Effect of Vitamin D3 on the Postprandial Lipid Profile in Obese Patients: A Non-Targeted Lipidomics Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Subjects and Study Design
2.3. Samples,External Calibration Curve and Quality Control Preparation
2.4. UHPLC-ESI-QTOF-MS Conditions
2.5. Data Analysis and Statistics
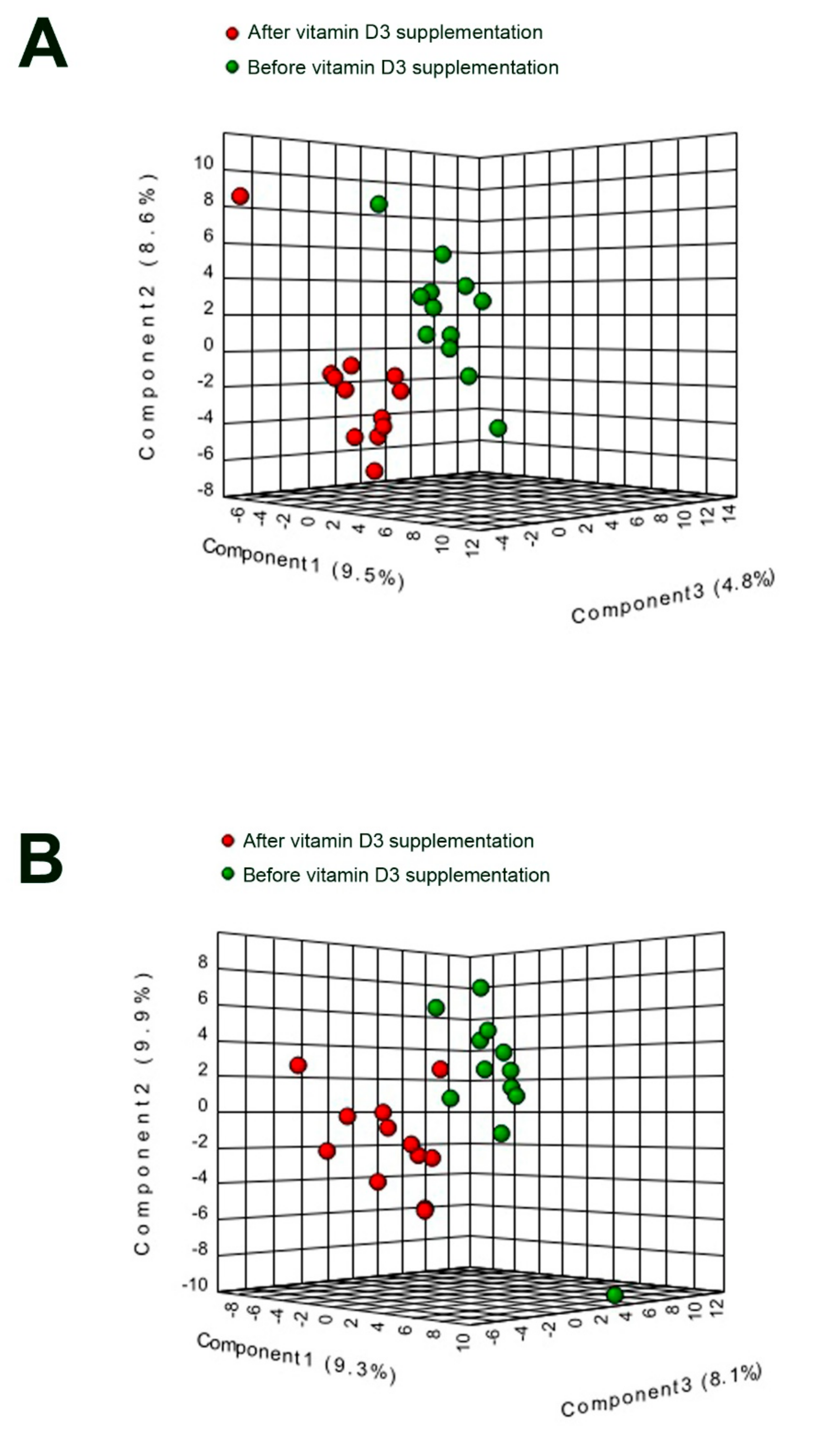
3. Results
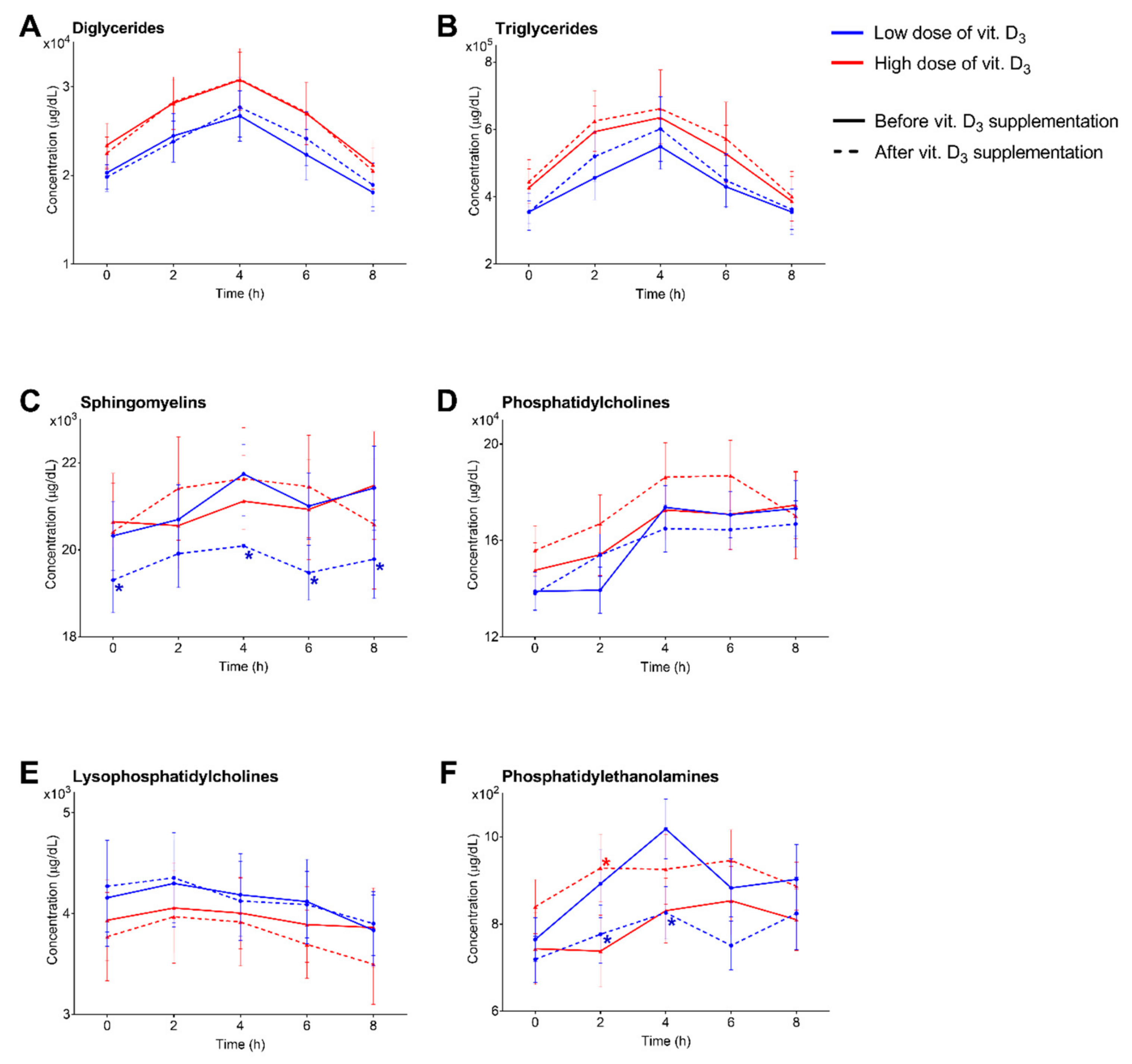
3.1. Characterization, Quantitation and Absorption Dynamics of Lipid Species
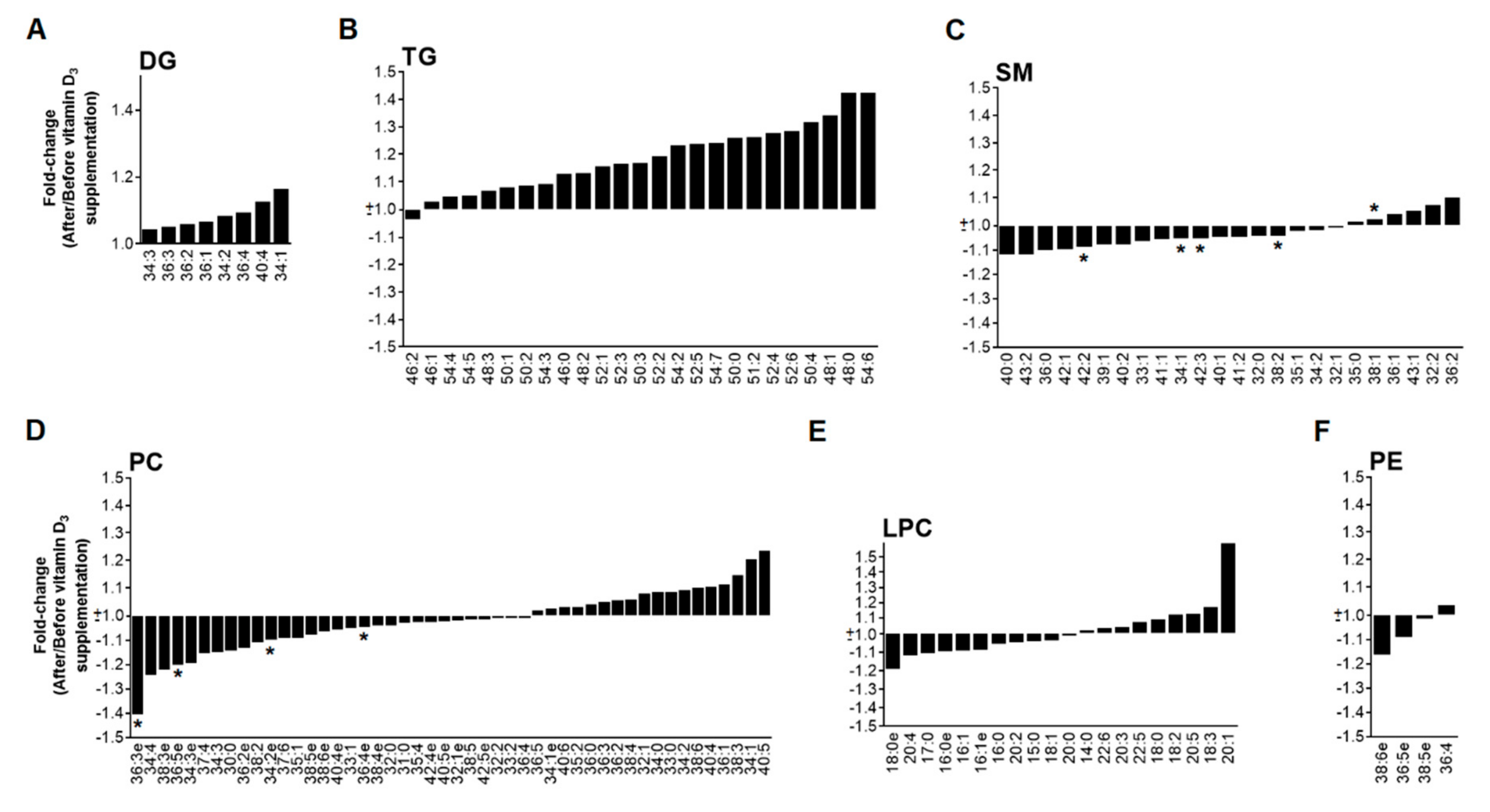
3.2. Low Dose of Vitamin D3 Decreases Total SM Levels and Specific PC and PE Species
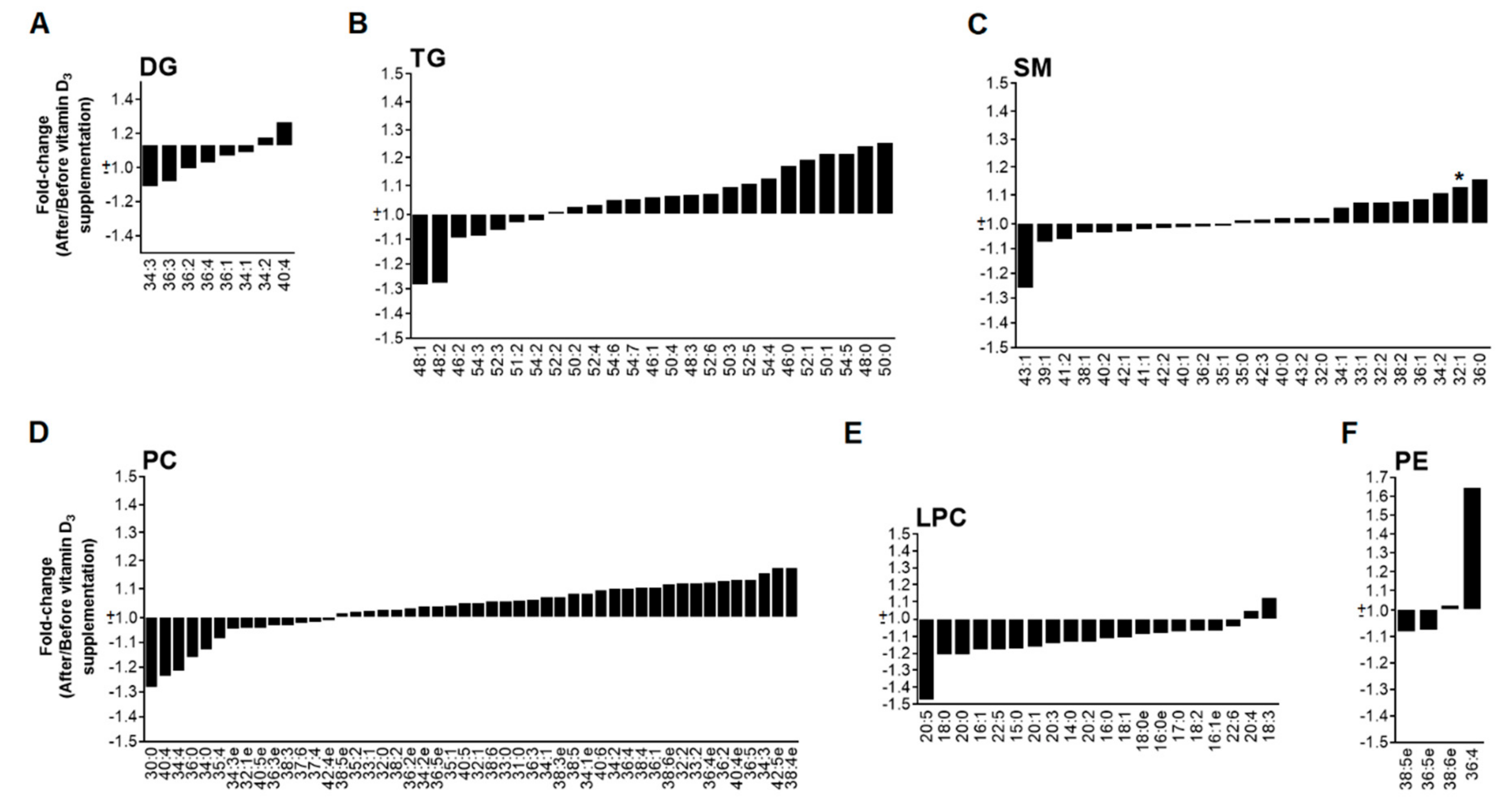
3.3. High Dose of Vitamin D3 Increases Specific PC and PE Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Vries, M.A.; Klop, B.; Alipour, A.; van de Geijn, G.J.; Prinzen, L.; Liem, A.H.; Valdivielso, P.; Rioja Villodres, J.; Ramirez-Bollero, J.; Castro Cabezas, M. In vivo evidence for chylomicrons as mediators of postprandial inflammation. Atherosclerosis 2015, 243, 540–545. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.A.; Klop, B.; Janssen, H.W.; Njo, T.L.; Westerman, E.M.; Castro Cabezas, M. Postprandial inflammation: Targeting glucose and lipids. Adv. Exp. Med. Biol. 2014, 824, 161–170. [Google Scholar] [PubMed]
- Alipour, A.; Elte, J.W.F.; van Zaanen, H.C.T.; Rietveld, A.P.; Castro Cabezas, M. Postprandial inflammation and endothelial dysfunction. Biochem. Soc. Trans. 2007, 35, 466–469. [Google Scholar] [CrossRef]
- Teeman, C.S.; Kurti, S.P.; Cull, B.J.; Emerson, S.R.; Haub, M.D.; Rosenkranz, S.K. Postprandial lipemic and inflammatory responses to high-fat meals: A review of the roles of acute and chronic exercise. Nutr. Metab. 2016, 13, 80. [Google Scholar]
- Klop, B.; Proctor, S.D.; Mamo, J.C.; Botham, K.M.; Castro Cabezas, M. Understanding postprandial inflammation and its relationship to lifestyle behaviour and metabolic diseases. Int. J. Vasc. Med. 2012, 2012, 947417. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Sheen, J.M.; Hu, W.L.; Hung, Y.C. Polyphenols and Oxidative Stress in Atherosclerosis-Related Ischemic Heart Disease and Stroke. Oxid. Med. Cell Longev. 2017, 2017, 8526438. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H.; Sheridan, Z.P. Studies on prevention of obesity, metabolic syndrome, diabetes, cardiovascular diseases and cancer by tea. J. Food Drug. Anal. 2018, 26, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, L.E.; Buchholz, A.C.; Mazurak, V.C. Inflammation, obesity, and fatty acid metabolism: Influence of n-3 polyunsaturated fatty acids on factors contributing to metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 1008–1024. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.L. The science of fatty acids and inflammation. Adv. Nutr. 2015, 6, 293S–301S. [Google Scholar] [CrossRef] [PubMed]
- Verseyden, C.; Meijssen, S.; van Dijk, H.; Jansen, H.; Castro Cabezas, M. Effects of atorvastatin on fasting and postprandial complement component 3 response in familial combined hyperlipidemia. J. Lipid Res. 2003, 44, 2100–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, B.; van Wijk, J.P.; Parra, S.; Castro Cabezas, M.; Hoepelman, I.M.; Alonso-Villaverde, C.; de Koning, E.J.; Camps, J.; Ferre, N.; Rabelink, T.J.; et al. Effects of rosiglitazone and metformin on postprandial paraoxonase-1 and monocyte chemoattractant protein-1 in human immunodeficiency virus-infected patients with lipodystrophy. Eur. J. Pharmacol. 2006, 544, 104–110. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Huskin, A.L.; Wolff, D.A.; Helenowski, I.B.; Rademaker, A.W. Fenofibrate reduces fasting and postprandial inflammatory responses among hypertriglyceridemia patients with the metabolic syndrome. Atherosclerosis 2008, 198, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Halkes, C.J.M.; van Dijk, H.; de Jaegere, P.P.T.; Plokker, H.W.M.; van der Helm, Y.; Erkelens, D.W.; Castro Cabezas, M. Postprandial increase of complement component 3 in normolipidemic patients with coronary artery disease. Effects of expanded-dose simvastatin. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1526–1530. [Google Scholar] [CrossRef]
- Silvagno, F.; Pescarmona, G. Spotlight on vitamin D receptor, lipid metabolism and mitochondria: Some preliminary emerging issues. Mol. Cell. Endocrinol. 2017, 450, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Girgis, C.M.; Clifton-Bligh, R.J.; Mokbel, N.; Cheng, K.; Gunton, J.E. Vitamin D signaling regulates proliferation, differentiation, and myotube size in C2C12 skeletal muscle cells. Endocrinology 2014, 155, 347–357. [Google Scholar] [CrossRef]
- Zhang, Y.; Leung, D.Y.; Richers, B.N.; Liu, Y.; Remigio, L.K.; Riches, D.W.; Goleva, E. Vitamin D inhibits monocyte/macrophage proinflammatory cytokine production by targeting MAPK phosphatase-1. J. Immunol. 2012, 188, 2127–2135. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Snyder, L.; Lin, Y.D.; Yang, L. Vitamin D and 1,25(OH)2D regulation of T cells. Nutrients 2015, 7, 3011–3021. [Google Scholar] [CrossRef]
- Prietl, B.; Treiber, G.; Pieber, T.R.; Amrein, K. Vitamin D and immune function. Nutrients 2013, 5, 2502–2521. [Google Scholar] [CrossRef]
- El-Fakhri, N.; McDevitt, H.; Shaikh, M.G.; Halsey, C.; Ahmed, S.F. Vitamin D and its effects on glucose homeostasis, cardiovascular function and immune function. Horm. Res. Paediatr. 2014, 81, 363–378. [Google Scholar] [CrossRef]
- Klop, B.; van de Geijn, G.J.; Birnie, E.; Njo, T.L.; Janssen, H.W.; Jansen, H.G.; Jukema, J.W.; Elte, J.W.; Castro Cabezas, M. Vitamin D3 mediated effects on postprandial leukocyte activation and arterial stiffness in men and women. Eur. J. Clin. Nutr. 2014, 68, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Querfeld, U. Vitamin D and inflammation. Pediatr. Nephrol. 2013, 28, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Boyan, B.D.; Sylvia, V.L.; Dean, D.D.; Pedrozo, H.; Del Toro, F.; Nemere, I.; Posner, G.H.; Schwartz, Z. 1,25-(OH)2D3 modulates growth plate chondrocytes via membrane receptor-mediated protein kinase C by a mechanism that involves changes in phospholipid metabolism and the action of arachidonic acid and PGE2. Steroids 1999, 64, 129–136. [Google Scholar] [PubMed]
- Sinha, A.; Hollingsworth, K.G.; Ball, S.; Cheetham, T. Improving the vitamin D status of vitamin D deficient adults is associated with improved mitochondrial oxidative function in skeletal muscle. J. Clin. Endocrinol. Metab. 2013, 98, E509–E513. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.A.; van der Meulen, N.; van de Geijn, G.M.; Klop, B.; van der Zwan, E.M.; Prinzen, L.; Birnie, E.; Westerman, E.M.; de Herder, W.W.; Castro Cabezas, M.; et al. Effect of a single dose of vitamin D3 on postprandial arterial stiffness and inflammation in vitamin D-deficient women. J. Clin. Endocrinol. Metab. 2017, 102, 992–1000. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Riera-Borrull, M.; Rodríguez-Gallego, E.; Hernández-Aguilera, A.; Luciano, F.; Ras, R.; Cuyàs, E.; Camps, J.; Segura-Carretero, A.; Menendez, J.A.; Joven, J.; et al. Exploring the process of energy generation in pathophysiology by targeted metabolomics: Performance of a simple and quantitative method. J. Am. Soc. Mass Spectrom. 2016, 27, 168–177. [Google Scholar] [CrossRef]
- Narvaez, C.J.; Simmons, K.M.; Brunton, J.; Salinero, A.; Chittur, S.V.; Welsh, J.E. Induction of STEAP4 correlates with 1,25-dihydroxyvitamin D3 stimulation of adipogenesis in mesenchymal progenitor cells derived from human adipose tissue. J. Cell Physiol. 2013, 228, 2024–2036. [Google Scholar] [CrossRef] [PubMed]
- Consiglio, M.; Viano, M.; Casarin, S.; Castagnoli, C.; Pescarmona, G.; Silvagno, F. Mitochondrial and lipogenic effects of vitamin D on differentiating and proliferating human keratinocytes. Exp. Dermatol. 2015, 24, 748–753. [Google Scholar] [CrossRef]
- Consiglio, M.; Destefanis, M.; Morena, D.; Foglizzo, V.; Forneris, M.; Pescarmona, G.; Silvagno, F. The vitamin D receptor inhibits the respiratory chain, contributing to the metabolic switch that is essential for cancer cell proliferation. PLoS ONE 2014, 9, e115816. [Google Scholar] [CrossRef]
- Merrill, A.H.; Sandhoff, K. Sphingolipids: metabolism and cell signaling. In Biochemistry of Lipids, Lipoproteins, 4th ed.; Vance, D.E., Vance, J.E., Eds.; Elsevier: London, UK, 2002; Volume 36, pp. 373–407. [Google Scholar]
- Okazaki, T.; Bell, R.M.; Hannun, Y.A. Sphingomyelin turnover induced by vitamin D3 in HL-60 cells. Role in cell differentiation. J. Biol. Chem. 1989, 264, 19076–19080. [Google Scholar]
- Wang, S.; Yang, X. Inter-organ regulation of adipose tissue browning. Cell. Mol. Life Sci. 2017, 74, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ji, X.; Wang, Q.; Li, J.Z. New insight into inter-organ crosstalk contributing to the pathogenesis of non-alcoholic fatty liver disease (NAFLD). Protein Cell 2018, 9, 164–177. [Google Scholar] [CrossRef]
- Geilen, C.C.; Bektas, M.; Wieder, T.; Kodelja, V.; Goerdt, S.; Orfanos, C.E. 1a,25-Dihydroxyvitamin D3 induces sphingomyelin hydrolysis in HaCaT cells via Tumor Necrosis Factor α. J. Biol. Chem. 1997, 272, 8997–9001. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.C.; Bektas, M.; Wieder, T.; Orfanos, C.E. The vitamin D, analogue, calcipotriol, induces sphingomyelin hydrolysis in human keratinocytes. FEBS Lett. 1996, 378, 88–92. [Google Scholar] [CrossRef]
- Magrassi, L.; Adorni, L.; Montorfano, G.; Rapelli, S.; Butti, G.; Berra, B.; Milanesi, G. Vitamin D metabolites activate the sphingomyelin pathway and induce death of glioblastoma cells. Acta Neurochir. 1998, 140, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Duan, R.D. Absorption and lipoprotein transport of sphingomyelin. J. Lipid Res. 2006, 47, 154–171. [Google Scholar] [CrossRef] [Green Version]
- Ohanian, J.; Ohanian, V. Sphingolipids in mammalian cell signalling. Cell. Mol. Life Sci. 2001, 58, 2053–2068. [Google Scholar] [CrossRef]
- Dressler, K.A.; Kan, C.C.; Kolesnick, R.N. Sphingomyelin synthesis is involved in adherence during macrophage differentiation of HL-60 cells. J. Biol. Chem. 1991, 266, 11522–11527. [Google Scholar]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef]
- Fan, M.; Sidhu, R.; Fujiwara, H.; Tortelli, B.; Zhang, J.; Davidson, C.; Walkley, S.U.; Bagel, J.H.; Vite, C.; Yanjanin, N.M.; et al. Identification of Niemann-Pick C1 disease biomarkers through sphingolipid profiling. J. Lipid Res. 2013, 54, 2800–2814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlitt, A.; Blankenberg, S.; Yan, D.; von Gizycki, H.; Buerke, M.; Werdan, K.; Bickel, C.; Lackner, K.J.; Meyer, J.; Rupprecht, H.J.; et al. Further evaluation of plasma sphingomyelin levels as a risk factor for coronary artery disease. Nutr. Metab. 2006, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Hashemipour, S.; Sarukhani, M.R.; Asef zadeh, S.; Ghazi, A.A.; Mehrtash, B.; Ahmadian Yazdi, M.H. Effect of different doses of parenteral vitamin D3 on serum 25 (OH) D concentrations. DARU 2009, 17, 26–29. [Google Scholar]
- Taylor, P.N.; Davies, J.S. A review of the growing risk of vitamin D toxicity from inappropriate practice. Br. J. Clin. Pharmacol. 2018, 84, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieth, R. Vitamin D supplementation, 25-hydroxyvitamin D concentrations, and safety. Am. J. Clin. Nutr. 1999, 69, 842–856. [Google Scholar] [CrossRef] [Green Version]
- Dalle Carbonare, L.; Valenti, M.; del Forno, F.; Caneva, E.; Pietrobelli, A. Vitamin D: Daily vs. monthly use in children and elderly—What is going on? Nutrients 2017, 9, 652. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low Dose Vit. D3 (n = 12) | High Dose Vit D3 (n = 12) | |
|---|---|---|
| Age (years) a | 29 ± 3 | 27 ± 2 |
| BMI (Kg/m2) a | 31.2 ± 1.3 | 33.0 ± 1.0 |
| Glucose (mmol/L) a | 5.2 ± 0.1 | 5.2 ± 0.1 |
| Total cholesterol (mmol/L) a | 5.1 ± 0.2 | 5.3 ± 0.3 |
| HDL-cholesterol (mmol/L) a | 1.5 ± 0.1 | 1.4 ± 0.1 |
| LDL-cholesterol (mmol/L) a | 3.1 ± 0.2 | 3.4 ± 0.3 |
| Leukocyte count (109/L) a,* | 6.6 ± 0.3 | 8.1 ± 0.6 |
| Monocyte count (109/L) a | 0.5 ± 0.06 | 0.5 ± 0.03 |
| Neutrophil count (109/L) a,* | 3.4 ± 0.3 | 4.4 ± 0.4 |
| 25-OH vitamin D (nmol/L) a | 27.3 ± 4.5 | 26.8 ± 3.6 |
| DG (μg/dL) | 20,312 ± 2095 | 23,425 ± 2467 |
| LPC (μg/dL) | 4158 ± 482 | 3935 ± 403 |
| PC (μg/dL) | 139,244 ± 7972 | 147,992 ± 11,465 |
| PE (μg/dL) | 765 ± 50 | 744 ± 82 |
| SM (μg/dL) | 20,325 ± 793 | 20,647 ± 1115 |
| TG (μg/dL) | 355,163 ± 54,657 | 427,374 ± 55,708 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Arroyo, S.; Hernández-Aguilera, A.; de Vries, M.A.; Burggraaf, B.; van der Zwan, E.; Pouw, N.; Joven, J.; Castro Cabezas, M. Effect of Vitamin D3 on the Postprandial Lipid Profile in Obese Patients: A Non-Targeted Lipidomics Study. Nutrients 2019, 11, 1194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051194
Fernández-Arroyo S, Hernández-Aguilera A, de Vries MA, Burggraaf B, van der Zwan E, Pouw N, Joven J, Castro Cabezas M. Effect of Vitamin D3 on the Postprandial Lipid Profile in Obese Patients: A Non-Targeted Lipidomics Study. Nutrients. 2019; 11(5):1194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051194
Chicago/Turabian StyleFernández-Arroyo, Salvador, Anna Hernández-Aguilera, Marijke A. de Vries, Benjamin Burggraaf, Ellen van der Zwan, Nadine Pouw, Jorge Joven, and Manuel Castro Cabezas. 2019. "Effect of Vitamin D3 on the Postprandial Lipid Profile in Obese Patients: A Non-Targeted Lipidomics Study" Nutrients 11, no. 5: 1194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051194