Physiological and Transcriptional Responses in Weaned Piglets Fed Diets with Varying Phosphorus and Calcium Levels

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diet, and Sample Collection
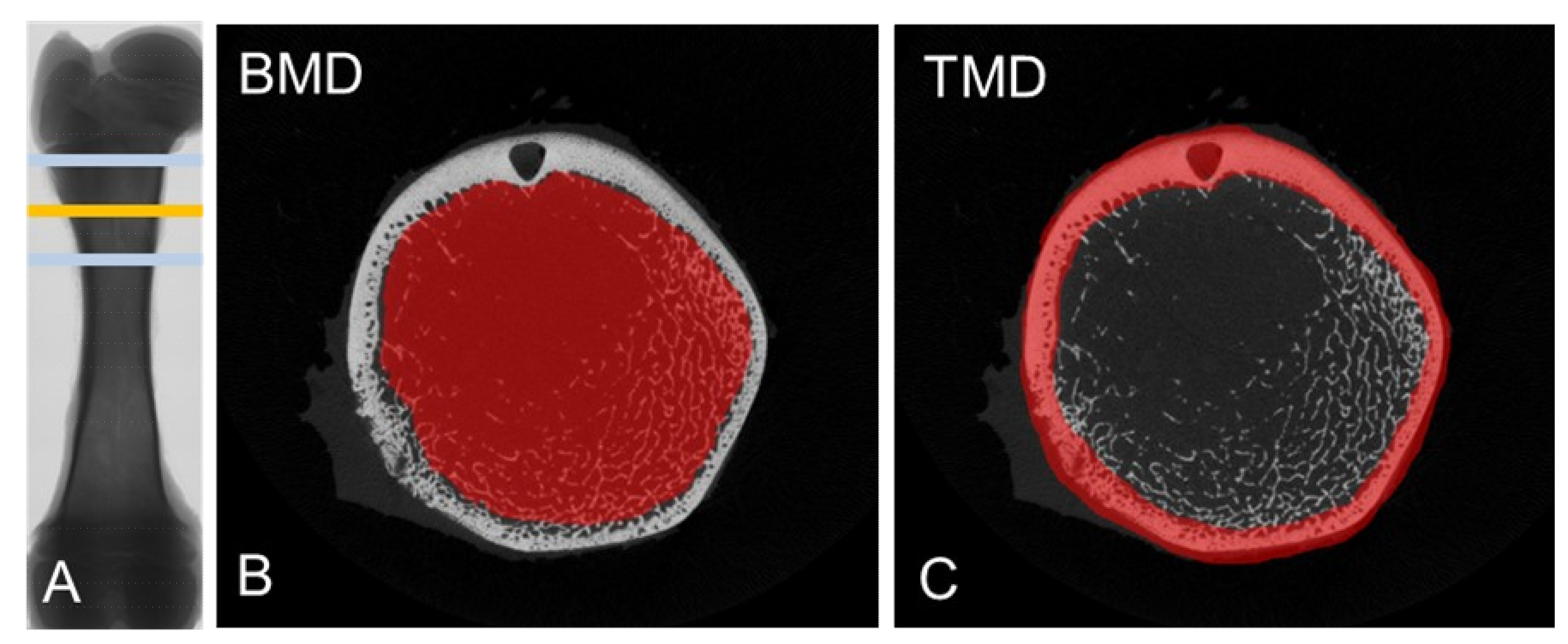
2.2. Measurement of Bone Characteristics
2.3. Measurements of Serum Parameters
2.4. RNA Isolation
2.5. Microarray Processing
2.6. Data Analysis
3. Results
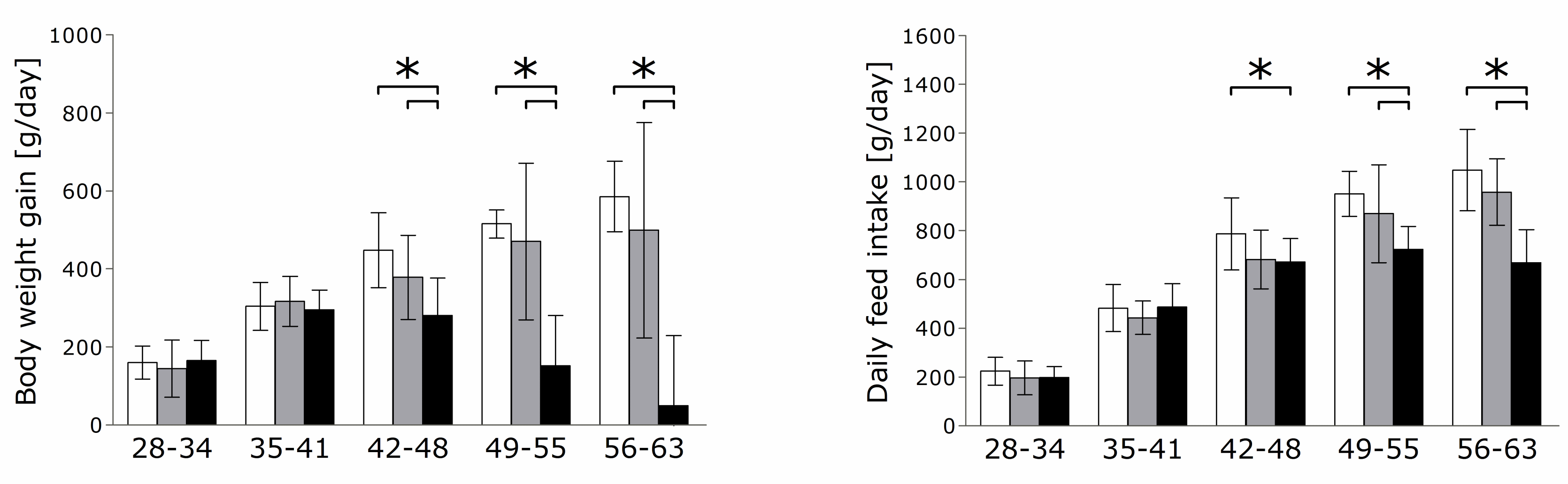
3.1. Performance and Feed Intake
3.2. Bone Characteristics
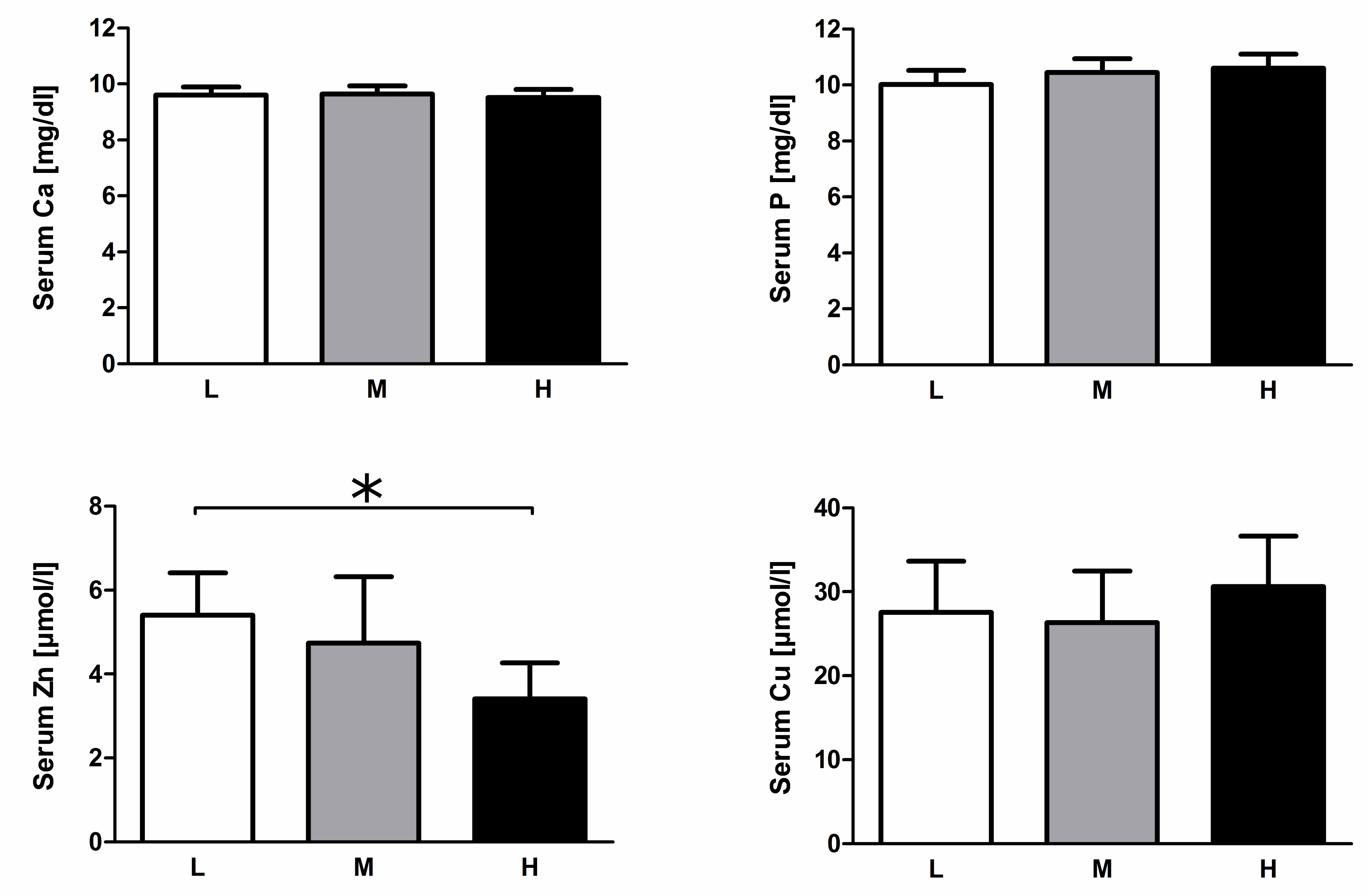
3.3. Serum Mineral Levels
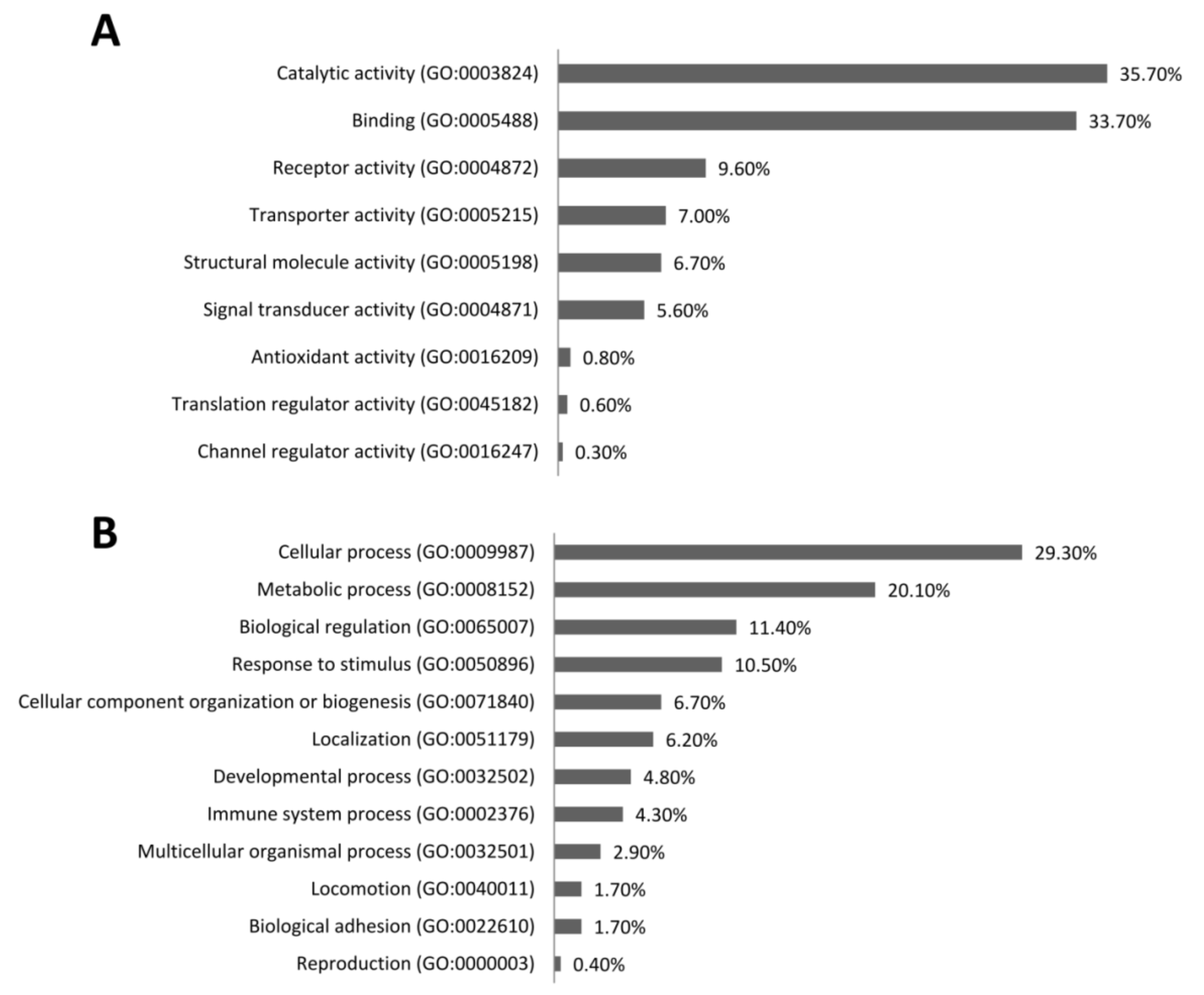
3.4. PBMC Gene Expression Pattern
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berndt, T.; Kumar, R. Novel Mechanisms in the regulation of phosphorus homeostasis. Physiology 2009, 24, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Tanck, E.; Homminga, J.; Van Lenthe, G.H.; Huiskes, R. Increase in bone volume fraction precedes architectural adaptation in growing bone. Bone 2001, 28, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Liesegang, B.A.L.; Ursprung, R.U.; Gasser, J.G.; Sassi, M.S.; Risteli, J.R.; Riond, J.-L.; Wanner, M. Influence of dietary phosphorus deficiency with or without addition of fumaric acid to a diet in pigs on bone parameters. J. Anim. Physiol. Anim. Nutr. 2002, 86, 1–16. [Google Scholar] [CrossRef]
- Nicodemo, M.L.F.; Scott, D.; Buchan, W.; Duncan, A.; Robins, S.P. Effects of variations in dietary calcium and phosphorus supply on plasma and bone osteocalcin concentrations and bone mineralization in growing pigs. Exp. Physiol. 1998, 83, 659–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, T.; Fernandez, J.A.; Engberg, R.M. Bone turnover in growing pigs fed three levels of dietary calcium. Can. J. Anim. Sci. 2000, 80, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Oster, M.; Just, F.; Büsing, K.; Wolf, P.; Polley, C.; Vollmar, B.; Muráni, E.; Ponsuksili, S.; Wimmers, K. Toward improved phosphorus efficiency in monogastrics—Interplay of serum, minerals, bone, and immune system after divergent dietary phosphorus supply in swine. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R917–R925. [Google Scholar] [CrossRef]
- Sommerville, B.A.; Maunder, E.; Ross, R.; Care, A.D.; Brown, R.C. Effect of dietary calcium and phosphorus depletion on vitamin d metabolism and calcium binding protein in the growing pig. Horm. Metab. Res. 1985, 17, 78–81. [Google Scholar] [CrossRef]
- Eklou-Kalonji, E.; Zerath, E.; Colin, C.; Lacroix, C.; Holy, X.; Denis, I.; Pointillart, A. Calcium-regulating hormones, bone mineral content, breaking load and trabecular remodeling are altered in growing pigs fed calcium-deficient diets. J. Nutr. 1999, 129, 188–193. [Google Scholar] [CrossRef]
- Sørensen, K.U.; Tauson, A.H.; Poulsen, H.D. Long term differentiated phosphorus supply from below to above requirement affects nutrient balance and retention, body weight gain and bone growth in growing-finishing pigs. Livest. Sci. 2018, 211, 14–20. [Google Scholar] [CrossRef]
- Sands, J.S.; Ragland, D.; Baxter, C.; Joern, B.C.; Sauber, T.E.; Adeola, O. Phosphorus bioavailability, growth performance, and nutrient balance in pigs fed high available phosphorus corn and phytase. J. Anim. Sci. 2001, 79, 2134–2142. [Google Scholar] [CrossRef]
- Max, D.; Brandsch, C.; Schumann, S.; Kühne, H.; Frommhagen, M.; Schutkowski, A.; Hirche, F.; Staege, M.S.; Stangl, G.I. Maternal vitamin D deficiency causes smaller muscle fibers and altered transcript levels of genes involved in protein degradation, myogenesis, and cytoskeleton organization in the newborn rat. Mol. Nutr. Food Res. 2014, 58, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Heyer, C.M.E.; Weiss, E.; Schmucker, S.; Rodehutscord, M.; Hoelzle, L.E.; Mosenthin, R.; Stefanski, V. The impact of phosphorus on the immune system and the intestinal microbiota with special focus on the pig. Nutr. Res. Rev. 2015, 28, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems Hiroshi. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, L.; Castañeda, S. Osteoimmunology: The Study of the Relationship Between the Immune System and Bone Tissue. Reumatol. Clínica 2013, 9, 303–315. [Google Scholar] [CrossRef]
- Liew, C.C.; Ma, J.; Tang, H.C.; Zheng, R.; Dempsey, A.A. The peripheral blood transcriptome dynamically reflects system wide biology: A potential diagnostic tool. J. Lab. Clin. Med. 2006, 147, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Just, F.; Oster, M.; Büsing, K.; Borgelt, L.; Murani, E.; Ponsuksili, S.; Wolf, P.; Wimmers, K. Lowered dietary phosphorus affects intestinal and renal gene expression to maintain mineral homeostasis with immunomodulatory implications in weaned piglets. BMC Genomics 2018, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.; Gerlinger, C.; Heide, K.; Just, F.; Borgelt, L.; Wolf, P.; Polley, C.; Vollmar, B.; Muráni, E.; Ponsuksili, S.; et al. Lower dietary phosphorus supply in pigs match both animal welfare aspects and resource efficiency. Ambio 2018, 47, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie. AfBN. Empfehlungen Zur Energie—Und Nährstoffversorgung von Schweinen; DLG-Verlag: Frankfurt, Germany, 2006. [Google Scholar]
- Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten. VDLUFA-Methodenbuch Band III Die chemische Untersuchung von Futtermitteln; Aufl.3 Ges.; VDLUFA-Verlag: Darmstadt, Germany, 1976. [Google Scholar]
- Hildebrand, T.; Rüegsegger, P. Quantification of bone microarchitecture with the structure model index. Comput. Methods Biomech. Biomed. Eng. 1997, 1, 15–23. [Google Scholar] [CrossRef]
- Bundesamt für Verbraucherschutz und Lebensmittelsicherheit (BVL). BVL F 0096:2013-04 Untersuchung von Futtermitteln—Bestimmung von Calcium, Natrium, Phosphor, Magnesium, Kalium, Schwefel, Eisen, Zink, Kupfer, Mangan und Kobalt in Futtermitteln mittels ICP-AES nach Druckaufschluss (Übernahme Der Gleichnamigen Norm DIN); Beuth Verlag GmbH: Berlin, Germany, 2013. [Google Scholar]
- Freeman, T.C.; Ivens, A.; Baillie, J.K.; Beraldi, D.; Barnett, M.W.; Dorward, D.; Downing, A.; Fairbairn, L.; Kapetanovic, R.; Raza, S.; et al. A gene expression atlas of the domestic pig. BMC Biol. 2012, 10, 1–21. [Google Scholar] [CrossRef]
- Edgar, R. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Kauffmann, A.; Gentleman, R.; Huber, W. ArrayQualityMetrics—A Bioconductor package for quality assessment of microarray data. Bioinformatics 2009, 25, 415–416. [Google Scholar] [CrossRef] [PubMed]
- Bourgon, R.; Gentleman, R.; Huber, W. Independent filtering increases detection power for high-throughput experiments. Proc. Natl. Acad. Sci. USA 2010, 107, 9546–9551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.D.; Tibshirani, R. Statistical significance for genome wide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]
- Alebrante, L.; Donzele, J.L.; Flávia, R.; Oliveira, M.; Saraiva, A.; Guimarães, S.E.F.; Ferreira, A.S. Available phosphorus levels in diets for pigs with high genetic potential for lean meat deposition kept in thermoneutral environment from 15 to 30 kg. Rev. Bras. Zootec. 2011, 40, 323–330. [Google Scholar] [CrossRef] [Green Version]
- González-Vega, J.C.; Liu, Y.; McCann, J.C.; Walk, C.L.; Loor, J.J.; Stein, H.H. Requirement for digestible calcium by eleven- to twenty-five-kilogram pigs as determined by growth performance, bone ash concentration, calcium and phosphorus balances, and expression of genes involved in transport of calcium in intestinal and kidney cell. J. Anim. Sci. 2016, 94, 3321–3334. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.G.; Mahan, D.C.; Graham, P.L. Changes in swine body composition from birth to 145 kg. J. Anim. Sci. 1983, 57, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Ammann, P.; Bourrin, S.; Bonjour, J.P.; Meyer, J.M.; Rizzoli, R. Protein undernutrition-induced bone loss is associated with decreased IGF-I levels and estrogen deficiency. J. Bone Miner. Res. 2000, 15, 683–690. [Google Scholar] [CrossRef]
- Lambe, N.R.; Wood, J.D.; McLean, K.A.; Walling, G.A.; Whitney, H.; Jagger, S.; Bünger, L. Effects of low protein diets on pigs with a lean genotype 2. Compositional traits measured with computed tomography (CT). Meat Sci. 2013, 95, 129–136. [Google Scholar] [CrossRef]
- Saraiva, A.; Donzele, J.L.; Oliveira, R.F.M.D.; Abreu, M.L.T.D.; De OliveiraSilva, F.C.; Santos, F.D.A. Available phosphorus levels in diets for swine from 15 to 30 kg genetically selected for meat deposition. Rev. Bras. Zootec. 2009, 38, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, A.; Donzele, J.L.; Oliveira, R.F.M.D.; Abreu, M.L.T.D.; Silva, F.C.D.O.; Vianna, R.A.; Lima, A.L. Available phosphorus levels in diets for 30 to 60 kg female pigs selected for meat deposition by maintaining calcium and available phosphorus ratio. Rev. Bras. Zootec. 2011, 40, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Hittmeier, L.J.; Grapes, L.; Lensing, R.L.; Rothschild, M.F.; Stahl, C.H. Genetic background influences metabolic response to dietary phosphorus restriction. J. Nutr. Biochem. 2006, 17, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Luecke, R.W.J.; Hoefer, A.; Brammell, W.S.; Schmidt, D.A. Calcium and zinc in parakeratosis of swine. J. Anim. Sci. 1957, 16, 3–11. [Google Scholar] [CrossRef]
- Bai, L.L.; Wu, F.; Liu, H.; Zhang, L.; Zhang, S.; Liu, L.; Piao, X.S.; Liu, Y.H.; Thacker, P.A.; Wang, F.L. Effects of dietary calcium levels on growth performance and bone characteristics in pigs in grower-finisher-transitional phase. Anim. Feed Sci. Technol. 2017, 224, 59–65. [Google Scholar] [CrossRef]
- Goretti, M.; Penido, M.G.; Alon, U.S. Phosphate homeostasis and its role in bone health. Pediatr. Nephrol. 2012, 27, 2039–2048. [Google Scholar] [CrossRef] [PubMed]
- Sapir-Koren, R.; Livshits, G. Bone mineralization and regulation of phosphate homeostasis. IBMS Bonekey 2011, 8, 286–300. [Google Scholar] [CrossRef]
- Corkum, C.P.; Ings, D.P.; Burgess, C.; Karwowska, S.; Kroll, W.; Michalak, T.I. Immune cell subsets and their gene expression profiles from human PBMC isolated by Vacutainer Cell Preparation Tube (CPTTM) and standard density gradient. BMC Immunol. 2015, 16, 48. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.; Horowitz, M.; Choi, Y. Osteoimmunology: Interactions of the bone and immune system. Endocr. Rev. 2008, 29, 403–440. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. SnapShot: Osteoimmunology. Cell Metab. 2015, 21, 502. [Google Scholar] [CrossRef]
- GeneCards. Available online: https://www.genecards.org (accessed on 7 August 2018).
- Stelzer, G.; Rosen, R.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Iny Stein, T.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The genecards suite: From gene data mining to disease genome sequence analysis. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar]
- Cravedi, P.; Leventhal, J.; Lakhani, P.; Ward, S.C.; Donovan, M.J.; Heeger, P.S. Immune cell-derived C3a and C5a costimulate human T cell alloimmunity. Am. J. Transplant. 2013, 13, 2530–2539. [Google Scholar] [CrossRef]
- Ignatius, A.; Schoengraf, P.; Kreja, L.; Liedert, A.; Recknagel, S.; Kandert, S.; Brenner, R.E.; Schneider, M.; Lambris, J.D.; Huber-Lang, M. Complement C3a and C5a modulate osteoclast formation and inflammatory response of osteoblasts in synergism with IL-1β. J. Cell. Biochem. 2011, 112, 2594–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, G.H.; Blesso, C.N. Dietary and endogenous sphingolipid metabolism in chronic inflammation. Nutrients 2017, 9, 1180. [Google Scholar] [CrossRef] [PubMed]
- Shockley, K.R.; Lazarenko, O.P.; Czernik, P.J.; Rosen, C.J.; Churchill, G.A.; Lecka-Czernik, B. PPARγ2 nuclear receptor controls multiple regulatory pathways of osteoblast differentiation from marrow mesenchymal stem cells. J. Cell. Biochem. 2009, 106, 232–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, I.; Engel, C.; Niestroj, A.J.; Kehlen, A.; Rahfeld, J.U.; Kleinschmidt, M.; Lehmann, K.; Roßner, S.; Demuth, H.U. A non-canonical function of eukaryotic elongation factor 1A1: Regulation of interleukin-6 expression. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 965–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, T.; Nara, K.; Yoshikawa, H.; Suzuki, N. Txk, a member of the non-receptor tyrosine kinase of the Tec family, forms a complex with poly (ADP-ribose) polymerase 1 and elongation factor 1α and regulates interferon-γ gene transcription in Th1 cells. Clin. Exp. Immunol. 2007, 147, 164–175. [Google Scholar] [CrossRef]
- Lönnerdal, B. Dietary Factors Influencing Zinc Absorption. J. Nutr. 2000, 130, 1378–1383. [Google Scholar] [CrossRef]
- Wood, R.J.; Zheng, J.J. High dietary calcium intakes reduce zinc absorption and balance in humans. Am. J. Clin. Nutr. 1997, 65, 1803–1809. [Google Scholar] [CrossRef] [Green Version]
- Revy, P.-S.; Jondreville, C.; Dourmad, J.-Y.; Guinotte, F.; Nys, Y. Bioavailability of two sources of zinc in weanling pigs. Anim. Res. 2002, 51, 315–326. [Google Scholar] [Green Version]
- Lewis, P.K., Jr.; Hoekstra, W.G.; Grummer, R.H. Restricted calcium feeding versus zinc supplementation for the control of parakeratosis in swine. J. Anim. Sci. 1957, 16, 578–588. [Google Scholar] [CrossRef]
- Magalhaes, J.G.; Tattoli, I.; Girardin, S.E. The intestinal epithelial barrier: How to distinguish between the microbial flora and pathogens. Semin. Immunol. 2007, 19, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Chengprapakorn, W. Toll like Receptor 2 and Toll like Receptor 4 Activation and Vitamin D Effects in Mouse Osteoblasts. PhD Thesis, University of California, Los Angeles, CA, USA, 2016. [Google Scholar]
- BioCyc. Available online: https://biocyc.org (accessed on 17 October 2018).
- Caspi, R.; Billington, R.; Ferrer, L.; Foerster, H.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Mueller, A.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of pathway/genome databases. Nucleic Acids Res. 2016, 44, D471–D480. [Google Scholar] [CrossRef] [PubMed]
- Katsoulidis, E.; Li, Y.; Mears, H.; Platanias, L.C. The p38 mitogen-activated protein kinase pathway in interferon signal transduction. J. Interferon Cytokine Res. 2005, 25, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Arts, R.J.W.; Joosten, L.A.B.; van der Meer, J.W.M.; Netea, M.G. TREM-1: Intracellular signaling pathways and interaction with pattern recognition receptors. J. Leukoc. Biol. 2013, 93, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Spahl, D.U.; Berendji-Grün, D.; Suschek, C.V.; Kolb-Bachofen, V.; Kröncke, K.-D. Regulation of zinc homeostasis by inducible NO synthase-derived NO: Nuclear metallothionein translocation and intranuclear Zn2+ release. Proc. Natl. Acad. Sci. USA 2003, 100, 13952–13957. [Google Scholar] [CrossRef]
- Kimball, S.R. Eukaryotic initiation factor eIF2. Int. J. Biochem. Cell. Biol. 1999, 31, 25–29. [Google Scholar] [CrossRef]
- Pain, V.M. Initiation of protein synthesis in eukaryotic cells. Eur. J. Biochem. 1996, 236, 747–771. [Google Scholar] [CrossRef]
- Ide, T.; Shimano, H.; Yoshikawa, T.; Yahagi, N.; Amemiya-Kudo, M.; Matsuzaka, T.; Nakakuki, M.; Yatoh, S.; Iizuka, Y.; Tomita, S.; et al. Cross-Talk between Peroxisome Proliferator-Activated Receptor (PPAR) α and Liver X Receptor (LXR) in nutritional regulation of fatty acid metabolism. II. LXRs suppress lipid degradation gene promoters through inhibition of PPAR signaling. Mol. Endocrinol. 2003, 17, 1255–1267. [Google Scholar] [CrossRef]
- Hill, C.S.; Wynne, J.; Treisman, R. The Rho family GTPases RhoA, Rac1, and CDC42Hs regulate transcriptional activation by SRF. Cell 1995, 81, 1159–1170. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L | M | H | |||||
|---|---|---|---|---|---|---|---|
| Item | Unit | Mean | SD | Mean | SD | Mean | SD |
| Tissue mineral density (TMD) | g/mm3 | 1.06 | 0.03 | 1.07 | 0.04 | 1.10 | 0.02 |
| Bone mineral density (BMD) | g/mm3 | 0.08 a | 0.02 | 0.16 b | 0.03 | 0.15 b | 0.04 |
| Bone volume/Total volume (BV/TV) | % | 16.68 a | 3.69 | 25.31 b | 6.05 | 23.80 b | 5.22 |
| Structural model index (SMI) | 1.09 | 0.19 | 1.20 | 0.73 | 1.23 | 0.49 | |
| Trabecular number (TbN) | 1/mm | 0.94 a | 0.19 | 1.39 b | 0.21 | 1.23 a,b | 0.49 |
| Trabecular separation (TbSp) | mm | 3.44 b | 1.87 | 0.95 a | 0.51 | 1.13 a | 1.03 |
| Trabecular thickness (TbTh) | mm | 0.18 | 0.01 | 0.18 | 0.02 | 0.31 | 0.33 |
| Fracture load | N | 764.9 a | 110.9 | 999.9 b | 153.2 | 920.4 b | 208.2 |
| Maximum deflection | mm | 6.5 b | 2.0 | 4.6 a | 1.1 | 4.1 a | 0.7 |
| Pathway Name | z-Score | Responsive Transcripts |
|---|---|---|
| Biosynthesis | ||
| Superpathway of Inositol Phosphate Compounds * | - | ACP1, DUSP2, IPMK, PIK3C2A, PIP5K1B, PPP1R12A, PTEN, PTPN12, SET, SOCS3, TNS3 |
| EIF2 Signaling * | 1.13 | ATF3, EIF3K, PIK3C2A, RPL12, RPL7, RPL24, RPL34, RPS8, RPS19, RPS26, RPSA, SOS2 |
| CDP-diacylglycerol Biosynthesis I † | - | CDS1, GPAT3, GPAT4, LPCAT2, MBOAT7 |
| Triacylglycerol Biosynthesis † | - | DGAT2, GPAT3, GPAT4, LPCAT2, MBOAT7, PLPP3 |
| Interferon Signaling * | 2.00 | IFIT3, IFNGR1, JAK2, OAS1 |
| Immune Response | ||
| IL-10 Signaling ‡ | - | ARG2, CCR1, HMOX1, IL1A, IL1R2, IL1RAP, IL4R, IL18, IL18RAP, SOCS3 |
| IL-6 Signaling ‡ | 2.89 | CSNK2A1, IL1A, IL1R2, IL1RAP, IL6R, IL18, IL18RAP, JAK2, PIK3C2A, SOCS3, SOS2, TNFRSF1A |
| Acute Phase Response Signaling ‡ | 2.11 | C4BPA, F8, HMOX1, IL1A, IL1RAP, IL6R, IL18, JAK2, RIPK1, SOCS3, SOD2, SOS2, TNFRSF1A |
| NF-κB Signaling † | 3.05 | BMPR2, CSNK2A1, IL1A, IL1R2, IL18, IRAK4, PIK3C2A, RIPK1, TGFA, TGFBR1, TLR4, TNFRSF1A, TNFSF13B |
| Th1 and Th2 Activation Pathway † | - | BMPR2, CCR1, CD247, CD274, IFNGR1, IL4R, IL6R, IL18, JAK2, NOTCH2, PIK3C2A, SOCS3, TGFBR1 |
| T Helper Cell Differentiation † | - | BCL6, IFNGR1, IL4R, IL6R, IL18, TGFBR1, TNFRSF1A |
| Th1 Pathway * | 1.41 | CD247, CD274, IFNGR1, IL6R, IL18, JAK2, NOTCH2, PIK3C2A, SOCS3 |
| Fcγ Receptor mediated Phagocytosis in Macrophages and Monocytes † | 2.12 | ARPC2, FGR, GAB2, HCK, HMOX1, NCF1, PTEN, PTK2B |
| Granulocyte Adhesion and Diapedesis † | - | CCL14, CSF3R, CXCR2, IL1A, IL1R2, IL1RAP, IL18, IL18RAP, ITGAM, SELL, TNFRSF1A |
| Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses * | 2.00 | C3AR1, DDX58, IL1A, IL18, NOD1, OAS1, PIK3C2A, TLR4 |
| LXR/RXR Activation ‡ | 2.53 | ABCA1, ARG2, FDFT1, IL1A, IL1R2, IL1RAP, IL18, IL18RAP, LY96, MYLIP, TLR4, TNFRSF1A |
| LPS/IL-1 Mediated Inhibition of RXR Function † | 1.89 | ABCA1, ACSL1, ACSL4, ALDH1L2, GSTM3, IL1A, IL1R2, IL1RAP, IL18, IL18RAP, LY96, RARA, TLR4, TNFRSF1A |
| PPAR Signaling * | 2.65 | IL1A, IL1R2, IL1RAP, IL18, IL18RAP, SOS2, TNFRSF1A |
| Signaling | ||
| RhoA Signaling * | 0.38 | ABL2, ARHGAP6, ARPC2, LIMK2, LPAR6, PIP5K1B, PPP1R12A, PTK2B |
| p38 MAPK Signaling † | 2.83 | IL1A, IL1R2, IL1RAP, IL18, IL18RAP, IRAK4, RPS6KA3, TGFBR1, TNFRSF1A |
| TREM1 Signaling * | 2.45 | CASP5, IL18, JAK2, NOD1, TLR4, TREM1 |
| iNOS Signaling * | 2.24 | IFNGR1, IRAK4, JAK2, LY96, TLR4 |
| Gene Symbol | FC 1 | p Value | q Value | Function |
|---|---|---|---|---|
| IL22RA2 | 5.95 | <0.001 | 0.075 | cytokine receptor |
| RNF128 | 3.55 | <0.001 | 0.102 | transmembrane protein/ligase |
| ARG2 | 3.40 | <0.001 | 0.087 | arginine metabolism |
| C3AR1 | 3.22 | 0.001 | 0.123 | G-protein coupled receptor |
| IL1R2 | 3.14 | 0.002 | 0.144 | interleukin receptor |
| SPTSSA | −3.81 | <0.001 | 0.102 | sphingolipid biosynthesis |
| PLPP3 | −2.09 | 0.003 | 0.151 | membrane glycoprotein |
| EEF1A1 | −1.84 | 0.007 | 0.186 | elongation factor |
| PEBP1 | −1.81 | 0.009 | 0.201 | binding protein |
| APEX1 | −1.74 | 0.003 | 0.151 | endonuclease |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerlinger, C.; Oster, M.; Borgelt, L.; Reyer, H.; Muráni, E.; Ponsuksili, S.; Polley, C.; Vollmar, B.; Reichel, M.; Wolf, P.; et al. Physiological and Transcriptional Responses in Weaned Piglets Fed Diets with Varying Phosphorus and Calcium Levels. Nutrients 2019, 11, 436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020436
Gerlinger C, Oster M, Borgelt L, Reyer H, Muráni E, Ponsuksili S, Polley C, Vollmar B, Reichel M, Wolf P, et al. Physiological and Transcriptional Responses in Weaned Piglets Fed Diets with Varying Phosphorus and Calcium Levels. Nutrients. 2019; 11(2):436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020436
Chicago/Turabian StyleGerlinger, Christian, Michael Oster, Luisa Borgelt, Henry Reyer, Eduard Muráni, Siriluck Ponsuksili, Christian Polley, Brigitte Vollmar, Martin Reichel, Petra Wolf, and et al. 2019. "Physiological and Transcriptional Responses in Weaned Piglets Fed Diets with Varying Phosphorus and Calcium Levels" Nutrients 11, no. 2: 436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020436