Interaction between Metabolic Genetic Risk Score and Dietary Fatty Acid Intake on Central Obesity in a Ghanaian Population

 , , and
, , and 
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Data Collection
2.3. Anthropometric Measurements
2.4. Physical Activity and Dietary Assessments
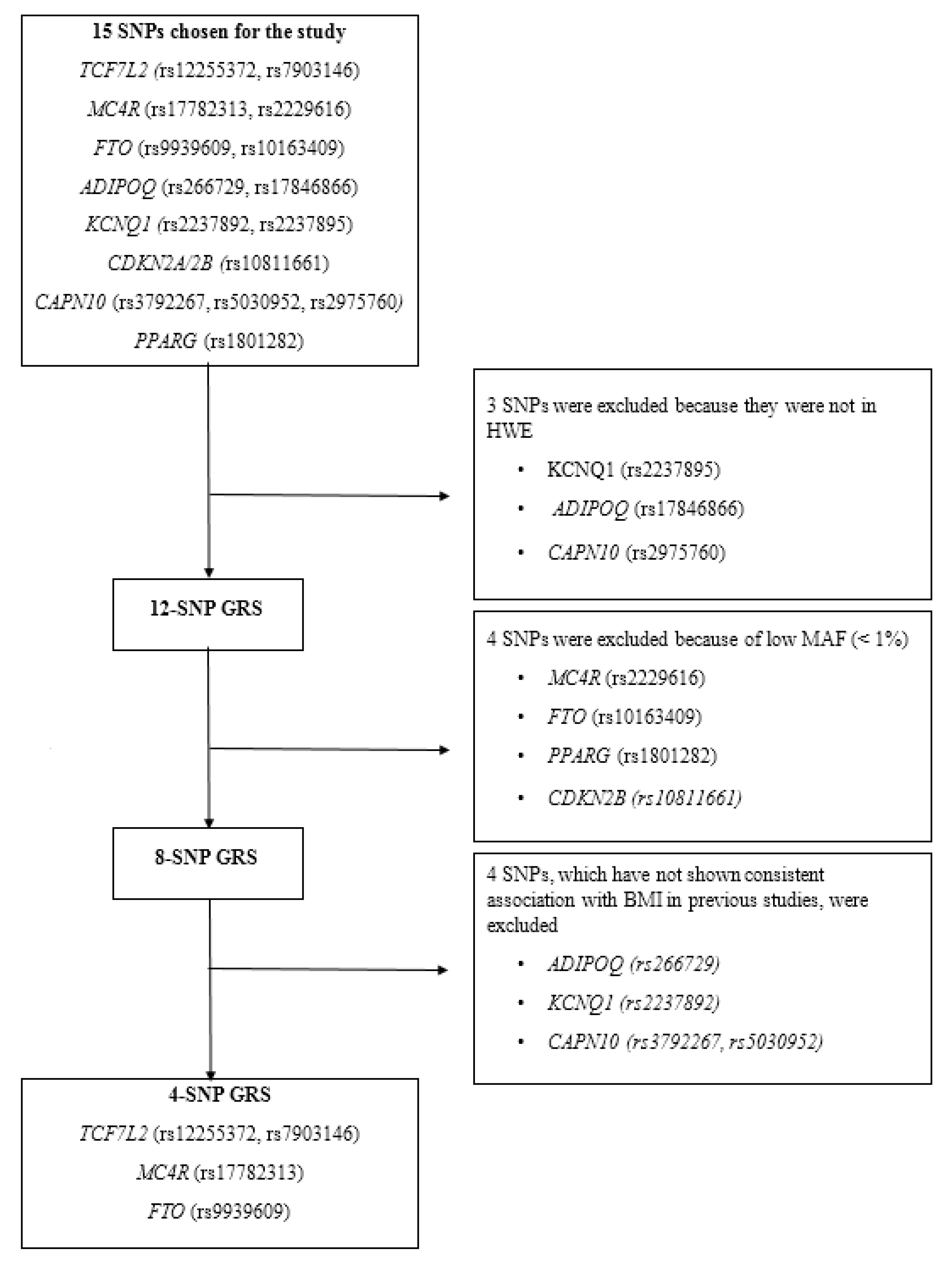
2.5. SNP Selection
2.6. Genotyping
2.7. Construction of the Metabolic GRSs
3. Results
3.1. Characteristics of the Study Participants
3.2. Effect of Metabolic GRSs on Obesity-Related Traits
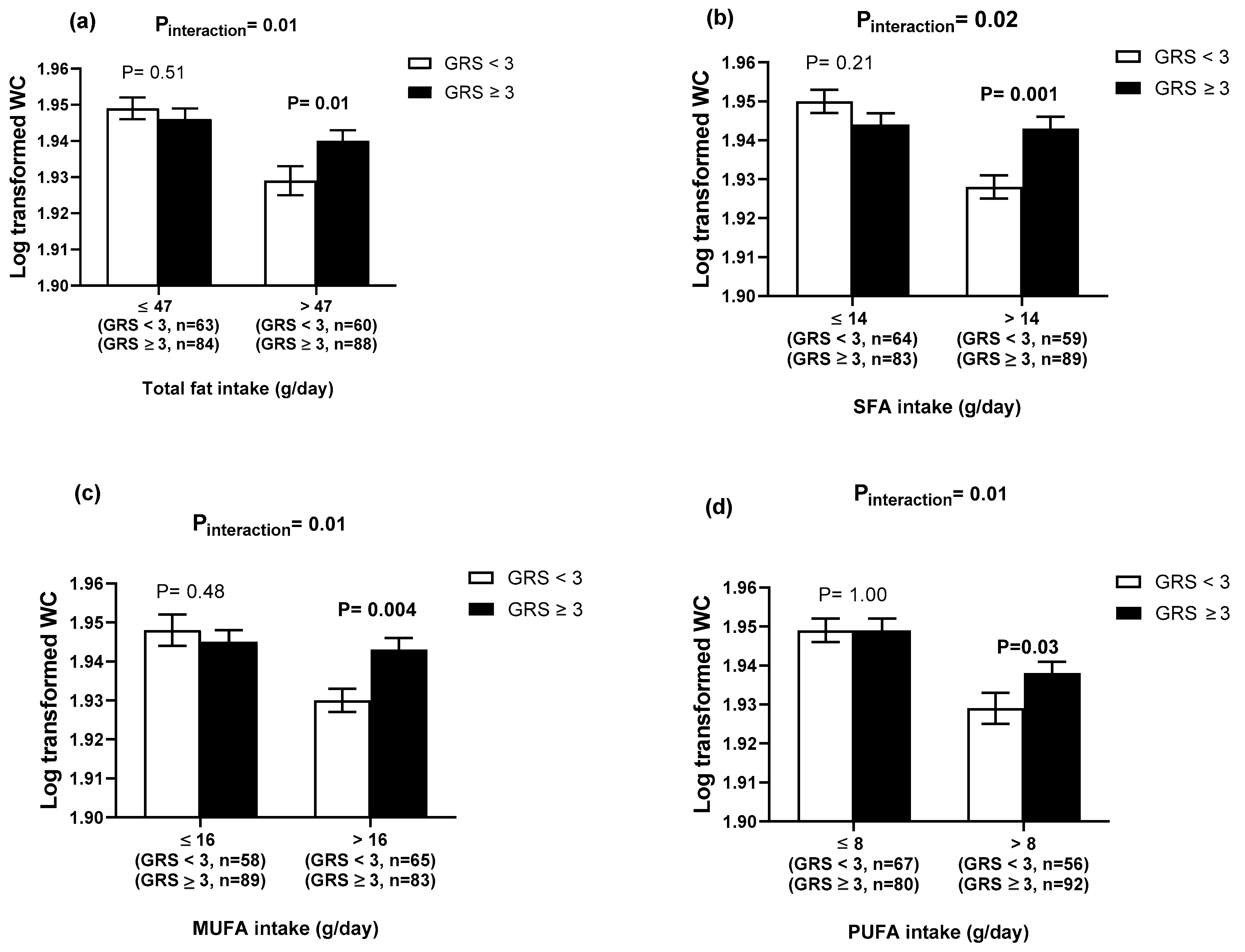
3.3. GRS–Lifestyle Interactions on Obesity-Related Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kelly, T.; Yang, W.; Chen, C.S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. (Lond.) 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Ofori-Asenso, R.; Agyeman, A.A.; Laar, A.; Boateng, D. Overweight and obesity epidemic in Ghana-a systematic review and meta-analysis. BMC Public Health 2016, 16, 1239. [Google Scholar] [CrossRef] [Green Version]
- Nicolaidis, S. Environment and obesity. Metabolism 2019, 100s, 153942. [Google Scholar] [CrossRef]
- Tilg, H.; Kaser, A. Gut microbiome, obesity, and metabolic dysfunction. J. Clin. Investig. 2011, 121, 2126–2132. [Google Scholar] [CrossRef]
- Froy, O. Metabolism and circadian rhythms--implications for obesity. Endocr. Rev. 2010, 31, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Egusquiza, R.J.; Blumberg, B. Environmental Obesogens and Their Impact on Susceptibility to Obesity: New Mechanisms and Chemicals. Endocrinology 2020, 161. [Google Scholar] [CrossRef] [Green Version]
- Herrera, B.M.; Keildson, S.; Lindgren, C.M. Genetics and epigenetics of obesity. Maturitas 2011, 69, 41–49. [Google Scholar] [CrossRef]
- Parsanathan, R.; Jain, S.K. Glutathione deficiency induces epigenetic alterations of vitamin D metabolism genes in the livers of high-fat diet-fed obese mice. Sci. Rep. 2019, 9, 14784. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabet. Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Owen, J.B. Genetic aspects of body composition. Nutrition 1999, 15, 609–613. [Google Scholar] [CrossRef]
- Berndt, S.I.; Gustafsson, S.; Magi, R.; Ganna, A.; Wheeler, E.; Feitosa, M.F.; Justice, A.E.; Monda, K.L.; Croteau-Chonka, D.C.; Day, F.R.; et al. Genome-wide meta-analysis identifies 11 new loci for anthropometric traits and provides insights into genetic architecture. Nat. Genet. 2013, 45, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Kubo, M.; Ohmiya, H.; Takahashi, A.; Kumasaka, N.; Hosono, N.; Maeda, S.; Wen, W.; Dorajoo, R.; Go, M.J.; et al. Common variants at CDKAL1 and KLF9 are associated with body mass index in east Asian populations. Nat. Genet. 2012, 44, 302–306. [Google Scholar] [CrossRef]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Lango Allen, H.; Lindgren, C.M.; Luan, J.; Magi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Cho, Y.S.; Zheng, W.; Dorajoo, R.; Kato, N.; Qi, L.; Chen, C.H.; Delahanty, R.J.; Okada, Y.; Tabara, Y.; et al. Meta-analysis identifies common variants associated with body mass index in east Asians. Nat. Genet. 2012, 44, 307–311. [Google Scholar] [CrossRef]
- Willer, C.J.; Li, Y.; Abecasis, G.R. METAL: Fast and efficient meta-analysis of genomewide association scans. Bioinformatics 2010, 26, 2190–2191. [Google Scholar] [CrossRef]
- Winkler, T.W.; Justice, A.E.; Graff, M.; Barata, L.; Feitosa, M.F.; Chu, S.; Czajkowski, J.; Esko, T.; Fall, T.; Kilpelainen, T.O.; et al. Correction: The Influence of Age and Sex on Genetic Associations with Adult Body Size and Shape: A Large-Scale Genome-Wide Interaction Study. PLoS Genet. 2016, 12, e1006166. [Google Scholar] [CrossRef]
- Fesinmeyer, M.D.; North, K.E.; Ritchie, M.D.; Lim, U.; Franceschini, N.; Wilkens, L.R.; Gross, M.D.; Buzkova, P.; Glenn, K.; Quibrera, P.M.; et al. Genetic risk factors for BMI and obesity in an ethnically diverse population: Results from the population architecture using genomics and epidemiology (PAGE) study. Obesity (Silver Spring) 2013, 21, 835–846. [Google Scholar] [CrossRef]
- Hester, J.M.; Wing, M.R.; Li, J.; Palmer, N.D.; Xu, J.; Hicks, P.J.; Roh, B.H.; Norris, J.M.; Wagenknecht, L.E.; Langefeld, C.D.; et al. Implication of European-derived adiposity loci in African Americans. Int. J. Obes. (Lond.) 2012, 36, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Rotimi, C.; Abayomi, A.; Abimiku, A.; Adabayeri, V.M.; Adebamowo, C.; Adebiyi, E.; Ademola, A.D.; Adeyemo, A.; Adu, D.; Affolabi, D.; et al. Research capacity. Enabling the genomic revolution in Africa. Science 2014, 344, 1346–1348. [Google Scholar]
- Rotimi, C.N.; Bentley, A.R.; Doumatey, A.P.; Chen, G.; Shriner, D.; Adeyemo, A. The genomic landscape of African populations in health and disease. Hum. Mol. Genet. 2017, 26, 225–236. [Google Scholar] [CrossRef]
- Adeyemo, A.; Chen, G.; Zhou, J.; Shriner, D.; Doumatey, A.; Huang, H.; Rotimi, C. FTO genetic variation and association with obesity in West Africans and African Americans. Diabetes 2010, 59, 1549–1554. [Google Scholar] [CrossRef] [Green Version]
- Vimaleswaran, K.S.; Loos, R.J. Progress in the genetics of common obesity and type 2 diabetes. Expert Rev. Mol. Med. 2010, 12, e7. [Google Scholar] [CrossRef]
- Belsky, D.W.; Moffitt, T.E.; Sugden, K.; Williams, B.; Houts, R.; McCarthy, J.; Caspi, A. Development and evaluation of a genetic risk score for obesity. Biodemogr. Soc. Biol. 2013, 59, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.F.; Breen, G.; Czamara, D.; Corre, T.; Wolf, C.; Kloiber, S.; Bergmann, S.; Craddock, N.; Gill, M.; Holsboer, F.; et al. A genetic risk score combining 32 SNPs is associated with body mass index and improves obesity prediction in people with major depressive disorder. BMC Med. 2015, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, J.H.; Luan, J.; Luben, R.N.; Rodwell, S.A.; Khaw, K.T.; Ong, K.K.; Wareham, N.J.; Loos, R.J. Cumulative effects and predictive value of common obesity-susceptibility variants identified by genome-wide association studies. Am. J. Clin. Nutr. 2010, 91, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.E.; Maes, H.H.; Holmans, P.; Sanders, A.R.; Levinson, D.F.; Shi, J.; Kendler, K.S.; Gejman, P.V.; Webb, B.T. Genetic risk sum score comprised of common polygenic variation is associated with body mass index. Hum. Genet. 2011, 129, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Sandholt, C.H.; Sparso, T.; Grarup, N.; Albrechtsen, A.; Almind, K.; Hansen, L.; Toft, U.; Jorgensen, T.; Hansen, T.; Pedersen, O. Combined analyses of 20 common obesity susceptibility variants. Diabetes 2010, 59, 1667–1673. [Google Scholar] [CrossRef] [Green Version]
- Fulford, A.J.; Ong, K.K.; Elks, C.E.; Prentice, A.M.; Hennig, B.J. Progressive influence of body mass index-associated genetic markers in rural Gambians. J. Med. Genet. 2015, 52, 375–380. [Google Scholar] [CrossRef]
- Hennig, B.J.; Fulford, A.J.; Sirugo, G.; Rayco-Solon, P.; Hattersley, A.T.; Frayling, T.M.; Prentice, A.M. FTO gene variation and measures of body mass in an African population. BMC Med. Genet. 2009, 10, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badsi, M.N.; Mediene-Benchekor, S.; Ouhaibi-Djellouli, H.; Lardjam-Hetraf, S.A.; Boulenouar, H.; Meroufel, D.N.; Hermant, X.; Hamani-Medjaoui, I.; Saidi-Mehtar, N.; Amouyel, P.; et al. Combined effect of established BMI loci on obesity-related traits in an Algerian population sample. BMC Genet. 2014, 15, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munthali, R.J.; Sahibdeen, V.; Kagura, J.; Hendry, L.M.; Norris, S.A.; Ong, K.K.; Day, F.R.; Lombard, Z. Genetic risk score for adult body mass index associations with childhood and adolescent weight gain in an African population. Genes Nutr. 2018, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Huang, J.; Rose, L.M.; Jensen, M.K.; Liang, L.; Curhan, G.C.; Pasquale, L.R.; Wiggs, J.L.; et al. Fried food consumption, genetic risk, and body mass index: Gene-diet interaction analysis in three US cohort studies. BMJ 2014, 348, g1610. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Jensen, M.K.; Curhan, G.C.; Pasquale, L.R.; Ridker, P.M.; Hunter, D.J.; Willett, W.C.; Rimm, E.B.; et al. Sugar-sweetened beverages and genetic risk of obesity. N. Engl. J. Med. 2012, 367, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Li, Y.; Chomistek, A.K.; Kang, J.H.; Curhan, G.C.; Pasquale, L.R.; Willett, W.C.; Rimm, E.B.; Hu, F.B.; Qi, L. Television watching, leisure time physical activity, and the genetic predisposition in relation to body mass index in women and men. Circulation 2012, 126, 1821–1827. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, J.H.; Luan, J.; Ekelund, U.; Luben, R.N.; Khaw, K.T.; Wareham, N.J.; Loos, R.J. Physical activity attenuates the genetic predisposition to obesity in 20,000 men and women from EPIC-Norfolk prospective population study. PLoS Med. 2010, 7, e1000332. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Heianza, Y.; Sun, D.; Huang, T.; Ma, W.; Rimm, E.B.; Manson, J.E.; Hu, F.B.; Willett, W.; Qi, L. Improving adherence to healthy dietary patterns, genetic risk, and long term weight gain: Gene-diet interaction analysis in two prospective cohort studies. BMJ 2018, 360, 693. [Google Scholar] [CrossRef] [Green Version]
- Kilpelainen, T.O.; Qi, L.; Brage, S.; Sharp, S.J.; Sonestedt, E.; Demerath, E.; Ahmad, T.; Mora, S.; Kaakinen, M.; Sandholt, C.H.; et al. Physical activity attenuates the influence of FTO variants on obesity risk: A meta-analysis of 218,166 adults and 19,268 children. PLoS Med. 2011, 8, e1001116. [Google Scholar] [CrossRef]
- Vimaleswaran, K.S. Gene–nutrient interactions on metabolic diseases: Findings from the GeNuIne collaboration. Nutr. Bull. 2017, 42, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Vimaleswaran, K.S. A nutrigenetics approach to study the impact of genetic and lifestyle factors on cardiometabolic traits in various ethnic groups: Findings from the GeNuIne Collaboration. Proc. Nutr. Soc. 2020, 79, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Global Physical Activity Surveillance. Available online: https://www.who.int/ncds/surveillance/steps/GPAQ/en/ (accessed on 19 March 2020).
- Janssens, A.C.; Moonesinghe, R.; Yang, Q.; Steyerberg, E.W.; van Duijn, C.M.; Khoury, M.J. The impact of genotype frequencies on the clinical validity of genomic profiling for predicting common chronic diseases. Genet. Med. 2007, 9, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzi, M.D.; Nasr, F.A.; Alanazi, M.S.; Alamri, A.; Turjoman, A.A.; Moustafa, A.S.; Alfadda, A.A.; Pathan, A.A.; Parine, N.R. Association between FTO, MC4R, SLC30A8, and KCNQ1 gene variants and type 2 diabetes in Saudi population. Genet. Mol. Res. 2014, 13, 10194–10203. [Google Scholar] [CrossRef] [PubMed]
- Holmkvist, J.; Banasik, K.; Andersen, G.; Unoki, H.; Jensen, T.S.; Pisinger, C.; Borch-Johnsen, K.; Sandbaek, A.; Lauritzen, T.; Brunak, S.; et al. The type 2 diabetes associated minor allele of rs2237895 KCNQ1 associates with reduced insulin release following an oral glucose load. PLoS ONE 2009, 4, e5872. [Google Scholar] [CrossRef]
- Hsiao, T.J.; Lin, E. A Validation Study of Adiponectin rs266729 Gene Variant with Type 2 Diabetes, Obesity, and Metabolic Phenotypes in a Taiwanese Population. Biochem. Genet. 2016, 54, 830–841. [Google Scholar] [CrossRef]
- Kommoju, U.J.; Maruda, J.; Kadarkarai Samy, S.; Irgam, K.; Kotla, J.P.; Reddy, B.M. Association of IRS1, CAPN10, and PPARG gene polymorphisms with type 2 diabetes mellitus in the high-risk population of Hyderabad, India. J. Diabetes 2014, 6, 564–573. [Google Scholar] [CrossRef]
- Lu, J.F.; Zhou, Y.; Huang, G.H.; Jiang, H.X.; Hu, B.L.; Qin, S.Y. Association of ADIPOQ polymorphisms with obesity risk: A meta-analysis. Hum. Immunol. 2014, 75, 1062–1068. [Google Scholar] [CrossRef]
- Martinez-Gomez, L.E.; Cruz, M.; Martinez-Nava, G.A.; Madrid-Marina, V.; Parra, E.; Garcia-Mena, J.; Espinoza-Rojo, M.; Estrada-Velasco, B.I.; Piza-Roman, L.F.; Aguilera, P.; et al. A replication study of the IRS1, CAPN10, TCF7L2, and PPARG gene polymorphisms associated with type 2 diabetes in two different populations of Mexico. Ann. Hum. Genet. 2011, 75, 612–620. [Google Scholar] [CrossRef]
- Galbete, C.; Nicolaou, M.; Meeks, K.A.; de-Graft Aikins, A.; Addo, J.; Amoah, S.K.; Smeeth, L.; Owusu-Dabo, E.; Klipstein-Grobusch, K.; Bahendeka, S.; et al. Food consumption, nutrient intake, and dietary patterns in Ghanaian migrants in Europe and their compatriots in Ghana. Food Nutr. Res. 2017, 61, 1341809. [Google Scholar] [CrossRef] [Green Version]
- Logan, M.; Van der Merwe, M.T.; Dodgen, T.M.; Myburgh, R.; Eloff, A.; Alessandrini, M.; Pepper, M.S. Allelic variants of the Melanocortin 4 receptor (MC4R) gene in a South African study group. Mol. Genet. Genom. Med. 2016, 4, 68–76. [Google Scholar] [CrossRef]
- Sahibdeen, V.; Crowther, N.J.; Soodyall, H.; Hendry, L.M.; Munthali, R.J.; Hazelhurst, S.; Choudhury, A.; Norris, S.A.; Ramsay, M.; Lombard, Z. Genetic variants in SEC16B are associated with body composition in black South Africans. Nutr. Diabetes 2018, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemo, A.A.; Tekola-Ayele, F.; Doumatey, A.P.; Bentley, A.R.; Chen, G.; Huang, H.; Zhou, J.; Shriner, D.; Fasanmade, O.; Okafor, G.; et al. Evaluation of Genome Wide Association Study Associated Type 2 Diabetes Susceptibility Loci in Sub Saharan Africans. Front. Genet. 2015, 6, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Sun, M.; Adeyemo, A.; Pirie, F.; Carstensen, T.; Pomilla, C.; Doumatey, A.P.; Chen, G.; Young, E.H.; Sandhu, M.; et al. Genome-wide association study of type 2 diabetes in Africa. Diabetologia 2019, 62, 1204–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Schumacher, F.; Lim, U.; Hindorff, L.A.; Haessler, J.; Buyske, S.; Carlson, C.S.; Rosse, S.; Buzkova, P.; Fornage, M.; et al. Fine Mapping and Identification of BMI Loci in African Americans. Am. J. Hum. Genet. 2013, 93, 661–671. [Google Scholar] [CrossRef] [Green Version]
- Shriner, D.; Adeyemo, A.; Gerry, N.P.; Herbert, A.; Chen, G.; Doumatey, A.; Huang, H.; Zhou, J.; Christman, M.F.; Rotimi, C.N. Transferability and fine-mapping of genome-wide associated loci for adult height across human populations. PLoS ONE 2009, 4, e8398. [Google Scholar] [CrossRef] [Green Version]
- Yako, Y.Y.; Echouffo-Tcheugui, J.B.; Balti, E.V.; Matsha, T.E.; Sobngwi, E.; Erasmus, R.T.; Kengne, A.P. Genetic association studies of obesity in Africa: A systematic review. Obes. Rev. 2015, 16, 259–272. [Google Scholar] [CrossRef]
- Chen, G.; Doumatey, A.P.; Zhou, J.; Lei, L.; Bentley, A.R.; Tekola-Ayele, F.; Adebamowo, S.N.; Baker, J.L.; Fasanmade, O.; Okafor, G.; et al. Genome-wide analysis identifies an african-specific variant in SEMA4D associated with body mass index. Obesity (Silver Spring) 2017, 25, 794–800. [Google Scholar] [CrossRef] [Green Version]
- Yako, Y.Y.; Madubedube, J.H.; Kengne, A.P.; Erasmus, R.T.; Pillay, T.S.; Matsha, T.E. Contribution of ENPP1, TCF7L2, and FTO polymorphisms to type 2 diabetes in mixed ancestry ethnic population of South Africa. Afr. Health Sci. 2015, 15, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Monda, K.L.; Chen, G.K.; Taylor, K.C.; Palmer, C.; Edwards, T.L.; Lange, L.A.; Ng, M.C.; Adeyemo, A.A.; Allison, M.A.; Bielak, L.F.; et al. A meta-analysis identifies new loci associated with body mass index in individuals of African ancestry. Nat. Genet. 2013, 45, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.C.; Tishkoff, S.A. African genetic diversity: Implications for human demographic history, modern human origins, and complex disease mapping. Annu. Rev. Genom. Hum. Genet. 2008, 9, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Schleinitz, D.; Bottcher, Y.; Bluher, M.; Kovacs, P. The genetics of fat distribution. Diabetologia 2014, 57, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rimm, E.B.; Stampfer, M.J.; Willett, W.C.; Hu, F.B. Comparison of abdominal adiposity and overall obesity in predicting risk of type 2 diabetes among men. Am. J. Clin. Nutr. 2005, 81, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Corella, D.; Arnett, D.K.; Tucker, K.L.; Kabagambe, E.K.; Tsai, M.; Parnell, L.D.; Lai, C.Q.; Lee, Y.C.; Warodomwichit, D.; Hopkins, P.N.; et al. A high intake of saturated fatty acids strengthens the association between the fat mass and obesity-associated gene and BMI. J. Nutr. 2011, 141, 2219–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, C.M.; Kesse-Guyot, E.; McManus, R.; Hercberg, S.; Lairon, D.; Planells, R.; Roche, H.M. High dietary saturated fat intake accentuates obesity risk associated with the fat mass and obesity-associated gene in adults. J. Nutr. 2012, 142, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Moleres, A.; Ochoa, M.C.; Rendo-Urteaga, T.; Martinez-Gonzalez, M.A.; Azcona San Julian, M.C.; Martinez, J.A.; Marti, A. Dietary fatty acid distribution modifies obesity risk linked to the rs9939609 polymorphism of the fat mass and obesity-associated gene in a Spanish case-control study of children. Br. J. Nutr. 2012, 107, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Sonestedt, E.; Roos, C.; Gullberg, B.; Ericson, U.; Wirfalt, E.; Orho-Melander, M. Fat and carbohydrate intake modify the association between genetic variation in the FTO genotype and obesity. Am. J. Clin. Nutr. 2009, 90, 1418–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.; Kilpelainen, T.O.; Downer, M.K.; Tanaka, T.; Smith, C.E.; Sluijs, I.; Sonestedt, E.; Chu, A.Y.; Renstrom, F.; Lin, X.; et al. FTO genetic variants, dietary intake and body mass index: Insights from 177,330 individuals. Hum. Mol. Genet. 2014, 23, 6961–6972. [Google Scholar] [CrossRef]
- De Luis, D.A.; Aller, R.; Izaola, O.; de la Fuente, B.; Conde, R.; Sagrado, M.G.; Primo, D. Evaluation of weight loss and adipocytokines levels after two hypocaloric diets with different macronutrient distribution in obese subjects with rs9939609 gene variant. Diabetes Metab. Res. Rev. 2012, 28, 663–668. [Google Scholar] [CrossRef]
- Bodhini, D.; Gaal, S.; Shatwan, I.; Ramya, K.; Ellahi, B.; Surendran, S.; Sudha, V.; Anjana, M.R.; Mohan, V.; Lovegrove, J.A.; et al. Interaction between TCF7L2 polymorphism and dietary fat intake on high density lipoprotein cholesterol. PLoS ONE 2017, 12, e0188382. [Google Scholar]
- Casas-Agustench, P.; Arnett, D.K.; Smith, C.E.; Lai, C.Q.; Parnell, L.D.; Borecki, I.B.; Frazier-Wood, A.C.; Allison, M.; Chen, Y.D.; Taylor, K.D.; et al. Saturated fat intake modulates the association between an obesity genetic risk score and body mass index in two US populations. J. Acad. Nutr. Diet. 2014, 114, 1954–1966. [Google Scholar] [CrossRef] [Green Version]
- Celis-Morales, C.A.; Lyall, D.M.; Gray, S.R.; Steell, L.; Anderson, J.; Iliodromiti, S.; Welsh, P.; Guo, Y.; Petermann, F.; Mackay, D.F.; et al. Dietary fat and total energy intake modifies the association of genetic profile risk score on obesity: Evidence from 48 170 UK Biobank participants. Int. J. Obes. (Lond.) 2017, 41, 1761–1768. [Google Scholar]
- Rukh, G.; Sonestedt, E.; Melander, O.; Hedblad, B.; Wirfalt, E.; Ericson, U.; Orho-Melander, M. Genetic susceptibility to obesity and diet intakes: Association and interaction analyses in the Malmo Diet and Cancer Study. Genes Nutr. 2013, 8, 535–547. [Google Scholar] [PubMed] [Green Version]
- Bray, G.A.; Paeratakul, S.; Popkin, B.M. Dietary fat and obesity: A review of animal, clinical and epidemiological studies. Physiol. Behav. 2004, 83, 549–555. [Google Scholar] [PubMed]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 Polyunsaturated Fatty Acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [PubMed] [Green Version]
- Iso, H.; Rexrode, K.M.; Stampfer, M.J.; Manson, J.E.; Colditz, G.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Intake of fish and omega-3 fatty acids and risk of stroke in women. JAMA 2001, 285, 304–312. [Google Scholar] [PubMed] [Green Version]
- He, K.; Rimm, E.B.; Merchant, A.; Rosner, B.A.; Stampfer, M.J.; Willett, W.C.; Ascherio, A. Fish consumption and risk of stroke in men. JAMA 2002, 288, 3130–3136. [Google Scholar]
- Kabir, M.; Skurnik, G.; Naour, N.; Pechtner, V.; Meugnier, E.; Rome, S.; Quignard-Boulange, A.; Vidal, H.; Slama, G.; Clement, K.; et al. Treatment for 2 mo with n 3 polyunsaturated fatty acids reduces adiposity and some atherogenic factors but does not improve insulin sensitivity in women with type 2 diabetes: A randomized controlled study. Am. J. Clin. Nutr. 2007, 86, 1670–1679. [Google Scholar]
- Thorsdottir, I.; Tomasson, H.; Gunnarsdottir, I.; Gisladottir, E.; Kiely, M.; Parra, M.D.; Bandarra, N.M.; Schaafsma, G.; Martinez, J.A. Randomized trial of weight-loss-diets for young adults varying in fish and fish oil content. Int. J. Obes. (Lond.) 2007, 31, 1560–1566. [Google Scholar]
- Weech, M.; Vafeiadou, K.; Hasaj, M.; Todd, S.; Yaqoob, P.; Jackson, K.G.; Lovegrove, J.A. Development of a food-exchange model to replace saturated fat with MUFAs and n-6 PUFAs in adults at moderate cardiovascular risk. J. Nutr. 2014, 144, 846–855. [Google Scholar]
- Kastorini, C.M.; Milionis, H.J.; Esposito, K.; Giugliano, D.; Goudevenos, J.A.; Panagiotakos, D.B. The effect of Mediterranean diet on metabolic syndrome and its components: A meta-analysis of 50 studies and 534,906 individuals. J. Am. Coll. Cardiol. 2011, 57, 1299–1313. [Google Scholar]
- Khaw, K.T.; Sharp, S.J.; Finikarides, L.; Afzal, I.; Lentjes, M.; Luben, R.; Forouhi, N.G. Randomised trial of coconut oil, olive oil or butter on blood lipids and other cardiovascular risk factors in healthy men and women. BMJ Open 2018, 8, e020167. [Google Scholar] [PubMed] [Green Version]
- Heianza, Y.; Qi, L. Gene-Diet Interaction and Precision Nutrition in Obesity. Int. J. Mol. Sci. 2017, 18, 787. [Google Scholar]
- Healthy Diet. Available online: https://www.who.int/nutrition/publications/nutrientrequirements/healthydiet_factsheet/en/. (accessed on 21 January 2020).
- Interim Summary of Conclusions and Dietary Recommendations on Total Fat & Fatty Acids. Available online: https://www.who.int/nutrition/topics/FFA_summary_rec_conclusion.pdf. (accessed on 21 January 2020).
- Micha, R.; Khatibzadeh, S.; Shi, P.; Fahimi, S.; Lim, S.; Andrews, K.G.; Engell, R.E.; Powles, J.; Ezzati, M.; Mozaffarian, D. Global, regional, and national consumption levels of dietary fats and oils in 1990 and 2010: A systematic analysis including 266 country-specific nutrition surveys. BMJ 2014, 348, 2272. [Google Scholar]
- Mertens, E.; Kuijsten, A.; Geleijnse, J.M.; Boshuizen, H.C.; Feskens, E.J.M.; Van’t Veer, P. FFQ versus repeated 24-h recalls for estimating diet-related environmental impact. Nutr. J. 2019, 18, 2. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Total (N = 302) | Non-Obese * (N = 126) | Overweight/Obese ** (N = 176) | p Value *** | |
|---|---|---|---|---|
| Age (years) | 38.17 ± 9.64 | 35.96 ± 9.55 | 39.75 ± 9.42 | 0.001 |
| BMI (kg/m2) | 26.63 ± 4.99 | 22.01 ± 1.79 | 29.95 ± 3.75 | <0.001 |
| WC (cm) | 88.48 ± 12.41 | 77.99 ± 7.13 | 96.00 ± 9.61 | <0.001 |
| WHR | 1.45 ± 6.96 | 1.55 ± 7.76 | 1.38 ± 6.34 | 0.84 |
| Visceral fat (%) | 8.02 ± 7.39 | 6.49 ± 10.97 | 9.12 ± 2.26 | 0.01 |
| Body fat (%) | 33.12 ± 13.90 | 22.05 ± 12.47 | 41.05 ± 8.36 | <0.001 |
| Total energy intake (%) | 1647.93 ± 685.83 | 1772.17 ± 723.85 | 1558.99 ± 644.75 | 0.008 |
| Protein intake (g/day) | 53.24 ± 23.73 | 57.38 ± 24.52 | 50.28 ± 22.76 | 0.01 |
| Total fat intake (g/day) | 51.17 ± 26.94 | 55.00 ± 29.29 | 48.42 ± 24.85 | 0.04 |
| Carbohydrates intake (g/day) | 239.03 ± 95.84 | 259.44 ± 104.01 | 224.42 ± 86.94 | 0.002 |
| Fibre intake (g/day) | 21.31 ± 10.84 | 23.19 ± 11.44 | 19.96 ± 10.21 | 0.01 |
| Total SFA intake (g/day) | 16.23 ± 10.36 | 17.41 ± 11.29 | 15.39 ± 9.58 | 0.10 |
| Total MUFA intake (g/day) | 18.08 ± 10.49 | 19.63 ± 11.30 | 16.96 ± 9.74 | 0.03 |
| Total PUFA intake (g/day) | 9.12 ± 5.03 | 10.20 ± 5.56 | 8.35 ± 4.47 | 0.002 |
| GRS < 3 Risk Alleles (N = 123) | GRS ≥ 3 Risk Allele (N = 172) | * p Value | |
|---|---|---|---|
| BMI (kg/m2) | 26.13 ± 0.45 | 26.85 ± 0.37 | 0.24 |
| WC (cm) | 87.13 ± 1.15 | 89.14 ± 0.92 | 0.19 |
| WHR | 2.27 ± 0.98 | 0.88 ± 0.01 | 0.18 |
| Visceral fat (%) | 7.89 ± 0.71 | 8.08 ± 0.55 | 0.43 |
| Body fat (%) | 31.75 ± 1.32 | 33.87 ± 1.02 | 0.15 |
| Protein (g/day) | Carbohydrate (g/day) | Fibre (g/day) | Fat (g/day) | SFA (g/day) | MUFA (g/day) | PUFA (g/day) | Physical Activity | |
|---|---|---|---|---|---|---|---|---|
| BMI (kg/m2) | 0.45 | 0.22 | 0.12 | 0.15 | - | - | - | 0.76 |
| WC (cm) | 0.08 | 0.21 | 0.41 | 0.01 | 0.02 | 0.01 | 0.01 | 0.24 |
| WHR | 0.82 | 0.88 | 0.49 | 0.80 | - | - | - | 0.002 |
| Visceral fat (%) | 0.50 | 0.35 | 0.32 | 0.38 | - | - | - | 0.93 |
| Body fat (%) | 0.46 | 0.11 | 0.04 | 0.75 | - | - | - | 0.60 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsulami, S.; Nyakotey, D.A.; Dudek, K.; Bawah, A.-M.; Lovegrove, J.A.; Annan, R.A.; Ellahi, B.; Vimaleswaran, K.S. Interaction between Metabolic Genetic Risk Score and Dietary Fatty Acid Intake on Central Obesity in a Ghanaian Population. Nutrients 2020, 12, 1906. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071906
Alsulami S, Nyakotey DA, Dudek K, Bawah A-M, Lovegrove JA, Annan RA, Ellahi B, Vimaleswaran KS. Interaction between Metabolic Genetic Risk Score and Dietary Fatty Acid Intake on Central Obesity in a Ghanaian Population. Nutrients. 2020; 12(7):1906. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071906
Chicago/Turabian StyleAlsulami, Sooad, David A. Nyakotey, Kamila Dudek, Abdul-Malik Bawah, Julie A. Lovegrove, Reginald A. Annan, Basma Ellahi, and Karani Santhanakrishnan Vimaleswaran. 2020. "Interaction between Metabolic Genetic Risk Score and Dietary Fatty Acid Intake on Central Obesity in a Ghanaian Population" Nutrients 12, no. 7: 1906. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071906