Sleep and Microbiome in Psychiatric Diseases

 , , , ,
, , , ,
Abstract
:1. Introduction
1.1. Background
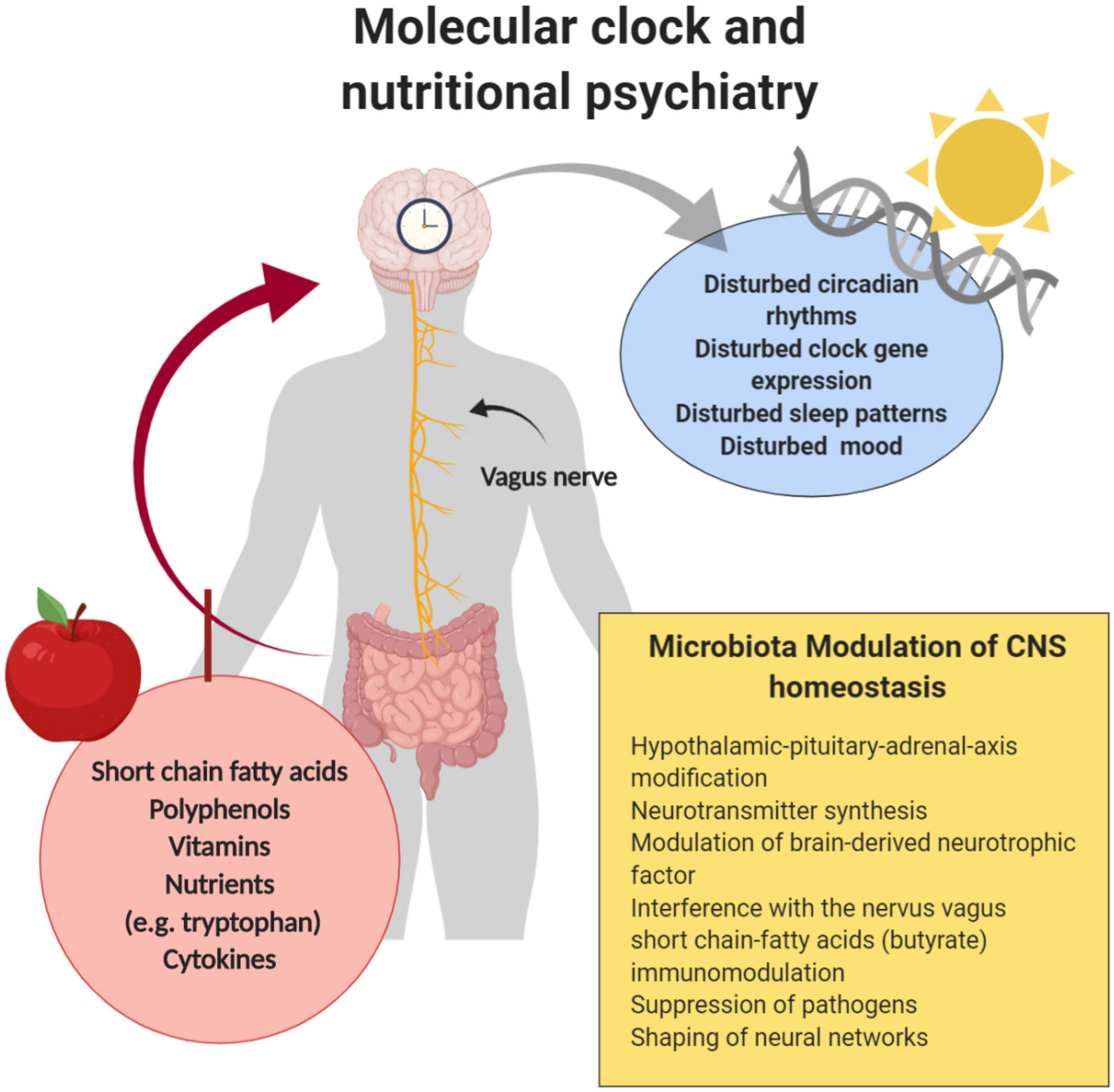
1.2. The Molecular 24-h Clock
1.3. Gut Microbiome and Sleep
- (1)
- We aimed to review all cross-sectional studies investigating sleep and the gut microbiome in individuals with psychiatric disorders.
- (2)
- Furthermore, we aimed to systematically review all interventional studies, which measured sleep and the gut microbiome in individuals with psychiatric disorders.
- (3)
- Additionally, we aimed to narratively review the association between circadian rhythms, sleep and the gut microbiome in patients with psychiatric disorders and discuss the clinical implications of such associations, with particular emphasis on mechanisms of regulation of the MGBA and circadian rhythms in order to improve sleep (such as nutrition, probiotics and metabolites, psychotherapy and chronotherapy).
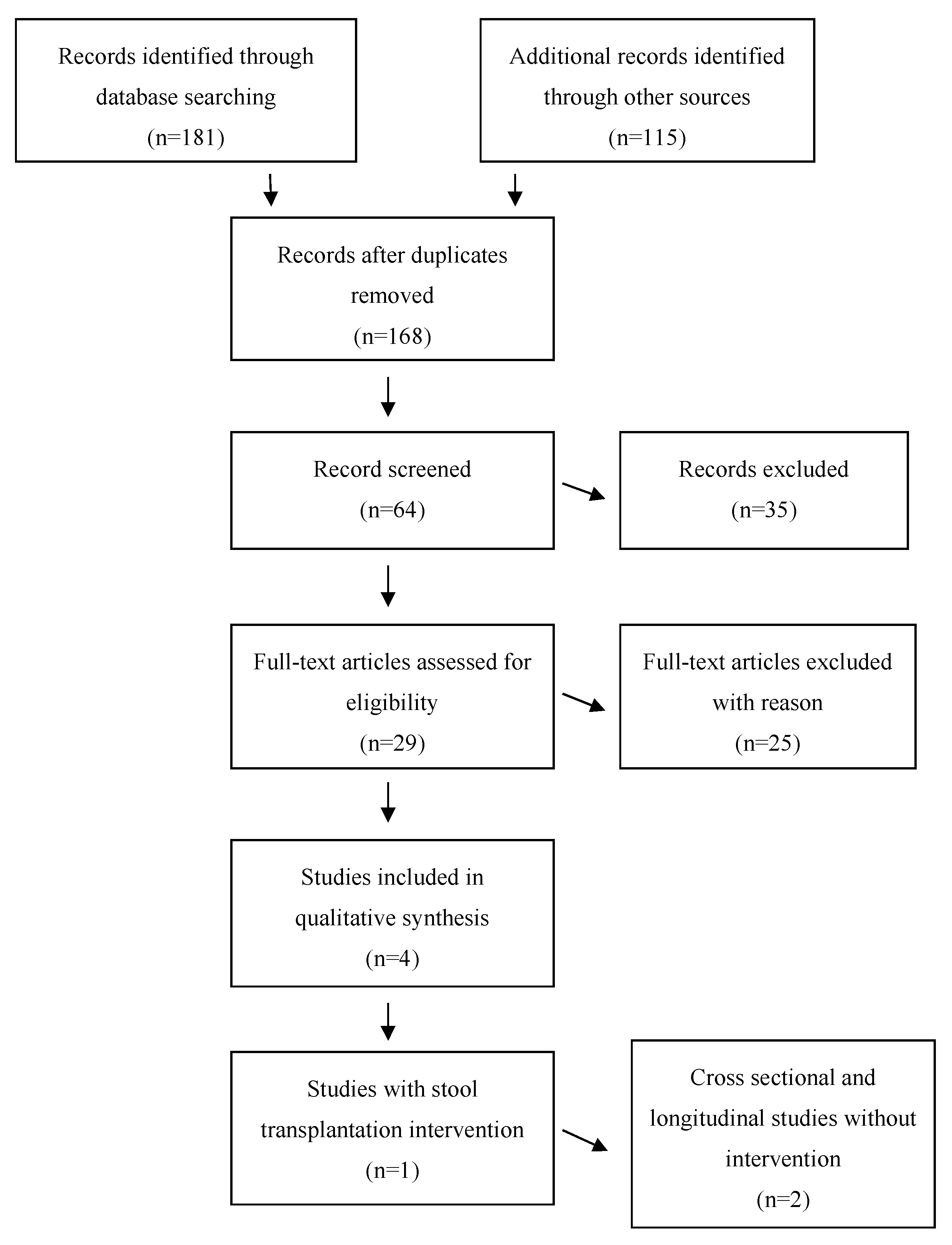
2. Methods
3. Results
4. Discussion
4.1. Clock Genes, Diet and Sleep Disturbances Affect Physiological Oscillations of the Gut Microbiome
4.2. Gut Microbiome Diversity Is Associated with Sleep Physiology in Healthy Individuals
4.3. Clinical Implications
4.3.1. The Interplay of Dietary and Nutritive Composition, Microbiota, Mental Health and Sleep Quality
4.3.2. Dietary Habits and Sleep
- (1)
- Sleep habits affect how much and what we eat
- (2)
- Dietary patterns and habits affect how we sleep
4.3.3. Psychotropic Medication Affecting Circadian Rhythms
4.3.4. Psychological and Psychotherapeutic Interventions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stickgold, R.; Walker, M.P. Sleep-Dependent Memory Consolidation and Reconsolidation. Sleep Med. 2007, 8, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertenstein, E.; Feige, B.; Gmeiner, T.; Kienzler, C.; Spiegelhalder, K.; Johann, A.; Jansson-Frojmark, M.; Palagini, L.; Rucker, G.; Riemann, D.; et al. Insomnia as a Predictor of Mental Disorders: A Systematic Review and Meta-Analysis. Sleep Med. Rev. 2019, 43, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lyall, L.M.; Wyse, C.A.; Graham, N.; Ferguson, A.; Lyall, D.M.; Cullen, B.; Celis Morales, C.A.; Biello, S.M.; Mackay, D.; Ward, J.; et al. Association of Disrupted Circadian Rhythmicity with Mood Disorders, Subjective Wellbeing, and Cognitive Function: A Cross-Sectional Study of 91 105 Participants from the UK Biobank. Lancet Psychiatry 2018, 5, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Gouin, J.P.; Connors, J.; Kiecolt-Glaser, J.K.; Glaser, R.; Malarkey, W.B.; Atkinson, C.; Beversdorf, D.; Quan, N. Altered Expression of Circadian Rhythm Genes among Individuals with a History of Depression. J. Affect. Disord. 2010, 126, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Partonen, T.; Treutlein, J.; Alpman, A.; Frank, J.; Johansson, C.; Depner, M.; Aron, L.; Rietschel, M.; Wellek, S.; Soronen, P.; et al. Three Circadian Clock Genes Per2, Arntl, and Npas2 Contribute to Winter Depression. Ann. Med. 2007, 39, 229–238. [Google Scholar] [CrossRef]
- Soria, V.; Martinez-Amoros, E.; Escaramis, G.; Valero, J.; Perez-Egea, R.; Garcia, C.; Gutierrez-Zotes, A.; Puigdemont, D.; Bayes, M.; Crespo, J.M.; et al. Differential Association of Circadian Genes with Mood Disorders: CRY1 and NPAS2 are Associated with Unipolar Major Depression and CLOCK and VIP with Bipolar Disorder. Neuropsychopharmacology 2010, 35, 1279–1289. [Google Scholar] [CrossRef]
- Utge, S.J.; Soronen, P.; Loukola, A.; Kronholm, E.; Ollila, H.M.; Pirkola, S.; Porkka-Heiskanen, T.; Partonen, T.; Paunio, T. Systematic Analysis of Circadian Genes in a Population-Based Sample Reveals Association of TIMELESS with Depression and Sleep Disturbance. PLoS ONE 2010, 5, e9259. [Google Scholar] [CrossRef] [Green Version]
- Mansour, H.A.; Talkowski, M.E.; Wood, J.; Chowdari, K.V.; McClain, L.; Prasad, K.; Montrose, D.; Fagiolini, A.; Friedman, E.S.; Allen, M.H.; et al. Association Study of 21 Circadian Genes with Bipolar I Disorder, Schizoaffective Disorder, and Schizophrenia. Bipolar Disord. 2009, 11, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Nievergelt, C.M.; Kripke, D.F.; Barrett, T.B.; Burg, E.; Remick, R.A.; Sadovnick, A.D.; McElroy, S.L.; Keck, P.E., Jr.; Schork, N.J.; Kelsoe, J.R. Suggestive Evidence for Association of the Circadian Genes PERIOD3 and ARNTL with Bipolar Disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Kripke, D.F.; Nievergelt, C.M.; Joo, E.; Shekhtman, T.; Kelsoe, J.R. Circadian Polymorphisms Associated with Affective Disorders. J. Circadian Rhythm. 2009, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Le-Niculescu, H.; Patel, S.D.; Bhat, M.; Kuczenski, R.; Faraone, S.V.; Tsuang, M.T.; McMahon, F.J.; Schork, N.J.; Nurnberger, J.I., Jr.; Niculescu, A.B., 3rd. Convergent Functional Genomics of Genome-Wide Association Data for Bipolar Disorder: Comprehensive Identification of Candidate Genes, Pathways and Mechanisms. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2009, 150, 155–181. [Google Scholar] [CrossRef] [PubMed]
- Dmitrzak-Weglarz, M.P.; Pawlak, J.M.; Maciukiewicz, M.; Moczko, J.; Wilkosc, M.; Leszczynska-Rodziewicz, A.; Zaremba, D.; Hauser, J. Clock Gene Variants Differentiate Mood Disorders. Mol. Biol. Rep. 2015, 42, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Rybakowski, J.K.; Dmitrzak-Weglarz, M.; Dembinska-Krajewska, D.; Hauser, J.; Akiskal, K.K.; Akiskal, H.H. Polymorphism of Circadian Clock Genes and Temperamental Dimensions of the TEMPS-A in Bipolar Disorder. J. Affect. Disord. 2014, 159, 80–84. [Google Scholar] [CrossRef]
- Benedetti, F.; Dallaspezia, S.; Colombo, C.; Pirovano, A.; Marino, E.; Smeraldi, E. A Length Polymorphism in the Circadian Clock Gene Per3 Influences Age at Onset of Bipolar Disorder. Neurosci. Lett. 2008, 445, 184–187. [Google Scholar] [CrossRef]
- Dallaspezia, S.; Lorenzi, C.; Pirovano, A.; Colombo, C.; Smeraldi, E.; Benedetti, F. Circadian Clock Gene Per3 Variants Influence the Postpartum Onset of Bipolar Disorder. Eur. Psychiatry 2011, 26, 138–140. [Google Scholar] [CrossRef]
- Xu, X.; Breen, G.; Chen, C.K.; Huang, Y.S.; Wu, Y.Y.; Asherson, P. Association Study between a Polymorphism at the 3’-Untranslated Region of CLOCK Gene and Attention Deficit Hyperactivity Disorder. Behav. Brain Funct. 2010, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Carpena, M.X.; Hutz, M.H.; Salatino-Oliveira, A.; Polanczyk, G.V.; Zeni, C.; Schmitz, M.; Chazan, R.; Genro, J.P.; Rohde, L.A.; Tovo-Rodrigues, L. CLOCK Polymorphisms in Attention-Deficit/Hyperactivity Disorder (ADHD): Further Evidence Linking Sleep and Circadian Disturbances and ADHD. Genes (Basel) 2019, 10, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.H.; Yu, J.C.; Lee, C.H.; Choi, K.S.; Choi, J.E.; Kim, S.H.; Joo, E.J. Human CLOCK gene-associated attention deficit hyperactivity disorder-related features in healthy adults: Quantitative association study using Wender Utah Rating Scale. Eur. Arch. Psychiatry Clin. Neurosci. 2014, 264, 71–81. [Google Scholar] [CrossRef]
- Seney, M.L.; Cahill, K.; Enwright, J.F., 3rd; Logan, R.W.; Huo, Z.; Zong, W.; Tseng, G.; McClung, C.A. Diurnal Rhythms in Gene Expression in the Prefrontal Cortex in Schizophrenia. Nat. Commun. 2019, 10, 3355. [Google Scholar] [CrossRef] [Green Version]
- Crnko, S.; Du Pre, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian Rhythms and the Molecular Clock in Cardiovascular Biology and Disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components CLOCK and BMAL1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengesser, S.R.E. (Ed.) Genetics of Bipolar Disorder; Peter-Lang: Frankfurt, Germany, 2013; p. 226. [Google Scholar]
- Hampp, G.; Ripperger, J.A.; Houben, T.; Schmutz, I.; Blex, C.; Perreau-Lenz, S.; Brunk, I.; Spanagel, R.; Ahnert-Hilger, G.; Meijer, J.H.; et al. Regulation of Monoamine Oxidase A by Circadian-Clock Components Implies Clock Influence on Mood. Curr. Biol. 2008, 18, 678–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampp, G.; Albrecht, U. The Circadian Clock and Mood-Related Behavior. Commun. Integr. Biol. 2008, 1, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic Nucleus in the Mouse: Retinal Innervation, Intrinsic Organization and Efferent Projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef]
- Bartlett, D.J.; Biggs, S.N.; Armstrong, S.M. Circadian Rhythm Disorders among Adolescents: Assessment and Treatment Options. Med. J. Aust. 2013, 199, S16–S20. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on Melatonin Receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef]
- Bengesser, S.A.; Reininghaus, E.Z.; Lackner, N.; Birner, A.; Fellendorf, F.T.; Platzer, M.; Kainzbauer, N.; Tropper, B.; Hormanseder, C.; Queissner, R.; et al. Is the Molecular Clock Ticking Differently in Bipolar Disorder? Methylation Analysis of the Clock Gene ARNTL. World J. Biol. Psychiatry 2016, 19, S21–S29. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, Y.; Breitling, L.P.; Brenner, H. Relationship of Tobacco Smoking and Smoking-Related DNA Methylation with Epigenetic Age Acceleration. Oncotarget 2016, 7, 46878–46889. [Google Scholar] [CrossRef] [Green Version]
- Yara, S.; Lavoie, J.C.; Levy, E. Oxidative Stress and DNA Methylation Regulation in the Metabolic Syndrome. Epigenomics 2015, 7, 283–300. [Google Scholar] [CrossRef]
- Qin, Y.; Wade, P.A. Crosstalk between the Microbiome and Epigenome: Messages from Bugs. J. Biochem. 2018, 163, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, M. Multifunctions of Dietary Polyphenols in the Regulation of Intestinal Inflammation. J. Food Drug Anal. 2017, 25, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengesser, S.A.; Morkl, S.; Painold, A.; Dalkner, N.; Birner, A.; Fellendorf, F.T.; Platzer, M.; Queissner, R.; Hamm, C.; Maget, A.; et al. Epigenetics of the Molecular Clock and Bacterial Diversity in Bipolar Disorder. Psychoneuroendocrinology 2019, 101, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hao, Y.; Fan, F.; Zhang, B. The Role of Microbiome in Insomnia, Circadian Disturbance and Depression. Front. Psychiatry 2018, 9, 669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poroyko, V.A.; Carreras, A.; Khalyfa, A.; Khalyfa, A.A.; Leone, V.; Peris, E.; Almendros, I.; Gileles-Hillel, A.; Qiao, Z.; Hubert, N.; et al. Chronic Sleep Disruption Alters Gut Microbiota, Induces Systemic and Adipose Tissue Inflammation and Insulin Resistance in Mice. Sci. Rep. 2016, 6, 35405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnucan, J.A.; Rubin, D.T.; Ali, T. Sleep and Inflammatory Bowel Disease: Exploring the Relationship between Sleep Disturbances and Inflammation. Gastroenterol. Hepatol. (N.Y.) 2013, 9, 718–727. [Google Scholar]
- Wu, G.; Tang, W.; He, Y.; Hu, J.; Gong, S.; He, Z.; Wei, G.; Lv, L.; Jiang, Y.; Zhou, H.; et al. Light exposure influences the diurnal oscillation of gut microbiota in mice. Biochem. Biophys Res. Commun. 2018, 501, 16–23. [Google Scholar] [CrossRef]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Teichman, E.M.; O’Riordan, K.J.; Gahan, C.G.M.; Dinan, T.G.; Cryan, J.F. When Rhythms Meet the Blues: Circadian Interactions with the Microbiota-Gut-Brain Axis. Cell Metab. 2020, 31, 448–471. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom Control of Microbiota Diurnal Oscillations Promotes Metabolic Homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [Green Version]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Engen, P.A.; Keshavarzian, A. Circadian Rhythm and the Gut Microbiome. Int. Rev. Neurobiol. 2016, 131, 193–205. [Google Scholar]
- Ni, Y.; Wu, L.; Jiang, J.; Yang, T.; Wang, Z.; Ma, L.; Zheng, L.; Yang, X.; Wu, Z.; Fu, Z.; et al. Late-Night Eating-Induced Physiological Dysregulation and Circadian Misalignment Are Accompanied by Microbial Dysbiosis. Mol. Nutr. Food Res. 2019, 63, e1900867. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A Novel Class of Psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, S.; Awwad, H.M.; Eskelund, A.R.; Treccani, G.; Geisel, J.; Wegener, G.; Obeid, R. Probiotics Affect One-Carbon Metabolites and Catecholamines in a Genetic Rat Model of Depression. Mol. Nutr. Food Res. 2018, 62, e1701070. [Google Scholar] [CrossRef] [PubMed]
- Burton, K.J.; Pimentel, G.; Zangger, N.; Vionnet, N.; Drai, J.; McTernan, P.G.; Pralong, F.P.; Delorenzi, M.; Vergeres, G. Modulation of the Peripheral Blood Transcriptome by the Ingestion of Probiotic Yoghurt and Acidified Milk in Healthy, Young Men. PLoS ONE 2018, 13, e0192947. [Google Scholar] [CrossRef]
- Solano-Aguilar, G.; Molokin, A.; Botelho, C.; Fiorino, A.M.; Vinyard, B.; Li, R.; Chen, C.; Urban, J., Jr.; Dawson, H.; Andreyeva, I.; et al. Transcriptomic Profile of Whole Blood Cells from Elderly Subjects Fed Probiotic Bacteria Lactobacillus Rhamnosus GG ATCC 53103 (LGG) in a Phase I Open Label Study. PLoS ONE 2016, 11, e0147426. [Google Scholar] [CrossRef]
- De Andres, J.; Jimenez, E.; Espinosa-Martos, I.; Rodriguez, J.M.; Garcia-Conesa, M.T. An Exploratory Search for Potential Molecular Targets Responsive to the Probiotic Lactobacillus Salivarius PS2 in Women with Mastitis: Gene Expression Profiling Vs. Interindividual Variability. Front. Microbiol. 2018, 9, 2166. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, R.; Gibson, G.R.; Vulevic, J.; Giallourou, N.; Castro-Mejia, J.L.; Hansen, L.H.; Leigh Gibson, E.; Nielsen, D.S.; Costabile, A. A Prebiotic Intervention Study in Children with Autism Spectrum Disorders (ASDs). Microbiome 2018, 6, 133. [Google Scholar] [CrossRef]
- Hong, Y.S.; Hong, K.S.; Park, M.H.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Lee, J.; Kim, I.K.; Hwang, G.S.; Kim, J.S. Metabonomic Understanding of Probiotic Effects in Humans with Irritable Bowel Syndrome. J. Clin. Gastroenterol. 2011, 45, 415–425. [Google Scholar] [CrossRef]
- Miccheli, A.; Capuani, G.; Marini, F.; Tomassini, A.; Pratico, G.; Ceccarelli, S.; Gnani, D.; Baviera, G.; Alisi, A.; Putignani, L.; et al. Urinary (1)H-NMR-Based Metabolic Profiling of Children with NAFLD Undergoing VSL#3 Treatment. Int. J. Obes. (Lond.) 2015, 39, 1118–1125. [Google Scholar]
- Kunze, K.N.; Hanlon, E.C.; Prachand, V.N.; Brady, M.J. Peripheral Circadian Misalignment: Contributor to Systemic Insulin Resistance and Potential Intervention to Improve Bariatric Surgical Outcomes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R558–R563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knippers, R. Molekulare Genetik; Thieme: Stuttgart, Germany, 2001; Volume 8, pp. 1–586. [Google Scholar]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalova, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, K.; Plomp, R.; Lao, O.; Middleton, B.; Revell, V.L.; Skene, D.J.; Kayser, M. Effect of Sleep Deprivation on Rhythms of Clock Gene Expression and Melatonin in Humans. Chronobiol. Int. 2013, 30, 901–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the Intestinal Microbiota is Regulated by Gender and the Host Circadian Clock. Proc. Natl. Acad. Sci. USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, S.J.; Vargas, F.; Gonzalez, A.; He, S.; Jiang, P.; Dorrestein, P.C.; Knight, R.; Wright, K.P., Jr.; Lowry, C.A.; Fleshner, M.; et al. Repeated Sleep Disruption in Mice Leads to Persistent Shifts in the Fecal Microbiome and Metabolome. PLoS ONE 2020, 15, e0229001. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Wang, Z.; Dong, Y.; Cao, J.; Lin, R.; Wang, X.; Yu, Z.; Chen, Y. Role of Melatonin in Sleep Deprivation-Induced Intestinal Barrier Dysfunction in Mice. J. Pineal Res. 2019, 67, e12574. [Google Scholar] [CrossRef]
- Godinho-Silva, C.; Domingues, R.G.; Rendas, M.; Raposo, B.; Ribeiro, H.; da Silva, J.A.; Vieira, A.; Costa, R.M.; Barbosa-Morais, N.L.; Carvalho, T.; et al. Light-Entrained and Brain-Tuned Circadian Circuits Regulate ILC3s and Gut Homeostasis. Nature 2019, 574, 254–258. [Google Scholar] [CrossRef]
- Ridaura, V.; Belkaid, Y. Gut Microbiota: The Link to Your Second Brain. Cell 2015, 161, 193–194. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.L.; Teesson, M.; Mills, K.L. Associations between Substance use, Post-Traumatic Stress Disorder and the Perpetration of Violence: A Longitudinal Investigation. Addict. Behav. 2014, 39, 1075–1080. [Google Scholar] [CrossRef]
- Breit, S.; Kupferberg, A.; Rogler, G.; Hasler, G. Vagus Nerve as Modulator of the Brain-Gut Axis in Psychiatric and Inflammatory Disorders. Front. Psychiatry. 2018, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Chen, W.D.; Wang, Y.D. Gut Microbiota: An Integral Moderator in Health and Disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kasper, L.H. The Role of Microbiome in Central Nervous System Disorders. Brain Behav. Immun. 2014, 38, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, F.L. Simplified Instrumentation for the Clinical Laboratory. Bull. Pan Am. Health Organ. 1976, 10, 212–218. [Google Scholar]
- Evans, S.J.; Bassis, C.M.; Hein, R.; Assari, S.; Flowers, S.A.; Kelly, M.B.; Young, V.B.; Ellingrod, V.E.; McInnis, M.G. The Gut Microbiome Composition Associates with Bipolar Disorder and Illness Severity. J. Psychiatr. Res. 2017, 87, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizawa, E.; Tsuji, H.; Asahara, T.; Takahashi, T.; Teraishi, T.; Yoshida, S.; Koga, N.; Hattori, K.; Ota, M.; Kunugi, H. Bifidobacterium and Lactobacillus Counts in the Gut Microbiota of Patients with Bipolar Disorder and Healthy Controls. Front. Psychiatry 2019, 9, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, S.; Kishimoto, T.; Mizuno, S.; Masaoka, T.; Naganuma, M.; Liang, K.C.; Kitazawa, M.; Nakashima, M.; Shindo, C.; Suda, W.; et al. The Effect of Fecal Microbiota Transplantation on Psychiatric Symptoms among Patients with Irritable Bowel Syndrome, Functional Diarrhea and Functional Constipation: An Open-Label Observational Study. J. Affect. Disord. 2018, 235, 506–512. [Google Scholar] [CrossRef]
- Hold, G.L.; Hansen, R. Impact of the Gastrointestinal Microbiome in Health and Disease: Co-Evolution with the Host Immune System. Curr. Top. Microbiol. Immunol. 2019, 421, 303–318. [Google Scholar]
- Szentirmai, E.; Kapas, L. Sleep and Body Temperature in TNFalpha Knockout Mice: The Effects of Sleep Deprivation, beta3-AR Stimulation and Exogenous TNFalpha. Brain Behav. Immun. 2019, 81, 260–271. [Google Scholar] [CrossRef]
- Takada, M.; Nishida, K.; Gondo, Y.; Kikuchi-Hayakawa, H.; Ishikawa, H.; Suda, K.; Kawai, M.; Hoshi, R.; Kuwano, Y.; Miyazaki, K.; et al. Beneficial Effects of Lactobacillus Casei Strain Shirota on Academic Stress-Induced Sleep Disturbance in Healthy Adults: A Double-Blind, Randomised, Placebo-Controlled Trial. Benef. Microbes 2017, 8, 153–162. [Google Scholar] [CrossRef]
- Miyazaki, K.; Itoh, N.; Yamamoto, S.; Higo-Yamamoto, S.; Nakakita, Y.; Kaneda, H.; Shigyo, T.; Oishi, K. Dietary Heat-Killed Lactobacillus Brevis SBC8803 Promotes Voluntary Wheel-Running and Affects Sleep Rhythms in Mice. Life Sci. 2014, 111, 47–52. [Google Scholar] [CrossRef]
- Sandhu, K.V.; Sherwin, E.; Schellekens, H.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Feeding the Microbiota-Gut-Brain Axis: Diet, Microbiome, and Neuropsychiatry. Transl. Res. 2017, 179, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.E.; Hsu, T.M.; Kanoski, S.E. Gut to Brain Dysbiosis: Mechanisms Linking Western Diet Consumption, the Microbiome, and Cognitive Impairment. Front. Behav. Neurosci. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The Neuroactive Potential of the Human Gut Microbiota in Quality of Life and Depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid Metabolism: The Interaction of Metabolites and Gut Microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beli, E.; Prabakaran, S.; Krishnan, P.; Evans-Molina, C.; Grant, M.B. Loss of Diurnal Oscillatory Rhythms in Gut Microbiota Correlates with Changes in Circulating Metabolites in Type 2 Diabetic db/db Mice. Nutrients 2019, 11, 2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Mutlu, E.; Engen, P.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. Circadian Disorganization Alters Intestinal Microbiota. PLoS ONE 2014, 9, e97500. [Google Scholar] [CrossRef]
- Tahara, Y.; Yamazaki, M.; Sukigara, H.; Motohashi, H.; Sasaki, H.; Miyakawa, H.; Haraguchi, A.; Ikeda, Y.; Fukuda, S.; Shibata, S. Gut Microbiota-Derived Short Chain Fatty Acids Induce Circadian Clock Entrainment in Mouse Peripheral Tissue. Sci. Rep. 2018, 8, 1395. [Google Scholar] [CrossRef]
- Szentirmai, E.; Millican, N.S.; Massie, A.R.; Kapas, L. Butyrate, a Metabolite of Intestinal Bacteria, Enhances Sleep. Sci. Rep. 2019, 9, 7035. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Lin, W.; Chen, S.; Xiang, T.; Yang, Y.; Yin, Y.; Xu, G.; Liu, Z.; Liu, L.; Pan, J.; et al. Gut Microbiota as a Subjective Measurement for Auxiliary Diagnosis of Insomnia Disorder. Front. Microbiol. 2019, 10, 1770. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.P.; Easson, C.; Lyle, S.M.; Kapoor, R.; Donnelly, C.P.; Davidson, E.J.; Parikh, E.; Lopez, J.V.; Tartar, J.L. Gut Microbiome Diversity is Associated with Sleep Physiology in Humans. PLoS ONE 2019, 14, e0222394. [Google Scholar] [CrossRef]
- Jansen, E.C.; She, R.; Rukstalis, M.M.; Alexander, G.L. Sleep Duration and Quality in Relation to Fruit and Vegetable Intake of US Young Adults: A Secondary Analysis. Int. J. Behav. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and Metabolic Response to Probiotic Administration in Patients with Major Depressive Disorder: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Gruenwald, J.; Graubaum, H.J.; Harde, A. Effect of a Probiotic Multivitamin Compound on Stress and Exhaustion. Adv. Ther. 2002, 19, 141–150. [Google Scholar] [CrossRef]
- Yu, L.; Han, X.; Cen, S.; Duan, H.; Feng, S.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; et al. Beneficial Effect of GABA-Rich Fermented Milk on Insomnia Involving Regulation of Gut Microbiota. Microbiol. Res. 2020, 233, 126409. [Google Scholar] [CrossRef] [PubMed]
- Wallace, C.J.K.; Milev, R. The Effects of Probiotics on Depressive Symptoms in Humans: A Systematic Review. Ann. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.S.; Vargas, F.; Dorrestein, P.C.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Dietary Prebiotics Alter Novel Microbial Dependent Fecal Metabolites that Improve Sleep. Sci. Rep. 2020, 10, 3848. [Google Scholar] [CrossRef] [Green Version]
- Pot, G.K. Sleep and Dietary Habits in the Urban Environment: The Role of Chrono-Nutrition. Proc. Nutr. Soc. 2018, 77, 189–198. [Google Scholar] [CrossRef] [Green Version]
- St-Onge, M.P.; Mikic, A.; Pietrolungo, C.E. Effects of Diet on Sleep Quality. Adv. Nutr. 2016, 7, 938–949. [Google Scholar] [CrossRef]
- Taniguchi, H.; Fernandez, A.F.; Setien, F.; Ropero, S.; Ballestar, E.; Villanueva, A.; Yamamoto, H.; Imai, K.; Shinomura, Y.; Esteller, M. Epigenetic Inactivation of the Circadian Clock Gene BMAL1 in Hematologic Malignancies. Cancer Res. 2009, 69, 8447–8454. [Google Scholar] [CrossRef] [Green Version]
- Iitaka, C.; Miyazaki, K.; Akaike, T.; Ishida, N. A Role for Glycogen Synthase Kinase-3beta in the Mammalian Circadian Clock. J. Biol. Chem. 2005, 280, 29397–29402. [Google Scholar] [CrossRef] [Green Version]
- Iwahana, E.; Akiyama, M.; Miyakawa, K.; Uchida, A.; Kasahara, J.; Fukunaga, K.; Hamada, T.; Shibata, S. Effect of Lithium on the Circadian Rhythms of Locomotor Activity and Glycogen Synthase Kinase-3 Protein Expression in the Mouse Suprachiasmatic Nuclei. Eur. J. Neurosci. 2004, 19, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.; Schlarb, A.A. Let’s Talk about Sleep: A Systematic Review of Psychological Interventions to Improve Sleep in College Students. J. Sleep Res. 2018, 27, 4–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, P.L.; Gengler, D.; Kelly, M. Social Rhythm Therapies for Mood Disorders: An Update. Curr. Psychiatry Rep. 2016, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Crowe, M.; Inder, M.; Swartz, H.A.; Murray, G.; Porter, R. Social Rhythm Therapy-A Potentially Translatable Psychosocial Intervention for Bipolar Disorder. Bipolar Disord. 2020, 22, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, A.K.; Kinrys, G. Treating Circadian Rhythm Disruption in Bipolar Disorder. Curr. Psychiatry Rep. 2019, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Crowe, M.; Inder, M.; Douglas, K.; Carlyle, D.; Wells, H.; Jordan, J.; Lacey, C.; Mulder, R.; Beaglehole, B.; Porter, R. Interpersonal and Social Rhythm Therapy for Patients with Major Depressive Disorder. Am. J. Psychother. 2020, 73, 29–34. [Google Scholar] [CrossRef]
- Hickie, I.B.; Naismith, S.L.; Robillard, R.; Scott, E.M.; Hermens, D.F. Manipulating the Sleep-Wake Cycle and Circadian Rhythms to Improve Clinical Management of Major Depression. BMC Med. 2013, 11, 79. [Google Scholar] [CrossRef]
- Melo, M.C.A.; Abreu, R.L.C.; Linhares Neto, V.B.; de Bruin, P.F.C.; de Bruin, V.M.S. Chronotype and Circadian Rhythm in Bipolar Disorder: A Systematic Review. Sleep Med. Rev. 2017, 34, 46–58. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Case Control Studies Investigating Sleep and Microbiome in Psychiatric Disorders | ||||
| Author and Year of Publication | Population | Measurements/Interventions | Comparison | Outcome |
| Evans et al., 2016 | Individuals with bipolar disorder | Pittsburg Sleep Quality Index (PSQI), Operational Taxonomical Unit level analysis (OTU), molecular variance (AMOVA), Short Form Health Survey (SF12), Generalized Anxiety Disorder scale (GAD), Altman Self-Rating Mania Scale (ASRM) | Bipolar Disorder (n = 115) healthy controls (n = 64); cross-sectional study | Negative relationship between Faecalibacterium and sleep quality (PSQI) at the subscale level (ß = −0.329; p = 0.001; d = −1.357; r = −0.5) |
| Aizawa et al., 2019 | Individuals with Bipolar Disorder | Bacterial counts (RT-qPCR) Hamilton Depression Rating Scale (HAM-D), Young Mania Rating Scale | Bipolar Disorder (13 bipolar Type I, 26 bipolar Type II), 58 healthy controls; cross-sectional study | Negative correlation between Lactobacillus counts and sleep. (p = −0.45, P = 0.01; d = 0.885; r = 0.405) |
| Intervention Studies Investigating Sleep and Microbiome in Psychiatric Disorders | ||||
| Author and Year of Publication | Population | Interventions | Comparison | Outcome |
| Kurokawa et al., 2018 | Individuals with Irritable Bowel Syndrome (IBS) | Fecal Microbiota Transplantation, Hamilton Rating Scale (HAM-D), Subscale of sleep related items, Hamilton Rating Scale for Anxiety (HAM-A), Quick Inventory for Depressive | 17 Patients with Irritable Bowel Syndrome (IBS), functional diarrhea FDr, functional constipation FC, DSM IV of Comorbid depression and anxiety, Intervention Study, single center, open-label, non-randomized observational study | Significant improvement in HAM-D total and sleep- subscale score. Baseline Shannon index show lower diversity in patients with HAM-D > 8 compared with patients HAMD < 8 (p = 0.005, FDR = 0.007) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner-Skacel, J.; Dalkner, N.; Moerkl, S.; Kreuzer, K.; Farzi, A.; Lackner, S.; Painold, A.; Reininghaus, E.Z.; Butler, M.I.; Bengesser, S. Sleep and Microbiome in Psychiatric Diseases. Nutrients 2020, 12, 2198. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082198
Wagner-Skacel J, Dalkner N, Moerkl S, Kreuzer K, Farzi A, Lackner S, Painold A, Reininghaus EZ, Butler MI, Bengesser S. Sleep and Microbiome in Psychiatric Diseases. Nutrients. 2020; 12(8):2198. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082198
Chicago/Turabian StyleWagner-Skacel, Jolana, Nina Dalkner, Sabrina Moerkl, Kathrin Kreuzer, Aitak Farzi, Sonja Lackner, Annamaria Painold, Eva Z. Reininghaus, Mary I. Butler, and Susanne Bengesser. 2020. "Sleep and Microbiome in Psychiatric Diseases" Nutrients 12, no. 8: 2198. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082198