Taurine Supplementation Increases Post-Exercise Lipid Oxidation at Moderate Intensity in Fasted Healthy Males

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Sample Size
2.2. Study Design
2.3. Taurine and Placebo Supplementation
2.4. Maximal Oxygen Uptake (VO2 max) Test and Basal Blood Sample Collection
2.5. Experimental Trials
2.5.1. Anthropometric Data
2.5.2. Biochemical Analysis
2.5.3. Glycerol Analysis
2.5.4. Taurine Levels
2.5.5. Lipid and Glycosidic Oxidation by Indirect Calorimetry
2.6. Statistical Analysis
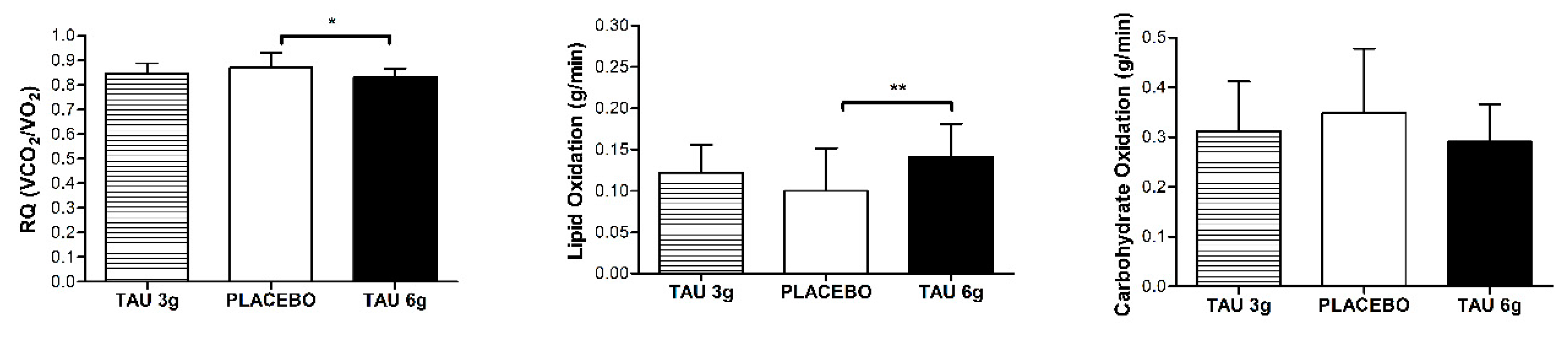
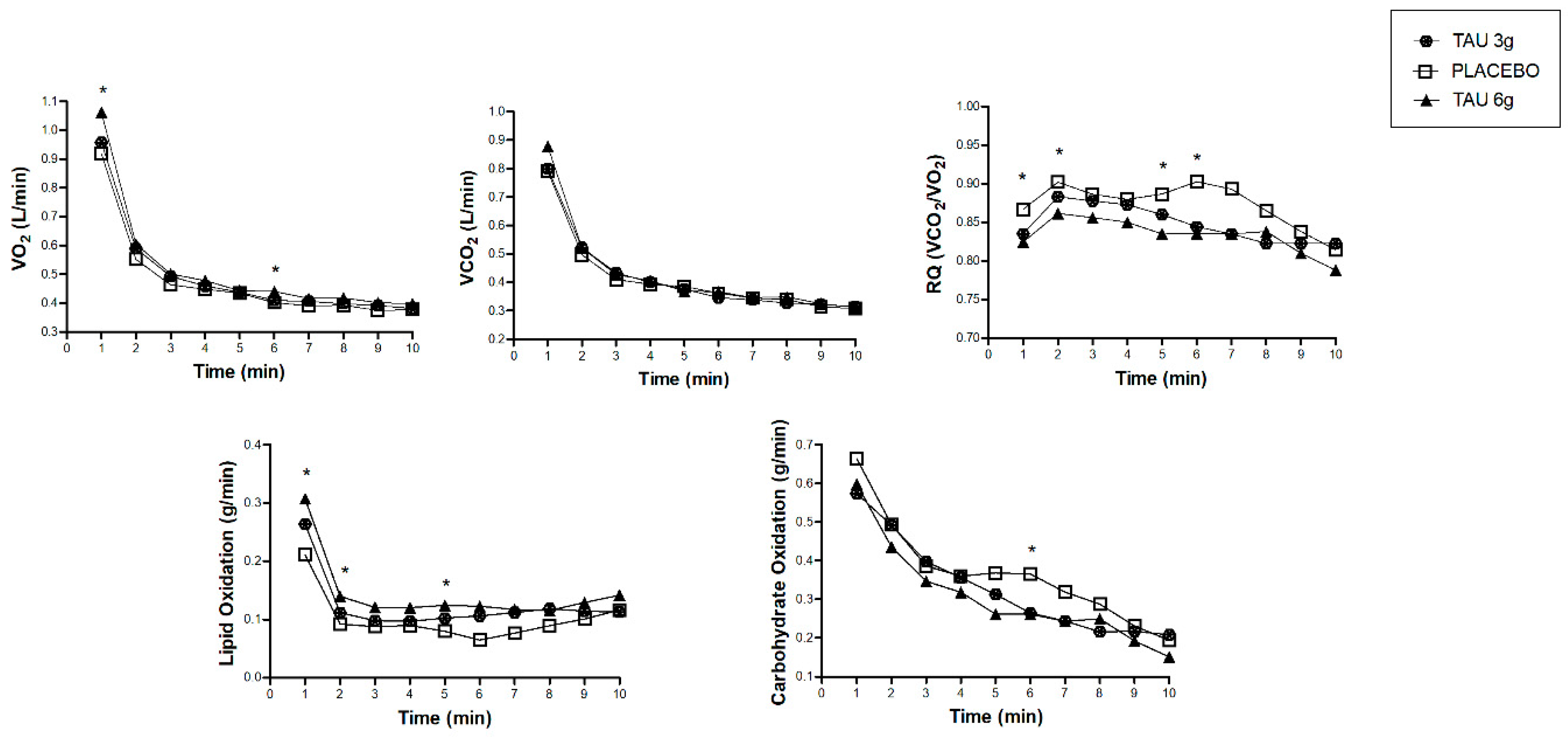
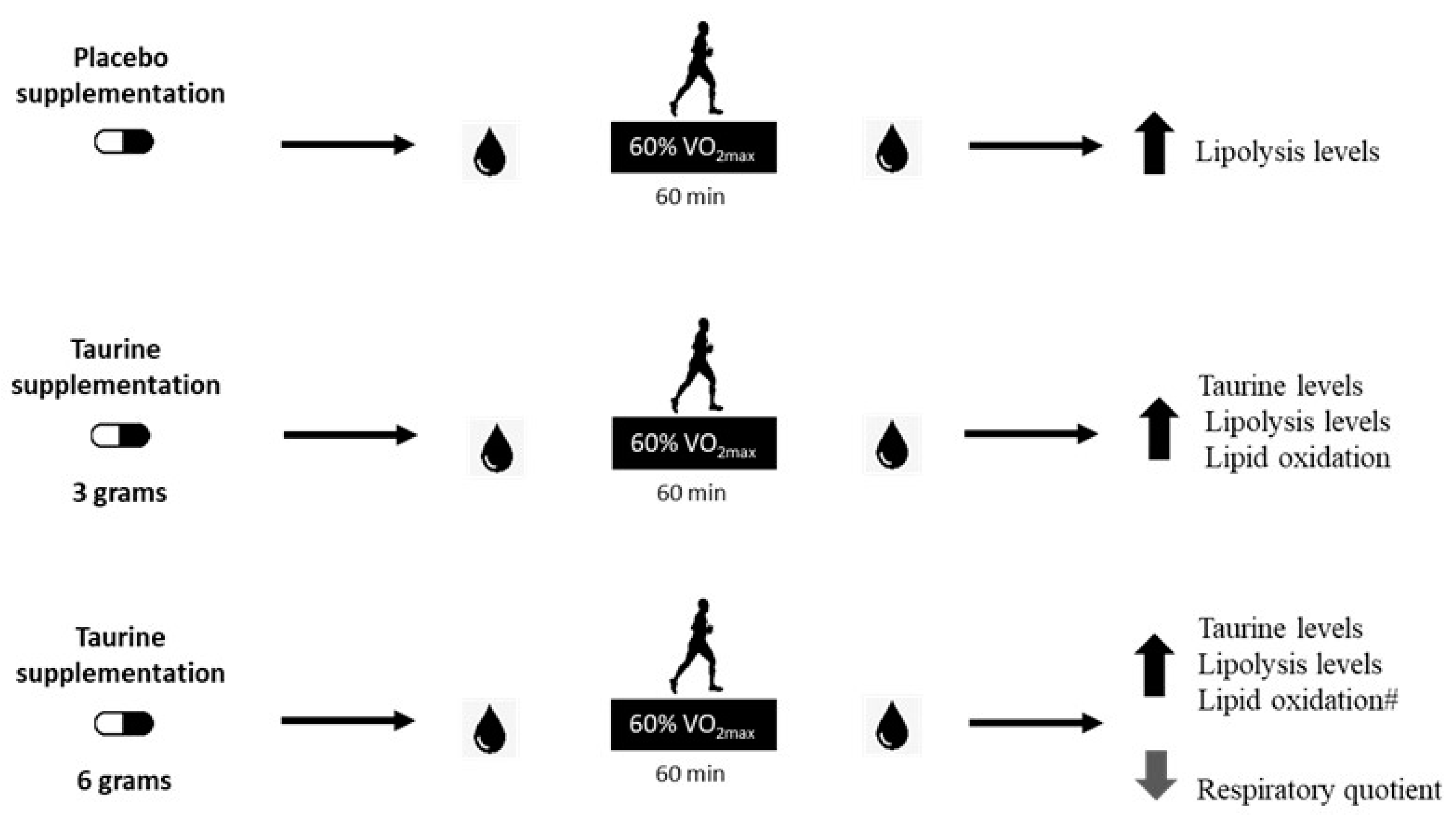
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ohyama, K.; Nogusa, Y.; Suzuki, K.; Shinoda, K.; Kajimura, S.; Bannai, M. A combination of exercise and capsinoid supplementation additively suppresses diet-induced obesity by increasing energy expenditure in mice. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E315–E323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisneros, L.C.V.; López-Uriarte, P.; López-Espinoza, A.; Meza, M.N.; Espinoza-Gallardo, A.C.; Aburto, M.B.G. Efectos del té verde y su contenido de galato de epigalocatequina (EGCG) sobre el peso corporal y la masa grasa en humanos. Una revisión sistemática. Nutrición Hospitalaria 2017, 34, 731–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneshiro, T.; Matsushita, M.; Hibi, M.; Tone, H.; Takeshita, M.; Yasunaga, K.; Katsuragi, Y.; Kameya, T.; Sugie, H.; Saito, M. Tea catechin and caffeine activate brown adipose tissue and increase cold-induced thermogenic capacity in humans. Am. J. Clin. Nutr. 2017, 105, 873–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, M.; Iwagaki, Y.; Sugawara, S.; Kushida, M.; Okouchi, R.; Yamamoto, K.; Tsuduki, T. Effects of Japanese diet in combination with exercise on visceral fat accumulation. Nutrition 2019, 57, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Jeukendrup, A.E. Optimizing fat oxidation through exercise and diet. Nutrition 2004, 20, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.E.; Tallan, H.H.; Lin, Y.Y.; Gaull, G.E. Taurine: Biological Update. Annu. Rev. Biochem. 1986, 55, 427–453. [Google Scholar] [CrossRef] [PubMed]
- Huxtable, R.J. Expanding the circle 1975-1999: Sulfur biochemistry and insights on the biological functions of taurine. Adv. Exp. Med. Biol. 2000, 483, 1–25. [Google Scholar]
- Sturman, J.A. Taurine in development. Physiol. Rev. 1993, 73, 119–147. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Shimada, K.; Jong, C.J.; Ito, T.; Azuma, J.; Takahashi, K. Effect of taurine and potential interactions with caffeine on cardiovascular function. Amino Acids 2014, 46, 1147–1157. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Jong, C.J.; Ramila, K.C.; Azuma, J. Physiological roles of taurine in heart and muscle. J. Biomed. Sci. 2010, 17, S2. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.W.; Minotto, J.B.; De Oliveira, M.R.; Zanotto-Filho, A.; Behr, G.A.; Rocha, R.F.; Moreira, J.C.; Klamt, F. Scavenging and antioxidant potential of physiological taurine concentrations against different reactive oxygen/nitrogen species. Pharmacol. Rep. 2010, 62, 185–193. [Google Scholar] [CrossRef]
- Murakami, S. Role of taurine in the pathogenesis of obesity. Mol. Nutr. Food Res. 2015, 59, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, F.G.; Barbieri, R.A.; Carvalho, M.B.; Dato, C.C.; Campos, E.Z.; Gobbi, R.B.; Papoti, M.; Da Silva, A.S.R.; De Freitas, E.C. Taurine supplementation can increase lipolysis and affect the contribution of energy systems during front crawl maximal effort. Amino Acids 2018, 50, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Martiniano, A.C.D.A.; De Carvalho, F.G.; Marchini, J.S.; Garcia, S.B.; Junior, J.E.; Mauad, F.M.; Da Silva, A.S.R.; De Moraes, C.; De Freitas, E.C. Effects of Taurine Supplementation on Adipose Tissue of Obese Trained Rats. Adv. Exp. Med. Biol. 2015, 803, 707–714. [Google Scholar] [CrossRef]
- Liu, Z.; Qi, B.; Zhang, M.; Izumi, I.; Kagamimori, S.; Sokejima, S.; Yamagami, T. Role of taurine supplementation to prevent exercise-induced oxidative stress in healthy young men. Amino Acids 2004, 26, 203–207. [Google Scholar] [CrossRef]
- Milioni, F.; Malta, E.; Rocha, L.G.S.D.A.; Mesquita, C.A.A.; De Freitas, E.C.; Zagatto, A.M. Acute administration of high doses of taurine does not substantially improve high-intensity running performance and the effect on maximal accumulated oxygen deficit is unclear. Appl. Physiol. Nutr. Metab. 2016, 41, 498–503. [Google Scholar] [CrossRef]
- Ra, S.-G.; Choi, Y.; Akazawa, N.; Ohmori, H.; Maeda, S. Taurine supplementation attenuates delayed increase in exercise-induced arterial stiffness. Appl. Physiol. Nutr. Metab. 2016, 41, 618–623. [Google Scholar] [CrossRef]
- Da Silva, L.A.; Tromm, C.B.; Bom, K.F.; Mariano, I.; Pozzi, B.; Da Rosa, G.L.; Tuon, T.; Da Luz, G.; Vuolo, F.; Petronilho, F.; et al. Effects of taurine supplementation following eccentric exercise in young adults. Appl. Physiol. Nutr. Metab. 2014, 39, 101–104. [Google Scholar] [CrossRef]
- Balshaw, T.; Bampouras, T.M.; Barry, T.J.; Sparks, A. The effect of acute taurine ingestion on 3-km running performance in trained middle-distance runners. Amino Acids 2013, 44, 555–561. [Google Scholar] [CrossRef]
- Rutherford, J.A.; Spriet, L.L.; Stellingwerff, T. The effect of acute taurine ingestion on endurance performance and metabolism in well-trained cyclists. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 322–329. [Google Scholar] [CrossRef]
- Galloway, S.D.; Talanian, J.L.; Shoveller, A.K.; Heigenhauser, G.J.F.; Spriet, L.L. Seven days of oral taurine supplementation does not increase muscle taurine content or alter substrate metabolism during prolonged exercise in humans. J. Appl. Physiol. 2008, 105, 643–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spriet, L.L. New insights into the interaction of carbohydrate and fat metabolism during exercise. Sports Med. 2014, 44, S87–S96. [Google Scholar] [CrossRef] [Green Version]
- Bonfleur, M.L.; Borck, P.C.; Ribeiro, R.A.; Caetano, L.C.; Soares, G.M.; Carneiro, E.M.; Balbo, S.L. Improvement in the expression of hepatic genes involved in fatty acid metabolism in obese rats supplemented with taurine. Life Sci. 2015, 135, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ueki, I.; Stipanuk, M.H. 3T3-L1 adipocytes and rat adipose tissue have a high capacity for taurine synthesis by the cysteine dioxygenase/cysteinesulfinate decarboxylase and cysteamine dioxygenase pathways. J. Nutr. 2008, 139, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, T.S.; Vendelbo, M.H.; Jessen, N.; Pedersen, S.B.; Jørgensen, J.O.L.; Lund, S.; Møller, N. Fasting, But Not Exercise, Increases Adipose Triglyceride Lipase (ATGL) Protein and Reduces G(0)/G(1) Switch Gene 2 (G0S2) Protein and mRNA Content in Human Adipose Tissue. J. Clin. Endocrinol. Metab. 2011, 96, E1293–E1297. [Google Scholar] [CrossRef] [Green Version]
- Samra, J.S.; Clark, M.L.; Humphreys, S.M.; Macdonald, I.A.; Frayn, K.N. Regulation of lipid metabolism in adipose tissue during early starvation. Am. J. Physiol. Metab. 1996, 271, E541–E546. [Google Scholar] [CrossRef]
- Schoenfeld, B.J.; Aragon, A.A.; Wilborn, C.; Krieger, J.W.; Sonmez, G.T. Body composition changes associated with fasted versus non-fasted aerobic exercise. J. Int. Soc. Sports Nutr. 2014, 11, 54. [Google Scholar] [CrossRef]
- Chenevière, X.; Borrani, F.; Droz, D.; Gojanovic, B.; Malatesta, D. Effects of 2 different prior endurance exercises on whole-body fat oxidation kinetics: Light vs. heavy exercise. Appl. Physiol. Nutr. Metab. 2012, 37, 955–964. [Google Scholar] [CrossRef]
- Jensen, L.; Gejl, K.D.; Ørtenblad, N.; Nielsen, J.L.; Bech, R.D.; Nygaard, T.; Sahlin, K.; Frandsen, U. Carbohydrate restricted recovery from long term endurance exercise does not affect gene responses involved in mitochondrial biogenesis in highly trained athletes. Physiol. Rep. 2015, 3, e12184. [Google Scholar] [CrossRef] [Green Version]
- Van Proeyen, K.; Szlufcik, K.; Nielens, H.; Ramaekers, M.; Hespel, P. Beneficial metabolic adaptations due to endurance exercise training in the fasted state. J. Appl. Physiol. 2010, 110, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Rosa, F.T.; De Freitas, E.C.; Deminice, R.; Jordão, A.A.; Marchini, J.S. Oxidative stress and inflammation in obesity after taurine supplementation: A double-blind, placebo-controlled study. Eur. J. Nutr. 2014, 53, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Ghandforoush-Sattari, M.; Mashayekhi, S.; Krishna, C.V.; Thompson, J.P.; Routledge, P.A. Pharmacokinetics of Oral Taurine in Healthy Volunteers. J. Amino Acids 2010, 2010, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.Q.; Thomas, T.R.; Ball, S.D. Effect of exercise timing on postprandial lipemia and HDL cholesterol subfractions. J. Appl. Physiol. 1998, 85, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Arena, R.; Riebe, D.; Pescatello, L.S. ACSM’s New Preparticipation Health Screening Recommendations from ACSM’s Guidelines for Exercise Testing and Prescription, Ninth Edition. Curr. Sports Med. Rep. 2013, 12, 215–217. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Physical Status: The Use and Interpretation of Anthropometry; WHO: Geneva, Switzerland, 1995; Available online: http://www.unu.edu/unupress/food/FNBv27n4_suppl_2_final.pdf (accessed on 19 May 2020).
- Jackson, A.S.; Pollock, M.L.; Ward, A. Generalized equations for predicting body density of women. Med. Sci. Sports Exerc. 1980, 12, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Deyl, Z.; Hyanek, J.; Horakova, M. Profiling of amino acids in body fluids and tissues by means of liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1986, 379, 177–250. [Google Scholar] [CrossRef]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Jeukendrup, A.; Saris, W.; Wagenmakers, A.J.M. Fat Metabolism During Exercise: A Review. Part I: Fatty Acid Mobilization and Muscle Metabolism. Int. J. Sports Med. 1998, 19, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Jeukendrup, A.; Saris, W.H.; Wagenmakers, A.J.M. Fat metabolism during exercise: A review--part II: Regulation of metabolism and the effects of training. Int. J. Sports Med. 1998, 19, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Jeukendrup, A.; Saris, W.H.; Wagenmakers, A.J.M. Fat metabolism during exercise: A review—Part III: Effects of nutritional interventions. Int. J. Sports Med. 1998, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Proeyen, K.; Szlufcik, K.; Nielens, H.; Deldicque, L.; Van Dyck, R.; Ramaekers, M.; Hespel, P. High-fat diet overrules the effects of training on fiber-specific intramyocellular lipid utilization during exercise. J. Appl. Physiol. 2011, 111, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulston, C.J.; Venables, M.; Mann, C.H.; Martin, C.; Philp, A.; Baar, K.; Jeukendrup, A.E. Training with Low Muscle Glycogen Enhances Fat Metabolism in Well-Trained Cyclists. Med. Sci. Sports Exerc. 2010, 42, 2046–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bock, K.; Richter, E.A.; Russell, A.; Eijnde, B.O.; Derave, W.; Ramaekers, M.; Koninckx, E.; Leger, B.; Verhaeghe, J.; Hespel, P. Exercise in the fasted state facilitates fibre type-specific intramyocellular lipid breakdown and stimulates glycogen resynthesis in humans. J. Physiol. 2005, 564, 649–660. [Google Scholar] [CrossRef]
- Lin, S.-C.; Hardie, D.G. AMPK: Sensing Glucose as well as Cellular Energy Status. Cell Metab. 2018, 27, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Guo, J.; Zhang, Y.; Zhang, J. The beneficial effects of taurine in preventing metabolic syndrome. Food Funct. 2016, 7, 1849–1863. [Google Scholar] [CrossRef]
- Tsuboyama-Kasaoka, N.; Shozawa, C.; Sano, K.; Kamei, Y.; Kasaoka, S.; Hosokawa, Y.; Ezaki, O. Taurine (2-Aminoethanesulfonic Acid) Deficiency Creates a Vicious Circle Promoting Obesity. Endocrinol. 2006, 147, 3276–3284. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.J.; Spriet, L.L. Regulation and role of hormone-sensitive lipase activity in human skeletal muscle. Proc. Nutr. Soc. 2004, 63, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.L.; Chambers, E.S.; Fletcher, G.; Wallis, G.A. Lipolytic Markers, Insulin and Resting Fat Oxidation are Associated with Maximal Fat Oxidation. Int. J. Sports Med. 2016, 37, 607–613. [Google Scholar] [CrossRef]
- Frayn, K.N. Fat as a fuel: Emerging understanding of the adipose tissue-skeletal muscle axis. Acta Physiol. 2010, 199, 509–518. [Google Scholar] [CrossRef]
- Van Loon, L.J.C.; Koopman, R.; Stegen, J.H.C.H.; Wagenmakers, A.J.M.; Keizer, H.A.; Saris, W.H.M. Intramyocellular lipids form an important substrate source during moderate intensity exercise in endurance-trained males in a fasted state. J. Physiol. 2003, 553, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Metab. 1993, 265, E380–E391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergman, B.C.; Brooks, G.A. Respiratory gas-exchange ratios during graded exercise in fed and fasted trained and untrained men. J. Appl. Physiol. 1999, 86, 479–487. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Mean ± SD |
|---|---|
| Age (years) | 24 ± 4 |
| Weight (kg) | 79 ± 10 |
| Height (m) | 1.80 ± 0.1 |
| BMI (kg/m²) | 23 ± 2 |
| Fat mass (%) | 14 ± 4 |
| Fat mass (kg) | 11 ± 4 |
| Fat-free mass (kg) | 68 ± 7 |
| VO2max (mL/kg/min) | 48 ± 8 |
| Glycerol (µm/dL) | Taurine (µmol/L) | |||||
|---|---|---|---|---|---|---|
| Groups | Pre | Post | p-Value | Pre | Post | p-Value |
| Placebo | 0.29 ± 0.01 a | 0.37 ± 0.08 | <0.001 | 16 ± 2 a | 16 ± 3 a | 0.573 |
| TAU 3 g | 0.31 ± 0.01 b | 0.41 ± 0.05 | <0.001 | 195 ± 57 b | 143 ± 37 b | 0.001 |
| TAU 6 g | 0.30 ± 0.03 a,b | 0.40 ± 0.04 | <0.001 | 411 ± 175 c | 278 ± 134 c | 0.017 |
| Groups | Pre | 30 min | 60 min | 15 min-Post | p-Value |
|---|---|---|---|---|---|
| Placebo | 65.2 ± 3.9c | 114.57 ± 5.2 a | 117.5 ± 2.9 a | 77.2 ± 7.5 b | p < 0.001 |
| TAU 3 g | 69.3 ± 2.4 c | 115.2 ± 4.0 a | 117.4 ± 6.3 a | 76.7 ± 5.2 b | p < 0.001 |
| TAU 6 g | 68.0 ± 8.4 c | 117.0 ± 2.1 a | 118.8 ± 6.1 a | 75.5 ± 5.3 b | p < 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, M.B.d.; Brandao, C.F.C.; Fassini, P.G.; Bianco, T.M.; Batitucci, G.; Galan, B.S.M.; Carvalho, F.G.D.; Vieira, T.S.; Ferriolli, E.; Marchini, J.S.; et al. Taurine Supplementation Increases Post-Exercise Lipid Oxidation at Moderate Intensity in Fasted Healthy Males. Nutrients 2020, 12, 1540. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051540
Carvalho MBd, Brandao CFC, Fassini PG, Bianco TM, Batitucci G, Galan BSM, Carvalho FGD, Vieira TS, Ferriolli E, Marchini JS, et al. Taurine Supplementation Increases Post-Exercise Lipid Oxidation at Moderate Intensity in Fasted Healthy Males. Nutrients. 2020; 12(5):1540. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051540
Chicago/Turabian StyleCarvalho, Milena Barbon de, Camila Fernanda Cunha Brandao, Priscila Giacomo Fassini, Thiago Mantello Bianco, Gabriela Batitucci, Bryan Steve Martinez Galan, Flávia Giolo De Carvalho, Tales Sambrano Vieira, Eduardo Ferriolli, Julio Sergio Marchini, and et al. 2020. "Taurine Supplementation Increases Post-Exercise Lipid Oxidation at Moderate Intensity in Fasted Healthy Males" Nutrients 12, no. 5: 1540. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051540