Expression of Selenoprotein Genes and Association with Selenium Status in Colorectal Adenoma and Colorectal Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Characteristics
2.2. Isolation of Total RNA
2.3. Synthesis of Complementary DNA (cDNA)
2.4. Relative Quantification of Gene Expression
2.5. Serum Selenium and Selenoprotein P Measurements
2.6. Statistical Analyses
3. Results
3.1. Selenoprotein Gene Transcript Levels in the Colorectal Adenomas
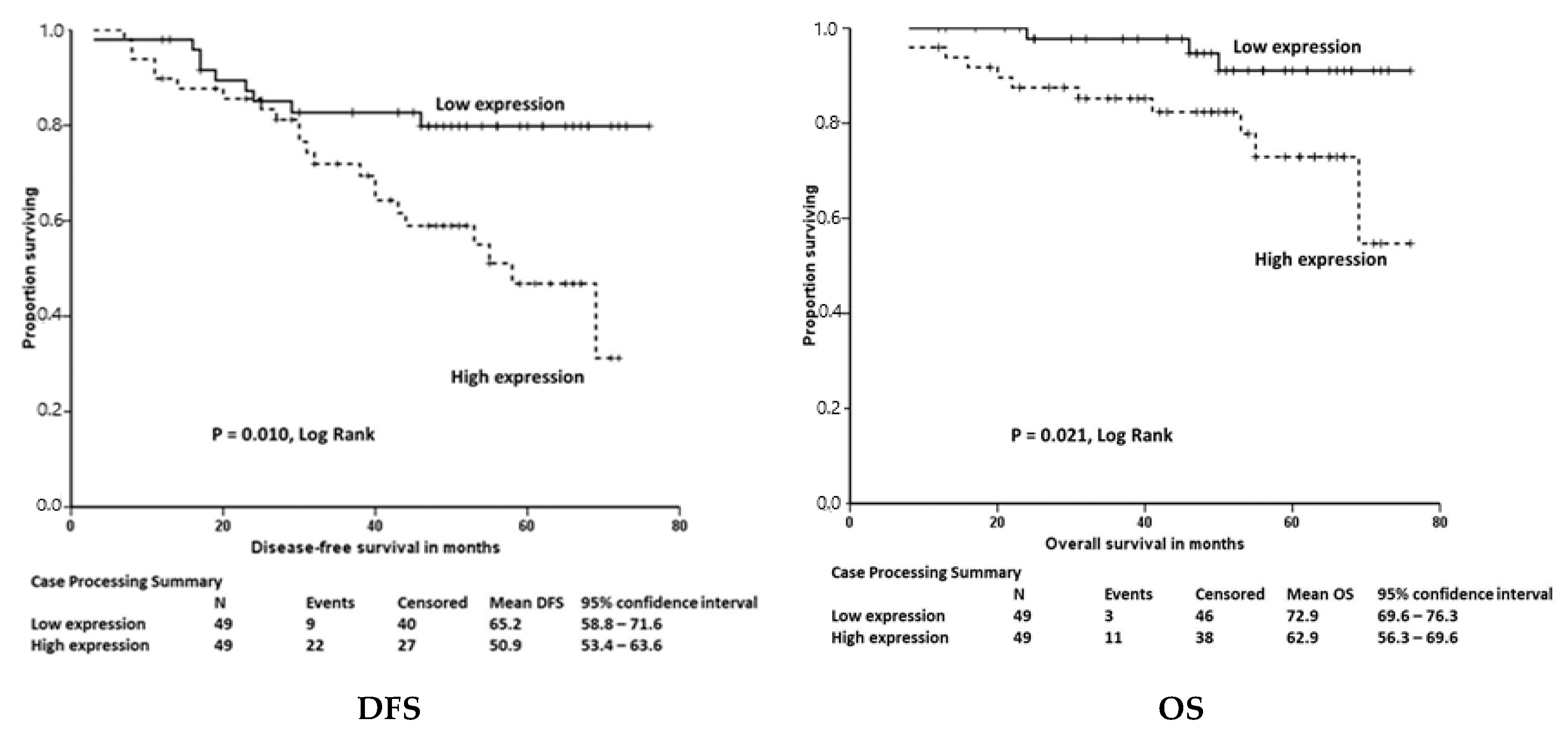
3.2. Selenoprotein Gene Transcript Levels in Colorectal Cancers
3.3. Correlation of the Selenoprotein Transcript Levels with Se Status
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Cappellani, A.; Zanghi, A.; Di Vita, M.; Cavallaro, A.; Piccolo, G.; Veroux, P.; Lo Menzo, E.; Cavallaro, V.; de Paoli, P.; Veroux, M.; et al. Strong correlation between diet and development of colorectal cancer. Front. Biosci. 2013, 18, 190–198. [Google Scholar]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, D.L.; Tsuji, P.A.; Carlson, B.A.; Gladyshev, V.N. Selenium and selenocysteine: Roles in cancer, health, and development. Trends Biochem. Sci. 2014, 39, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Speckmann, B.; Klotz, L.O. Selenoproteins: Antioxidant selenoenzymes and beyond. Arch. Biochem. Biophys. 2016, 595, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Meplan, C. Selenium and chronic diseases: A nutritional genomics perspective. Nutrients 2015, 7, 3621–3651. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Fedirko, V.; Jenab, M.; Schomburg, L.; Meplan, C.; Freisling, H.; Bueno-de-Mesquita, H.B.; Hybsier, S.; Becker, N.P.; Czuban, M.; et al. Selenium status is associated with colorectal cancer risk in the European prospective investigation of cancer and nutrition cohort. Int. J. Cancer 2015, 136, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Meplan, C.; Hesketh, J. Selenium and cancer: A story that should not be forgotten-insights from genomics. Adv. Nutr. Cancer 2014, 159, 145–166. [Google Scholar]
- Meplan, C.; Johnson, I.T.; Polley, A.C.; Cockell, S.; Bradburn, D.M.; Commane, D.M.; Arasaradnam, R.P.; Mulholland, F.; Zupanic, A.; Mathers, J.C.; et al. Transcriptomics and proteomics show that selenium affects inflammation, cytoskeleton, and cancer pathways in human rectal biopsies. FASEB J. 2016, 30, 2812–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, K.M.; Carlson, B.A.; Gladyshev, V.N.; Tsuji, P.A. Selenoproteins in colon cancer. Free Radic. Biol. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Schweizer, U. Hierarchical regulation of selenoprotein expression and sex-specific effects of selenium. Biochim. Biophys. Acta 2009, 1790, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Bertz, M.; Kuhn, K.; Koeberle, S.C.; Muller, M.F.; Hoelzer, D.; Thies, K.; Deubel, S.; Thierbach, R.; Kipp, A.P. Selenoprotein H controls cell cycle progression and proliferation of human colorectal cancer cells. Free Radic. Biol. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Al-Taie, O.H.; Uceyler, N.; Eubner, U.; Jakob, F.; Mork, H.; Scheurlen, M.; Brigelius-Flohe, R.; Schottker, K.; Abel, J.; Thalheimer, A.; et al. Expression profiling and genetic alterations of the selenoproteins GI-GPx and SePP in colorectal carcinogenesis. Nutr. Cancer 2004, 48, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Yagublu, V.; Arthur, J.R.; Babayeva, S.N.; Nicol, F.; Post, S.; Keese, M. Expression of selenium-containing proteins in human colon carcinoma tissue. Anticancer Res. 2011, 31, 2693–2698. [Google Scholar] [PubMed]
- Murawaki, Y.; Tsuchiya, H.; Kanbe, T.; Harada, K.; Yashima, K.; Nozaka, K.; Tanida, O.; Kohno, M.; Mukoyama, T.; Nishimuki, E.; et al. Aberrant expression of selenoproteins in the progression of colorectal cancer. Cancer Lett. 2008, 259, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Meplan, C.; Hughes, D.J.; Pardini, B.; Naccarati, A.; Soucek, P.; Vodickova, L.; Hlavata, I.; Vrana, D.; Vodicka, P.; Hesketh, J.E. Genetic variants in selenoprotein genes increase risk of colorectal cancer. Carcinogenesis 2010, 31, 1074–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reszka, E.; Jablonska, E.; Gromadzinska, J.; Wasowicz, W. Relevance of selenoprotein transcripts for selenium status in humans. Genes Nutr. 2012, 7, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M.; et al. Fusobacterium nucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1381–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winawer, S.J.; Zauber, A.G. The advanced adenoma as the primary target of screening. Gastrointest. Endosc. Clin. N. Am. 2002, 12, 1–9. [Google Scholar] [CrossRef]
- Hlavata, I.; Mohelnikova-Duchonova, B.; Vaclavikova, R.; Liska, V.; Pitule, P.; Novak, P.; Bruha, J.; Vycital, O.; Holubec, L.; Treska, V.; et al. The role of ABC transporters in progression and clinical outcome of colorectal cancer. Mutagenesis 2012, 27, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brynychova, V.; Hlavac, V.; Ehrlichova, M.; Vaclavikova, R.; Pecha, V.; Trnkova, M.; Wald, M.; Mrhalova, M.; Kubackova, K.; Pikus, T.; et al. Importance of transcript levels of caspase-2 isoforms s and l for breast carcinoma progression. Future Oncol. 2013, 9, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Soucek, P.; Anzenbacher, P.; Skoumalova, I.; Dvorak, M. Expression of cytochrome p450 genes in CD34+ hematopoietic stem and progenitor cells. Stem Cells 2005, 23, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A. Why the need for qPCR publication guidelines?—The case for MIQE. Methods 2010, 50, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Duarte-Salles, T.; Hybsier, S.; Trichopoulou, A.; Stepien, M.; Aleksandrova, K.; Overvad, K.; Tjonneland, A.; Olsen, A.; Affret, A.; et al. Prediagnostic selenium status and hepatobiliary cancer risk in the European prospective investigation into cancer and nutrition cohort. Am. J. Clin. Nutr. 2016, 104, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Meplan, C.; Hesketh, J. The influence of selenium and selenoprotein gene variants on colorectal cancer risk. Mutagenesis 2012, 27, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipp, A.; Banning, A.; van Schothorst, E.M.; Meplan, C.; Schomburg, L.; Evelo, C.; Coort, S.; Gaj, S.; Keijer, J.; Hesketh, J.; et al. Four selenoproteins, protein biosynthesis, and Wnt signalling are particularly sensitive to limited selenium intake in mouse colon. Molecular Nutr. Food Res. 2009, 53, 1561–1572. [Google Scholar] [CrossRef] [PubMed]
- Speckmann, B.; Steinbrenner, H. Selenium and selenoproteins in inflammatory bowel diseases and experimental colitis. Inflamm. Bowel Dis. 2014, 20, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, J.; Meplan, C. Transcriptomics and functional genetic polymorphisms as biomarkers of micronutrient function: Focus on selenium as an exemplar. Proc. Nutr. Soc. 2011, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Liu, M.; Ni, J.; Tian, J. Role of selenoprotein F in protein folding and secretion: Potential involvement in human disease. Nutrients 2018, 10, 1619. [Google Scholar] [CrossRef] [PubMed]
- Irons, R.; Tsuji, P.A.; Carlson, B.A.; Ouyang, P.; Yoo, M.H.; Xu, X.M.; Hatfield, D.L.; Gladyshev, V.N.; Davis, C.D. Deficiency in the 15-kDa selenoprotein inhibits tumorigenicity and metastasis of colon cancer cells. Cancer Prev. Res. 2010, 3, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, P.A.; Carlson, B.A.; Naranjo-Suarez, S.; Yoo, M.H.; Xu, X.M.; Fomenko, D.E.; Gladyshev, V.N.; Hatfield, D.L.; Davis, C.D. Knockout of the 15 kDa selenoprotein protects against chemically-induced aberrant crypt formation in mice. PLoS ONE 2012, 7, e50574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mork, H.; Al-Taie, O.H.; Bahr, K.; Zierer, A.; Beck, C.; Scheurlen, M.; Jakob, F.; Kohrle, J. Inverse mRNA expression of the selenocysteine-containing proteins GI-GPX and SePP in colorectal adenomas compared with adjacent normal mucosa. Nutr. Cancer 2000, 37, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Novoselov, S.V.; Kryukov, G.V.; Xu, X.M.; Carlson, B.A.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein H is a nucleolar thioredoxin-like protein with a unique expression pattern. J. Biol. Chem. 2007, 282, 11960–11968. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, Y.; Yang, X.; Jiang, Y. Selenium-binding protein 1 is associated with the degree of colorectal cancer differentiation and is regulated by histone modification. Oncol. Rep. 2014, 31, 2506–2514. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Fedirko, V.; Jones, J.S.; Méplan, C.; Schomburg, L.; Hybsier, S.; Riboli, E.; Hesketh, J.; Jenab, M. Association of selenoprotein and selenium pathway genetic variations with colorectal cancer risk and interaction with selenium status. In Global Advances in Selenium Research from Theory to Application, 1st ed.; Banuelos, G.S., Lin, Z.Q., Moraes, M.F., Guilherme, L.R.G., dos Reis, A.R., Eds.; Taylor & Francis: London, UK, 2015; pp. 53–54. ISBN 9781138027312. [Google Scholar]
- Click, B.; Pinsky, P.F.; Hickey, T.; Doroudi, M.; Schoen, R.E. Association of colonoscopy adenoma findings with long-term colorectal cancer incidence. JAMA 2018, 319, 2021–2031. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.C.; Fordyce, F.M.; Rayman, M.P. Symposium on ‘Geographical and geological influences on nutrition’: Factors controlling the distribution of selenium in the environment and their impact on health and nutrition. Proc. Nutr. Soc. 2010, 69, 119–132. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Cohort | Irish | Irish | Czech |
|---|---|---|---|
| Diagnosis | CRA | CRC | CRC |
| Total n of tissue samples | 40 | 22 | 105 |
| Sex n (male/female) | 26/14 | 11/11 | 64/25 |
| Age at diagnosis, median ± SD (years) | 61 ± 7 years | 59 ± 11 years | 69 ± 11 years |
| Location n (colon/rectum) | 28/12 | 15/7 | 82/17 |
| T staging n (/T1/T2/T3/T4) | - | 6/4/9/3 | - |
| N staging n (N0/N1 or N2) (missing) | - | 19/3 | 78/25 (2) |
| M staging n (M0/M1) | - | 21/1 | - |
| Stage (I/II/III/IV (missing)) | - | 10/7/4/1 | 2/76/15/10 (2) |
| Gene Name | Discovery (Irish Cohort, n = 62; 40 CRA and 22 CRC) | Validation (Czech Cohort, N = 105 CRC) Carcinoma Expression (p-Value) | TCGA (COAD/READ) Carcinoma Expression | |
|---|---|---|---|---|
| Adenoma Expression (p-Value) | Carcinoma Expression (p-Value) | |||
| GPX1 | NS | NS | ↑ (<0.001) | NS/NS |
| GPX2 | ↑ (<0.001) | ↑ (0.023) | ↑ (<0.001) | ↑/↑ |
| GPX3 | ↓ (<0.001) | ↓ (<0.001) | ↓ (<0.001) | ↓/↓ |
| GPX4 | NS | ↑ (0.039) | NS | NS/NS |
| SELENOF | NS | NS | NS | NS/NS |
| SELENOH | ↑ (<0.001) | NS | ↑ (<0.001) | NS/NS |
| SELENOK | NS | ↓ (0.015) | ↓ (<0.001) | NS/NS |
| SELENON | NS | NS | ↓ (<0.001) | NS/NS |
| SELENOP | ↓ (<0.001) | ↓ (0.001) | ↓ (<0.001) | ↓/↓ |
| SELENOS | ↓ (<0.001) | ↓ (<0.001) | ↓ (<0.001) | NS/NS |
| SELENOW | NS | NS | NS | NS/NS |
| SEPHS2 | ↓ (<0.001) | ↓ (0.002) | ↓ (<0.001) | NS/NS |
| TXNRD1 | NS | NS | NS | NS/NS |
| TXNRD2 | NS | NS | NS | NS/NS |
| TXNRD3 | ↑ (<0.001) | ↑ (0.003) | ↑ (<0.001) | ↑/↑ |
| SELENBP1 | NS | ↓ (<0.001) | ↓ (<0.001) | ↑/↑ |
| SOD2 | NS | ↑ (0.036) | ↑ (<0.001) | NS/NS |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hughes, D.J.; Kunická, T.; Schomburg, L.; Liška, V.; Swan, N.; Souček, P. Expression of Selenoprotein Genes and Association with Selenium Status in Colorectal Adenoma and Colorectal Cancer. Nutrients 2018, 10, 1812. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111812
Hughes DJ, Kunická T, Schomburg L, Liška V, Swan N, Souček P. Expression of Selenoprotein Genes and Association with Selenium Status in Colorectal Adenoma and Colorectal Cancer. Nutrients. 2018; 10(11):1812. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111812
Chicago/Turabian StyleHughes, David J., Tereza Kunická, Lutz Schomburg, Václav Liška, Niall Swan, and Pavel Souček. 2018. "Expression of Selenoprotein Genes and Association with Selenium Status in Colorectal Adenoma and Colorectal Cancer" Nutrients 10, no. 11: 1812. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111812