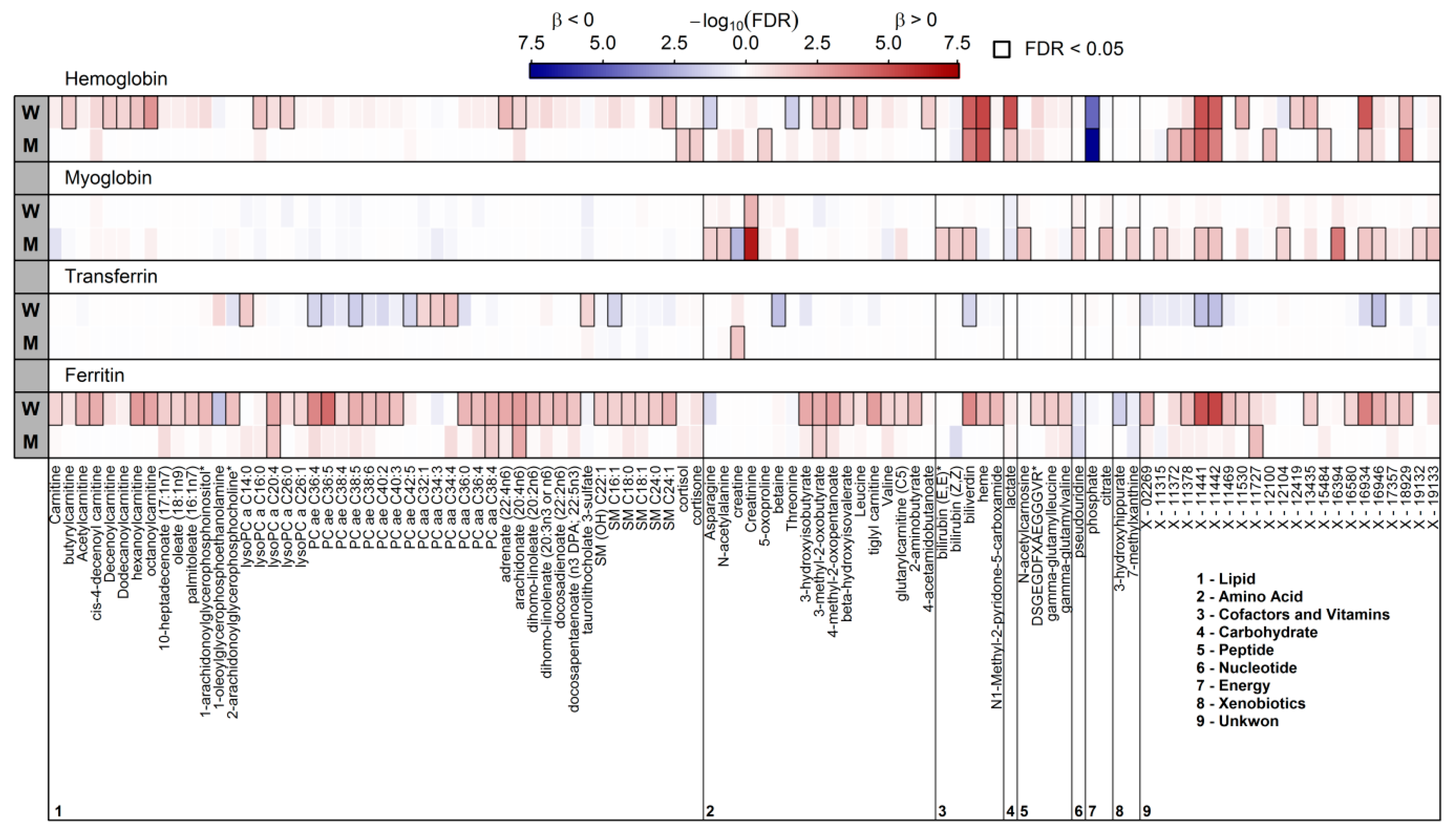
3.1. Surrogates of Iron Metabolism and the Plasma Metabolome
3.2. Significantly Associated Plasma Metabolites in Women
3.3. Significantly Associated Plasma Metabolites in Men
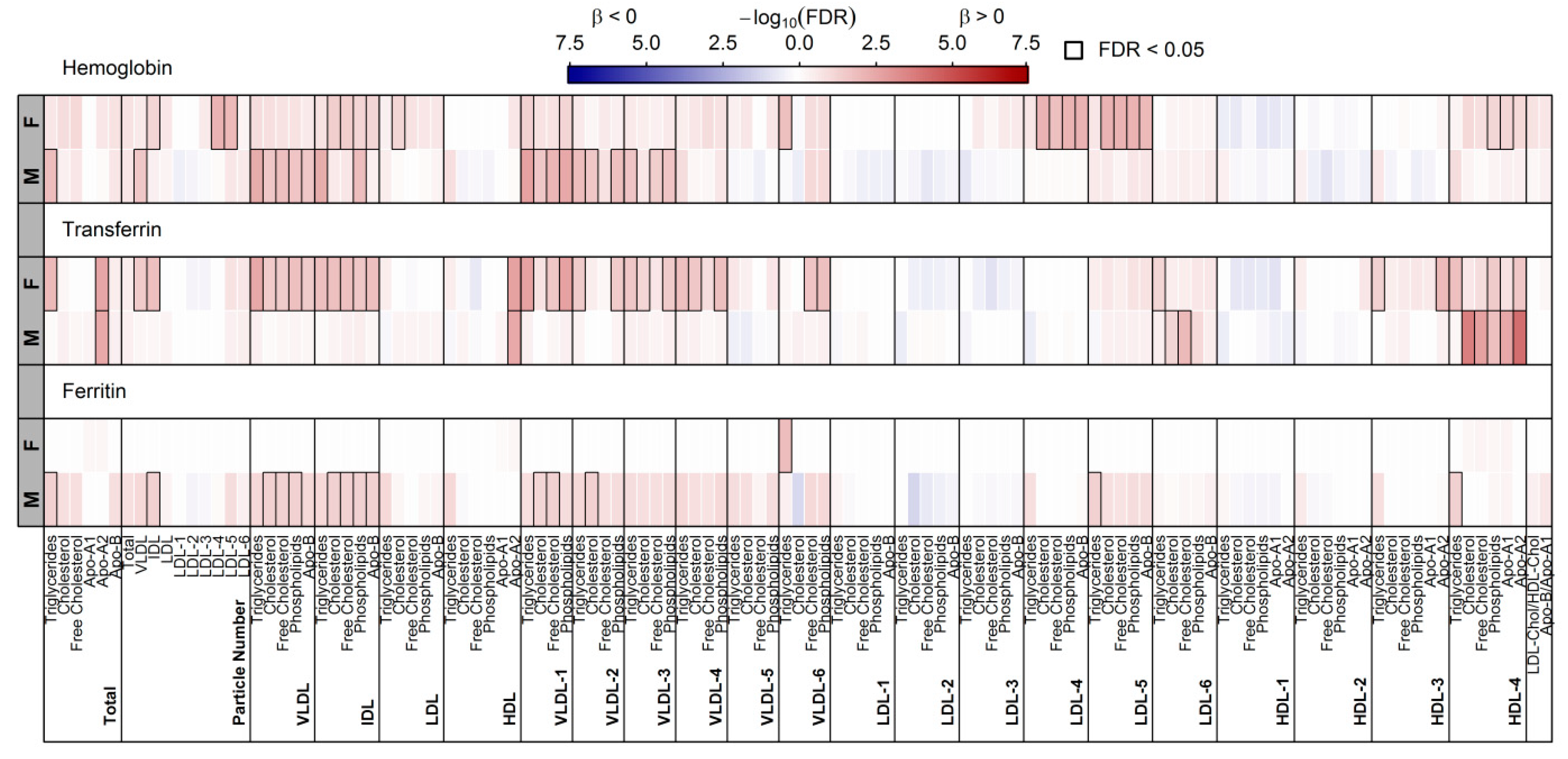
3.4. Surrogates of Iron Metabolism and Lipoprotein Particles
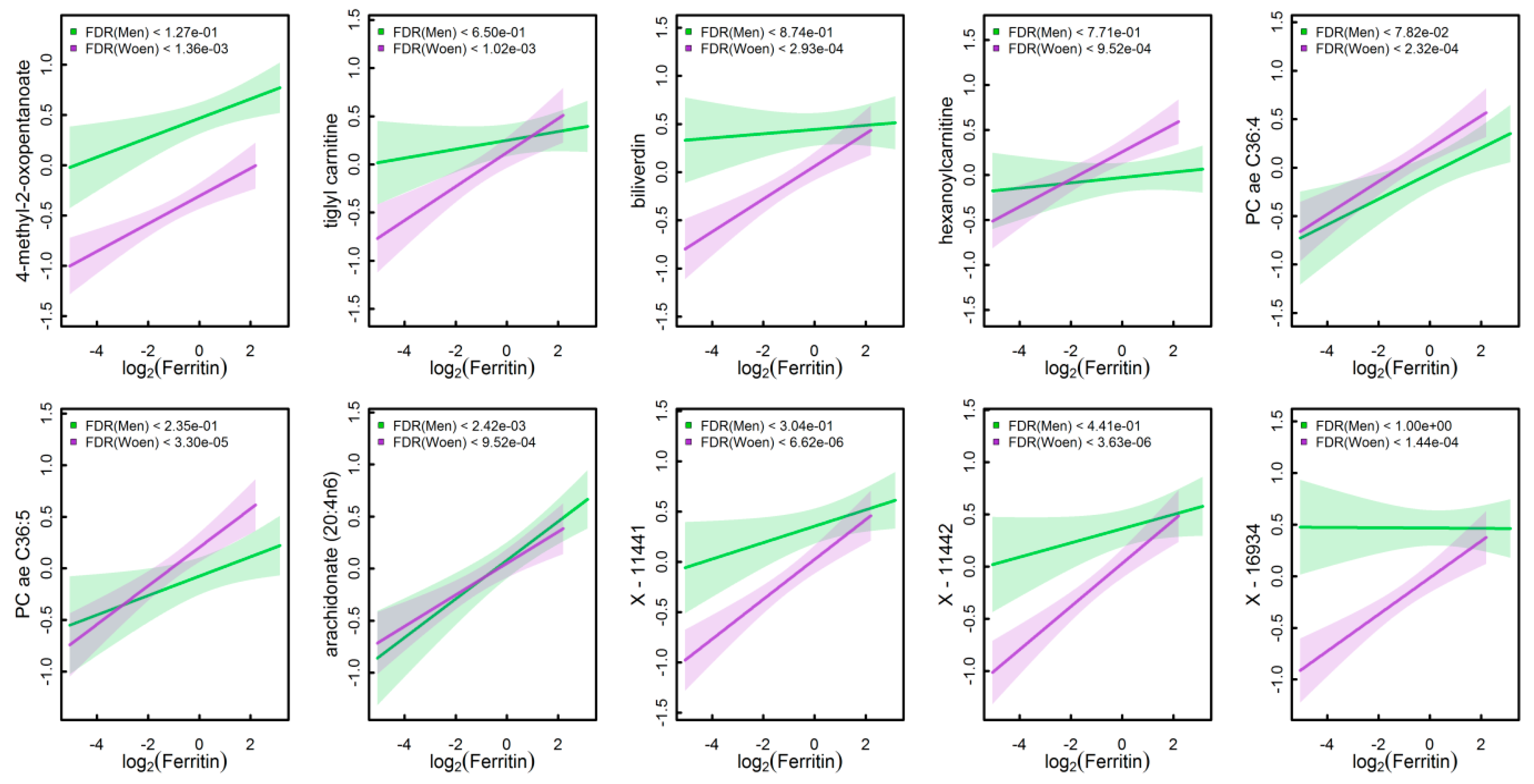
3.5. Identification of Markers to Differentiate High from Low Iron Status
4. Discussion
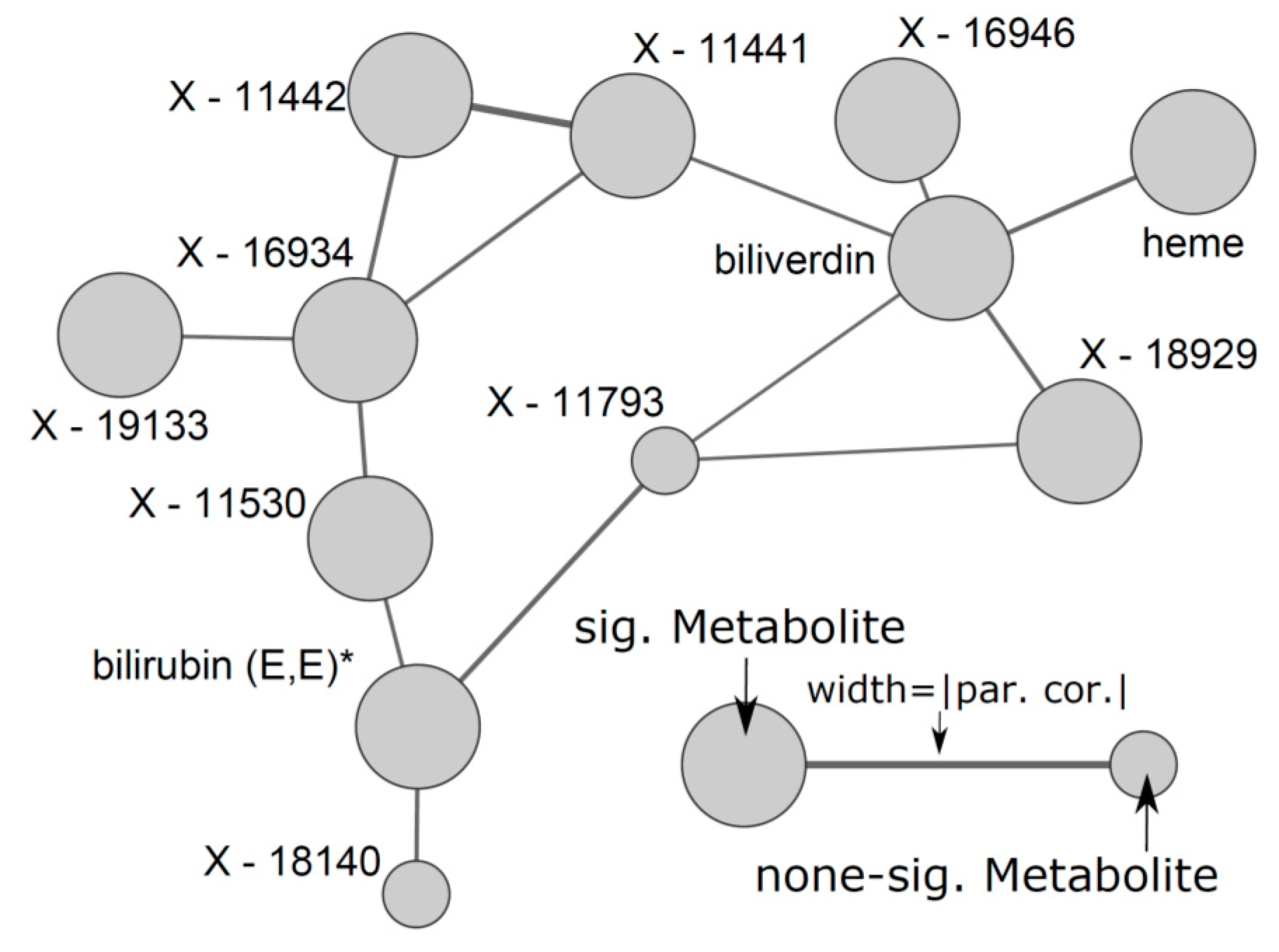
4.1. Overlapping Metabolites Reflect Heme Degradation
4.2. A Complex Metabolic Pattern is Associated with Serum Ferritin
4.3. Haemoglobin and Lipoprotein Metabolism in Males
4.4. Serum Transferrin and Lipoprotein Metabolism
4.5. Potential Markers to Predict Iron Depletion
4.6. A Short Note on the Strong Sex—Differences in Iron Metabolism
4.7. Myoglobin Reflects Male Muscle Metabolism
4.8. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Smith, R.S. Iron deficiency and iron overload. Arch. Dis. Child. 1965, 40, 343–363. [Google Scholar] [CrossRef] [PubMed]
- Trump, B.F.; Valigorsky, J.M.; Arstila, A.U.; Mergner, W.J.; Kinney, T.D. The relationship of intracellular pathways of iron metabolism to cellular iron overload and the iron storage diseases. Cell sap and cytocavitary network pathways in relation to lysosomal storage and turnover of iron macromolecules. Am. J. Pathol. 1973, 72, 295–336. [Google Scholar] [PubMed]
- Thorstensen, K.; Romslo, I. The role of transferrin in the mechanism of cellular iron uptake. Biochem. J. 1990, 271, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, P.; Kulkarni, H.; Dheda, S.; Betti, S.; Harrison, C.; St Pierre, T.G.; Olynyk, J.K. Serum iron markers are inadequate for guiding iron repletion in chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2011, 6, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Warner, M.J.; Kamran, M.T. Anemia, iron deficiency. In Statpearls; StatPearls: Treasure Island, FL, USA, 2018. [Google Scholar]
- Sahay, M.; Kalra, S.; Badani, R.; Bantwal, G.; Bhoraskar, A.; Das, A.K.; Dhorepatil, B.; Ghosh, S.; Jeloka, T.; Khandelwal, D.; et al. Diabetes and anaemia: International diabetes federation (idf)—southeast asian region (sear) position statement. Diabetes Metab. Syndr. 2017, 11 (Suppl. 2), S685–S695. [Google Scholar] [CrossRef] [PubMed]
- Vela, D.; Sopi, R.B.; Mladenov, M. Low hepcidin in type 2 diabetes mellitus: Examining the molecular links and their clinical implications. Can. J. Diabetes 2018, 42, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Aranda, N.; Fernandez-Cao, J.C.; Tous, M.; Arija, V. Increased iron levels and lipid peroxidation in a mediterranean population of Spain. Eur. J. Clin. Investig. 2016, 46, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Zaribaf, F.; Entezari, M.H.; Hassanzadeh, A.; Mirzaian, S. Association between dietary iron, iron stores and serum lipid profile in reproductive age women. J. Educ. Health Promot. 2014, 3, 15. [Google Scholar] [PubMed]
- Kraml, P. The role of iron in the pathogenesis of atherosclerosis. Physiol. Res. 2017, 66, S55–S67. [Google Scholar] [PubMed]
- Shipra, B.K.G.; Solanki, R.; Punia, H.; Agarwal, V.; Kaur, J.; Shukla, A. Relationship of lipid profile and serum ferritin levels with acute myocardial infarction. J. Clinical Diagn. Res. 2014, 8, CC10–CC13. [Google Scholar]
- Zhu, Y.N.; He, B.T.; Jing, J.; Ma, J.; Li, X.H.; Yang, W.H.; Jin, Y.; Chen, Y.J. Hepcidin and iron metabolism associated with cardiometabolic risk factors in children: A case-control study. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Britton, L.J.; Subramaniam, V.N.; Crawford, D.H. Iron and non-alcoholic fatty liver disease. World J. Gastroenterol. 2016, 22, 8112–8122. [Google Scholar] [CrossRef] [PubMed]
- Bictash, M.; Ebbels, T.M.; Chan, Q.; Loo, R.L.; Yap, I.K.; Brown, I.J.; de Iorio, M.; Daviglus, M.L.; Holmes, E.; Stamler, J.; et al. Opening up the “black box”: Metabolic phenotyping and metabolome-wide association studies in epidemiology. J. Clin. Epidemiol. 2010, 63, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Wittenbecher, C.; Muhlenbruch, K.; Kroger, J.; Jacobs, S.; Kuxhaus, O.; Floegel, A.; Fritsche, A.; Pischon, T.; Prehn, C.; Adamski, J.; et al. Amino acids, lipid metabolites and ferritin as potential mediators linking red meat consumption to type 2 diabetes. Am. J. Clin. Nutr. 2015, 101, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.H. Current information gaps in micronutrient research, programs and policy: How can we fill them? World Rev. Nutr. Diet. 2016, 115, 109–117. [Google Scholar] [PubMed]
- Volzke, H.; Alte, D.; Schmidt, C.O.; Radke, D.; Lorbeer, R.; Friedrich, N.; Aumann, N.; Lau, K.; Piontek, M.; Born, G.; et al. Cohort profile: The study of health in pomerania. Int. J. Epidemiol. 2011, 40, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Volzke, H.; Alte, D.; Schwahn, C.; Grabe, H.J.; Hoffmann, W.; John, U.; Doren, M. Menopause and determinants of quality of life in women at midlife and beyond: The study of health in pomerania (ship). Menopause 2007, 14, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Inker, L.A.; Schmid, C.H.; Tighiouart, H.; Eckfeldt, J.H.; Feldman, H.I.; Greene, T.; Kusek, J.W.; Manzi, J.; Van Lente, F.; Zhang, Y.L.; et al. Estimating glomerular filtration rate from serum creatinine and cystatin c. N. Engl. J. Med. 2012, 367, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Masuch, A.; Pietzner, M.; Bahls, M.; Budde, K.; Kastenmuller, G.; Zylla, S.; Artati, A.; Adamski, J.; Volzke, H.; Dorr, M.; et al. Metabolomic profiling implicates adiponectin as mediator of a favorable lipoprotein profile associated with NT-proBNP. Cardiovasc. Diabetol. 2018, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Pietzner, M.; Budde, K.; Homuth, G.; Kastenmuller, G.; Henning, A.K.; Artati, A.; Krumsiek, J.; Volzke, H.; Adamski, J.; Lerch, M.M.; et al. Hepatic steatosis is associated with adverse molecular signatures in subjects without diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 3856–3868. [Google Scholar] [CrossRef] [PubMed]
- Knacke, H.; Pietzner, M.; Do, K.T.; Romisch-Margl, W.; Kastenmuller, G.; Volker, U.; Volzke, H.; Krumsiek, J.; Artati, A.; Wallaschofski, H.; et al. Metabolic fingerprints of circulating igf-1 and the igf-1/igfbp-3 ratio: A multifluid metabolomics study. J. Clin. Endocrinol. Metab. 2016, 101, 4730–4742. [Google Scholar] [CrossRef] [PubMed]
- Pietzner, M.; Kaul, A.; Henning, A.K.; Kastenmuller, G.; Artati, A.; Lerch, M.M.; Adamski, J.; Nauck, M.; Friedrich, N. Comprehensive metabolic profiling of chronic low-grade inflammation among generally healthy individuals. BMC Med. 2017, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Non-esterified fatty acid metabolism and postprandial lipaemia. Atherosclerosis 1998, 141 (Suppl. 1), S41–S46. [Google Scholar] [CrossRef]
- Bhagavan, N.V. Lipids iii: Plasma lipoproteins. In Medical Biochemistry, 4th ed.; Harcourt/Academic Press: London, UK, 2002; pp. 439–450. [Google Scholar]
- Zhao, X.; Fritsche, J.; Wang, J.; Chen, J.; Rittig, K.; Schmitt-Kopplin, P.; Fritsche, A.; Haring, H.U.; Schleicher, E.D.; Xu, G.; et al. Metabonomic fingerprints of fasting plasma and spot urine reveal human pre-diabetic metabolic traits. Metabolomics 2010, 6, 362–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Ginsberg, H.N. Increased very low density lipoprotein (VLDL) secretion, hepatic steatosis and insulin resistance. Trends Endocrinol. Metab. 2011, 22, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Halle, M.; Konig, D.; Berg, A.; Keul, J.; Baumstark, M.W. Relationship of serum ferritin concentrations with metabolic cardiovascular risk factors in men without evidence for coronary artery disease. Atherosclerosis 1997, 128, 235–240. [Google Scholar] [CrossRef]
- Upadhyay, R.K. Emerging risk biomarkers in cardiovascular diseases and disorders. J. Lipids 2015, 2015, 971453. [Google Scholar] [CrossRef] [PubMed]
- Asghari, G.; Farhadnejad, H.; Teymoori, F.; Mirmiran, P.; Tohidi, M.; Azizi, F. High dietary intake of branched-chain amino acids is associated with an increased risk of insulin resistance in adults. J. Diabetes 2018, 10, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Xie, G.; Jia, W.; Jia, W. Insulin resistance and the metabolism of branched-chain amino acids. Front. Med. 2013, 7, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Honda, T.; Shiraki, M.; Murakami, T.; Sato, J.; Kobayashi, H.; Mawatari, K.; Obayashi, M.; Harris, R.A. Branched-chain amino acid catabolism in exercise and liver disease. J. Nutr. 2006, 136, 250s–253s. [Google Scholar] [CrossRef] [PubMed]
- Stechemesser, L.; Eder, S.K.; Wagner, A.; Patsch, W.; Feldman, A.; Strasser, M.; Auer, S.; Niederseer, D.; Huber-Schonauer, U.; Paulweber, B.; et al. Metabolomic profiling identifies potential pathways involved in the interaction of iron homeostasis with glucose metabolism. Mol. Metab. 2017, 6, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, G. Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, R.M.; Labad, J.; Strachan, M.W.; Braun, A.; Fowkes, F.G.; Lee, A.J.; Frier, B.M.; Seckl, J.R.; Walker, B.R.; Price, J.F. Elevated fasting plasma cortisol is associated with ischemic heart disease and its risk factors in people with type 2 diabetes: The edinburgh type 2 diabetes study. J. Clin. Endocrinol. Metab. 2010, 95, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.J.; Herzig, S. Metabolic control through glucocorticoid hormones: An update. Mol. Cell. Endocrinol. 2013, 380, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltmanns, K.M.; Dodt, B.; Schultes, B.; Raspe, H.H.; Schweiger, U.; Born, J.; Fehm, H.L.; Peters, A. Cortisol correlates with metabolic disturbances in a population study of type 2 diabetic patients. Eur. J. Endocrinol. 2006, 154, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staufenbiel, S.M.; Penninx, B.W.; Spijker, A.T.; Elzinga, B.M.; van Rossum, E.F. Hair cortisol, stress exposure and mental health in humans: A systematic review. Psychoneuroendocrinology 2013, 38, 1220–1235. [Google Scholar] [CrossRef] [PubMed]
- Dugue, B.; Leppanen, E.; Grasbeck, R. The driving license examination as a stress model: Effects on blood picture, serum cortisol and the production of interleukins in man. Life Sci. 2001, 68, 1641–1647. [Google Scholar] [PubMed]
- Vaisar, T.; Pennathur, S.; Green, P.S.; Gharib, S.A.; Hoofnagle, A.N.; Cheung, M.C.; Byun, J.; Vuletic, S.; Kassim, S.; Singh, P.; et al. Shotgun proteomics implicates protease inhibition and complement activation in the antiinflammatory properties of HDL. J. Clin. Investig. 2007, 117, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherson, P.A.; Young, I.S.; McKibben, B.; McEneny, J. High density lipoprotein subfractions: Isolation, composition and their duplicitous role in oxidation. J. Lipid Res. 2007, 48, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Antappanavar, V.B.; Biradar, S.G.; Patil, V.; Biradar, P.M.; Mithare, S.; Sharma, A.K. A study of correlation between iron deficiency anaemia and serum lipid profile in indian adults in brims, bidar. Int. J. Adv. Med. 2014, 1, 96–100. [Google Scholar] [CrossRef]
- Verma, U.; Shankar, N.; Madhu, S.V.; Tandon, O.P.; Madan, N.; Verma, N. Relationship between iron deficiency anaemia and serum lipid levels in indian adults. J. Indian Med. Assoc. 2010, 108, 555–558. [Google Scholar] [PubMed]
- Soga, T.; Sugimoto, M.; Honma, M.; Mori, M.; Igarashi, K.; Kashikura, K.; Ikeda, S.; Hirayama, A.; Yamamoto, T.; Yoshida, H.; et al. Serum metabolomics reveals gamma-glutamyl dipeptides as biomarkers for discrimination among different forms of liver disease. J. Hepatol. 2011, 55, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.D.; Finch, C.A. Iron nutrition. West. J. Med. 1975, 122, 474–481. [Google Scholar] [PubMed]
- Rushton, D.H.; Barth, J.H. What is the evidence for gender differences in ferritin and haemoglobin? Crit. Rev. Oncol. Hematol. 2010, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.M.; Whalley, P.J.; Pritchard, J.A. Measurements of menstrual blood loss. Am. J. Obstet. Gynecol. 1961, 81, 739–742. [Google Scholar] [CrossRef]
- Qamar, K.; Saboor, M.; Qudsia, F.; Khosa, S.M.; Moinuddin; Usman, M. Malabsorption of iron as a cause of iron deficiency anaemia in postmenopausal women. Pak. J. Med. Sci. 2015, 31, 304–308. [Google Scholar] [PubMed]
- Chen, B.; Li, G.F.; Shen, Y.; Huang, X.I.; Xu, Y.J. Reducing iron accumulation: A potential approach for the prevention and treatment of postmenopausal osteoporosis. Exp. Ther. Med. 2015, 10, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, J.; Pelle, E.; Huang, X. Iron and menopause: Does increased iron affect the health of postmenopausal women? Antioxid. Redox Signal. 2009, 11, 2939–2943. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, E.M.; Yurgalevitch, S.M.; Saritelli, A.L.; Herbert, P.N.; Thompson, P.D. Variations in plasma volume affect total and low-density lipoprotein cholesterol concentrations during the menstrual cycle. Metab. Clin. Exp. 1995, 44, 965–971. [Google Scholar] [CrossRef]
- Mumford, S.L.; Dasharathy, S.; Pollack, A.Z.; Schisterman, E.F. Variations in lipid levels according to menstrual cycle phase: Clinical implications. Clin. Lipidol. 2011, 6, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Vashishta, S.; Gahlot, S.; Goyal, R. Effect of menstrual cycle phases on plasma lipid and lipoprotein levels in regularly menstruating women. J. Clin. Diagn. Res. 2017, 11, CC05–CC07. [Google Scholar] [CrossRef] [PubMed]
- Sekula, P.; Dettmer, K.; Vogl, F.C.; Gronwald, W.; Ellmann, L.; Mohney, R.P.; Eckardt, K.U.; Suhre, K.; Kastenmuller, G.; Oefner, P.J.; et al. From discovery to translation: Characterization of C-Mannosyltryptophan and pseudouridine as markers of kidney function. Sci. Rep. 2017, 7, 17400. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Male (N = 409) | Female (N = 411) | p * |
|---|---|---|---|
| Age, years | 50 (40; 61) | 54 (44; 62) | 0.93 |
| Smoking, % | <0.01 | ||
| Never | 30.6 | 51.8 | |
| Former | 45.9 | 28.7 | |
| Current | 23.5 | 19.5 | |
| Waist Circumference, cm | 94 (87; 102) | 83 (76; 91) | <0.01 |
| Ferritin, µg/L | 149 (86; 255) | 60 (28; 104) | <0.01 |
| Transferrin, g/L | 2.5 (2.2; 2.7) | 2.5 (2.3; 2.8) | <0.01 |
| Myoglobin, µg/L | 58 (50; 71) | 44 (37; 54) | <0.01 |
| Haemoglobin, mmol/L | 9.1 (8.8; 9.5) | 8.3 (7.9; 8.6) | <0.01 |
| eGFR, mL/min/1.73 m² | 116 (108; 126) | 110 (101; 118) | <0.01 |
| ALT, µkatal/L | 0.47 (0.35; 0.65) | 0.31 (0.25; 0.44) | <0.01 |
| hsCRP, mg/L | 0.99 (0.56; 1,83) | 1.16 (0.62; 2.45) | <0.01 |
| Fibrinogen, g/L | 2.8 (2.3; 3.3) | 3.1 (2.6; 3.5) | <0.01 |
| Glucose, mmol/L | 5.4 (5.1;5.8) | 5.2 (4.9; 5.6) | <0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaul, A.; Masuch, A.; Budde, K.; Kastenmüller, G.; Artati, A.; Adamski, J.; Völzke, H.; Nauck, M.; Friedrich, N.; Pietzner, M. Molecular Fingerprints of Iron Parameters among a Population-Based Sample. Nutrients 2018, 10, 1800. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111800
Kaul A, Masuch A, Budde K, Kastenmüller G, Artati A, Adamski J, Völzke H, Nauck M, Friedrich N, Pietzner M. Molecular Fingerprints of Iron Parameters among a Population-Based Sample. Nutrients. 2018; 10(11):1800. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111800
Chicago/Turabian StyleKaul, Anne, Annette Masuch, Kathrin Budde, Gabi Kastenmüller, Anna Artati, Jerzy Adamski, Henry Völzke, Matthias Nauck, Nele Friedrich, and Maik Pietzner. 2018. "Molecular Fingerprints of Iron Parameters among a Population-Based Sample" Nutrients 10, no. 11: 1800. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111800