Dietary Egg Sphingomyelin Prevents Aortic Root Plaque Accumulation in Apolipoprotein-E Knockout Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Serum Biochemical Analysis
2.3. Atherosclerosis Analysis
2.4. RNA Isolation, cDNA Synthesis, and qRT-PCR
2.5. Cecal Feces Lipid Extraction
2.6. Gut Microbiota Analysis
2.7. Statistical Analysis
3. Results
3.1. ESM Reduces Epididymal Fat Accumulation
3.2. ESM Significantly Lowers ALT Activity and Serum SAA
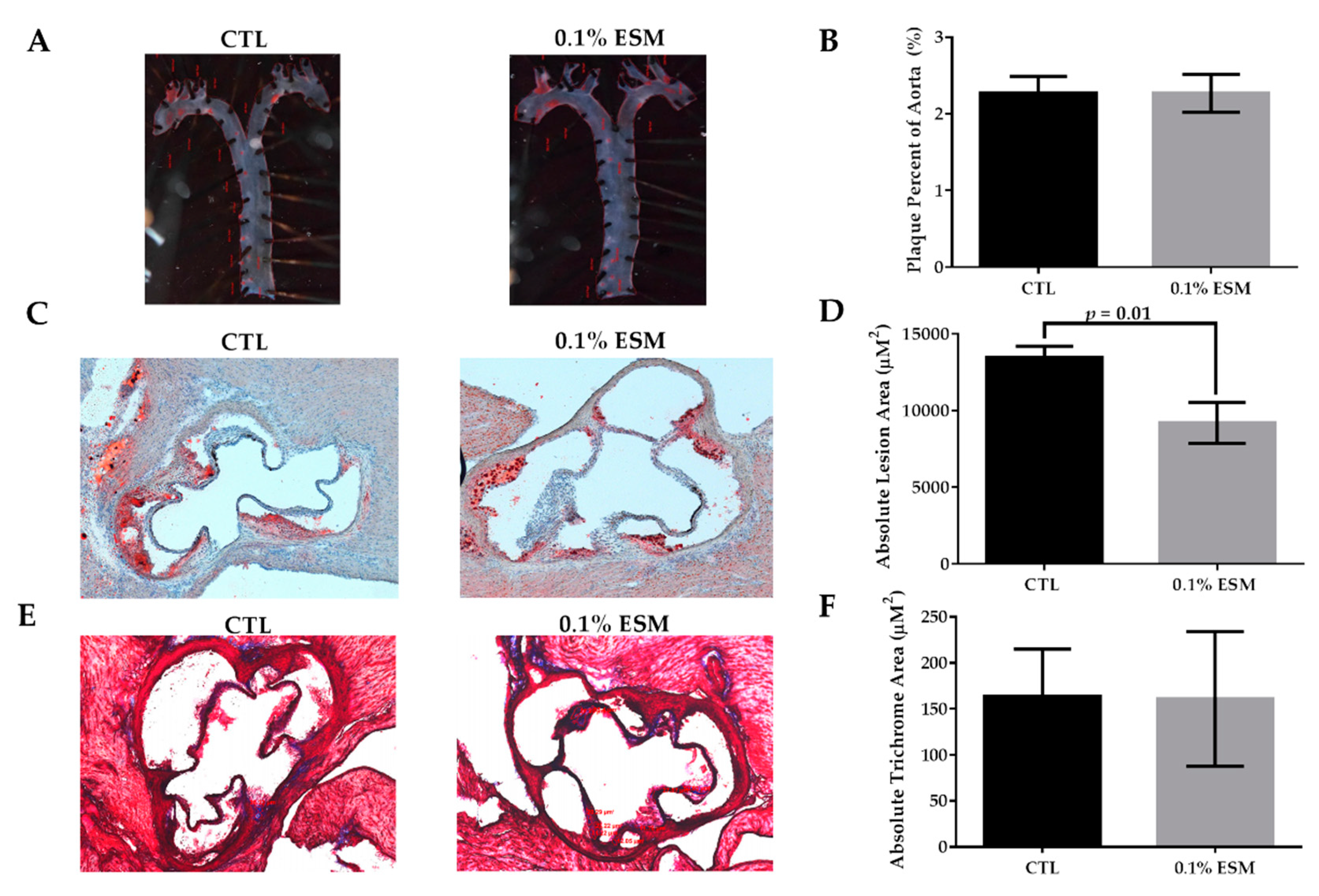
3.3. Aortic Root Plaque Accumulation Was Lower in ESM-Fed Mice
3.4. Effects of ESM on Gut Microbiota Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xu, J.; Murphy, S.L.; Kochanek, K.D.; Bastian, B.A. Deaths: Final data for 2013. Natl. Vital Stat. Rep. 2016, 64, 1–19. [Google Scholar]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; De Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics—2017 Update: A Report from the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Moore, K.J.; Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Lin, J.; Yang, R.; Tarr, P.T.; Wu, P.H.; Handschin, C.; Li, S.; Yang, W.; Pei, L.; Uldry, M.; Tontonoz, P.; et al. Hyperlipidemic effects of dietary saturated fats mediated through PGC-1beta coactivation of SREBP. Cell 2005, 120, 261–273. [Google Scholar] [CrossRef]
- Subramanian, S.; Han, C.Y.; Chiba, T.; McMillen, T.S.; Wang, S.A.; Haw, A.; Kirk, E.A.; O’Brien, K.D.; Chait, A.; Iii, A.H. Dietary cholesterol worsens adipose tissue macrophage accumulation and atherosclerosis in obese LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Boil. 2008, 28, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Goodspeed, L.; Wang, S.; Kim, J.; Zeng, L.; Ioannou, G.N.; Haigh, W.G.; Yeh, M.M.; Kowdley, K.V.; O’Brien, K.D.; et al. Dietary cholesterol exacerbates hepatic steatosis and inflammation in obese LDL receptor-deficient mice. J. Lipid Res. 2011, 52, 1626–1635. [Google Scholar] [CrossRef] [Green Version]
- Norris, G.H.; Blesso, C.N. Dietary sphingolipids: Potential for management of dyslipidemia and nonalcoholic fatty liver disease. Nutr. Rev. 2017, 75, 274–285. [Google Scholar] [CrossRef]
- Sacks, F.M.; Lichtenstein, A.H.; Wu, J.H.; Appel, L.J.; Creager, M.A.; Kris-Etherton, P.M.; Miller, M.; Rimm, E.B.; Rudel, L.L.; Robinson, J.G.; et al. Dietary Fats and Cardiovascular Disease: A Presidential Advisory from the American Heart Association. Circulation 2017, 136, 1–23. [Google Scholar] [CrossRef]
- Wilczek, M.M.; Olszewski, R.; Krupienicz, A. Trans-Fatty Acids and Cardiovascular Disease: Urgent Need for Legislation. Cardiology 2017, 138, 254–258. [Google Scholar] [CrossRef]
- Korakas, E.; Dimitriadis, G.; Raptis, A.; Lambadiari, V. Dietary Composition and Cardiovascular Risk: A Mediator or a Bystander? Nutrients 2018, 10, 1912. [Google Scholar] [CrossRef]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated fat, carbohydrate, and cardiovascular disease. Am. J. Clin. Nutr. 2010, 91, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Eussen, S.R.; Verhagen, H.; Klungel, O.H.; Garssen, J.; Van Loveren, H.; Van Kranen, H.J.; Rompelberg, C.J. Functional foods and dietary supplements: Products at the interface between pharma and nutrition. Eur. J. Pharm. 2011, 668, S2–S9. [Google Scholar] [CrossRef]
- Casas, R.; Estruch, R.; Sacanella, E. Influence of Bioactive Nutrients on the Atherosclerotic Process: A Review. Nutrients 2018, 10, 1630. [Google Scholar] [CrossRef]
- Andersen, C.J. Bioactive Egg Components and Inflammation. Nutrients 2015, 7, 7889–7913. [Google Scholar] [CrossRef] [Green Version]
- Vesper, H.; Schmelz, E.-M.; Nikolova-Karakashian, M.N.; Dillehay, D.L.; Lynch, D.V.; Merrill, A.H. Sphingolipids in Food and the Emerging Importance of Sphingolipids to Nutrition. J. Nutr. 1999, 129, 1239–1250. [Google Scholar] [CrossRef]
- Cohn, J.S.; Kamili, A.; Wat, E.; Chung, R.W.S.; Tandy, S. Dietary Phospholipids and Intestinal Cholesterol Absorption. Nutrients 2010, 2, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelz, E.M.; Roberts, P.C.; Kustin, E.M.; A Lemonnier, L.; Sullards, M.C.; Dillehay, D.L.; Merrill, A.H. Modulation of intracellular beta-catenin localization and intestinal tumorigenesis in vivo and in vitro by sphingolipids. Cancer Res. 2001, 61, 6723–6729. [Google Scholar]
- Chung, R.W.S.; Kamili, A.; Tandy, S.; Weir, J.M.; Gaire, R.; Wong, G.; Meikle, P.J.; Cohn, J.S.; Rye, K.-A. Dietary Sphingomyelin Lowers Hepatic Lipid Levels and Inhibits Intestinal Cholesterol Absorption in High-Fat-Fed Mice. PLoS ONE 2013, 8, 55949. [Google Scholar] [CrossRef] [PubMed]
- Norris, G.H.; Porter, C.M.; Jiang, C.; Millar, C.L.; Blesso, C.N. Dietary sphingomyelin attenuates hepatic steatosis and adipose tissue inflammation in high-fat-diet-induced obese mice. J. Nutr. Biochem. 2017, 40, 36–43. [Google Scholar] [CrossRef]
- Noh, S.K.; I Koo, S. Egg sphingomyelin lowers the lymphatic absorption of cholesterol and alpha-tocopherol in rats. J. Nutr. 2003, 133, 3571–3576. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.K.; Koo, S.I. Milk Sphingomyelin Is More Effective than Egg Sphingomyelin in Inhibiting Intestinal Absorption of Cholesterol and Fat in Rats. J. Nutr. 2004, 134, 2611–2616. [Google Scholar] [CrossRef] [Green Version]
- Nyberg, L.; Duan, R.-D.; Nilsson, Å. A mutual inhibitory effect on absorption of sphingomyelin and cholesterol. J. Nutr. Biochem. 2000, 11, 244–249. [Google Scholar] [CrossRef]
- Chung, R.W.S.; Wang, Z.; Bursill, C.A.; Wu, B.J.; Barter, P.J.; Rye, K.-A. Effect of long-term dietary sphingomyelin supplementation on atherosclerosis in mice. PLoS ONE 2017, 12, e0189523. [Google Scholar] [CrossRef]
- Center for Drug Evaluation and Research (CDER). Guidance for Industry; Food and Drug Administration: Silver Spring, MD, USA, 2005.
- Weihrauch, J.L.; Son, Y.-S. Phospholipid content of foods. J. Am. Oil Chem. Soc. 1983, 60, 1971–1978. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). USDA Food Composition Database. Available online: https://ndb.nal.usda.gov/ndb/ (accessed on 1 January 2019).
- Millar, C.L.; Norris, G.H.; Jiang, C.; Kry, J.; Vitols, A.; Garcia, C.; Park, Y.-K.; Lee, J.-Y.; Blesso, C.N. Long-Term Supplementation of Black Elderberries Promotes Hyperlipidemia, but Reduces Liver Inflammation and Improves HDL Function and Atherosclerotic Plaque Stability in Apolipoprotein E-Knockout Mice. Mol. Nutr. Food Res. 2018, 62, 1800404. [Google Scholar] [CrossRef]
- Norris, G.H.; Jiang, C.; Ryan, J.; Porter, C.M.; Blesso, C.N. Milk sphingomyelin improves lipid metabolism and alters gut microbiota in high fat diet-fed mice. J. Nutr. Biochem. 2016, 30, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Norris, G.H.; Porter, C.M.; Jiang, C.; Blesso, C.N. Dietary Milk Sphingomyelin Reduces Systemic Inflammation in Diet-Induced Obese Mice and Inhibits LPS Activity in Macrophages. Beverages 2017, 3, 37. [Google Scholar] [CrossRef]
- Duivenvoorden, I.; Voshol, P.J.; Rensen, P.C.; Van Duyvenvoorde, W.; A Romijn, J.; Emeis, J.J.; Havekes, L.M.; Nieuwenhuizen, W.F. Dietary sphingolipids lower plasma cholesterol and triacylglycerol and prevent liver steatosis in APOE*3Leiden mice. Am. J. Clin. Nutr. 2006, 84, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Basterr, M.J.; Hailemariam, T.K.; Hojjati, M.R.; Lu, S.; Liu, J.; Liu, R.; Zhou, H.; Jiang, X.C. The effect of dietary sphingolipids on plasma sphingomyelin metabolism and athersclerosis. Biochim. Biophys. Acta 2005, 1735, 130–134. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Animal models of atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.D.; Kip, K.E.; Marroquin, O.C.; Ridker, P.M.; Kelsey, S.F.; Shaw, L.J.; Pepine, C.J.; Sharaf, B.; Bairey Merz, C.N.; Sopko, G.; et al. Serum amyloid A as a predictor of coronary artery disease and cardiovascular outcome in women: The National Heart, Lung, and Blood Institute-Sponsored Women’s Ischemia Syndrome Evaluation (WISE). Circulation 2004, 109, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.C.; Jayne, C.; Thompson, J.; Wilson, P.G.; Yoder, M.H.; Webb, N.; Tannock, L.R. A brief elevation of serum amyloid A is sufficient to increase atherosclerosis. J. Lipid Res. 2015, 56, 286–293. [Google Scholar] [CrossRef]
- Vaisar, T.; Tang, C.; Babenko, I.; Hutchins, P.; Wimberger, J.; Suffredini, A.F.; Heinecke, J.W. Inflammatory remodeling of the HDL proteome impairs cholesterol efflux capacity. J. Lipid Res. 2015, 56, 1519–1530. [Google Scholar] [CrossRef]
- Milard, M.; Laugerette, F.; Durand, A.; Buisson, C.; Meugnier, E.; Loizon, E.; Louche-Pelissier, C.; Sauvinet, V.; Garnier, L.; Viel, S.; et al. Milk Polar Lipids in a High-Fat Diet Can Prevent Body Weight Gain: Modulated Abundance of Gut Bacteria in Relation with Fecal Loss of Specific Fatty Acids. Mol. Nutr. Food Res. 2019, 63, 1801078. [Google Scholar] [CrossRef]
- Jonsson, A.L.; Bäckhed, F. Role of gut microbiota in atherosclerosis. Nat. Rev. Cardiol. 2016, 14, 79–87. [Google Scholar] [CrossRef]
- Luedde, M.; Winkler, T.; Heinsen, F.-A.; Rühlemann, M.C.; Spehlmann, M.E.; Bajrovic, A.; Lieb, W.; Franke, A.; Ott, S.J.; Frey, N. Heart failure is associated with depletion of core intestinal microbiota. ESC Heart Fail. 2017, 4, 282–290. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Measure | Control | 0.1% ESM | p-Value |
|---|---|---|---|
| Body Weight (g) | 29.6 ± 0.34 | 28.8 ± 0.51 | 0.16 |
| Liver Weight (g) | 1.32 ± 0.05 | 1.26 ± 0.05 | 0.43 |
| % Liver Weight | 4.55 ± 0.12 | 4.49 ± 0.12 | 0.75 |
| Epididymal Fat Weight (g) | 0.46 ± 0.07 | 0.25 ± 0.03 | 0.01 |
| % Epididymal Weight | 1.60 ± 0.24 | 0.90 ± 0.08 | 0.01 |
| Spleen Weight (g) | 0.13 ± 0.01 | 0.15 ± 0.01 | 0.09 |
| % Spleen Weight | 0.46 ± 0.02 | 0.53 ± 0.03 | 0.05 |
| Foot Intake (g/mouse) | 3.18 ± 0.07 | 3.25 ± 0.08 | 0.51 |
| Measure | Control | 0.1% ESM | p-Value |
|---|---|---|---|
| Total Cholesterol (mg/dL) | 797 ± 33.8 | 744 ± 47.4 | 0.36 |
| HDL-C (mg/dL) | 78.5 ± 9.8 | 63.3 ± 12.9 | 0.35 |
| Non-HDL-C (mg/dL) | 719 ± 34.8 | 681 ± 49.6 | 0.53 |
| Triglycerides (mg/dL) | 86.9 ± 10.0 | 87.6 ± 13.6 | 0.97 |
| NEFAs (mg/dL) | 0.49 ± 0.05 | 0.47 ± 0.05 | 0.71 |
| Glucose (mg/dL) | 269 ± 19.6 | 288 ± 33.8 | 0.13 |
| ALT (U/L) | 86.5 ± 7.0 | 67.0 ± 4.6 | 0.03 |
| AST (U/L) | 104 ± 8.8 | 88.1 ± 6.5 | 0.16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Millar, C.L.; Norris, G.H.; Vitols, A.; Garcia, C.; Seibel, S.; Anto, L.; Blesso, C.N. Dietary Egg Sphingomyelin Prevents Aortic Root Plaque Accumulation in Apolipoprotein-E Knockout Mice. Nutrients 2019, 11, 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051124
Millar CL, Norris GH, Vitols A, Garcia C, Seibel S, Anto L, Blesso CN. Dietary Egg Sphingomyelin Prevents Aortic Root Plaque Accumulation in Apolipoprotein-E Knockout Mice. Nutrients. 2019; 11(5):1124. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051124
Chicago/Turabian StyleMillar, Courtney L., Gregory H. Norris, Addison Vitols, Chelsea Garcia, Samantha Seibel, Liya Anto, and Christopher N. Blesso. 2019. "Dietary Egg Sphingomyelin Prevents Aortic Root Plaque Accumulation in Apolipoprotein-E Knockout Mice" Nutrients 11, no. 5: 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051124