Effects of Multi-Strain Probiotics on Immune Responses and Metabolic Balance in Helicobacter pylori-Infected Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Cell Lines, and Culture Conditions
2.2. The Mice Infection Model
2.3. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) Evaluation of the Immune Responses of H. pylori-Infected Tissue and Cells
2.5. Amino Acids and Acylcarnitines Analysis
2.6. Statistical Analysis
3. Results
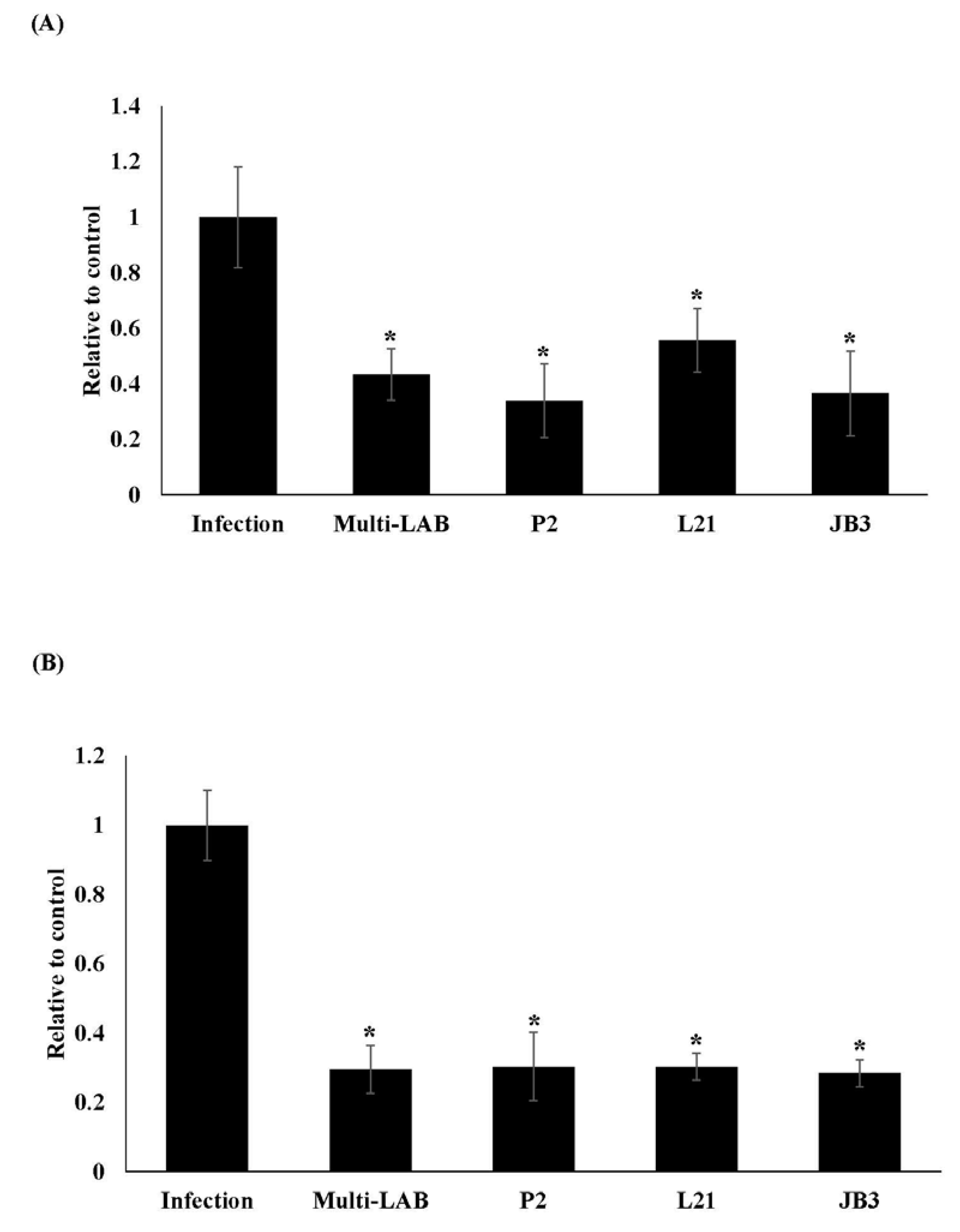
3.1. Effects of LAB Treatments on H. pylori Colonization
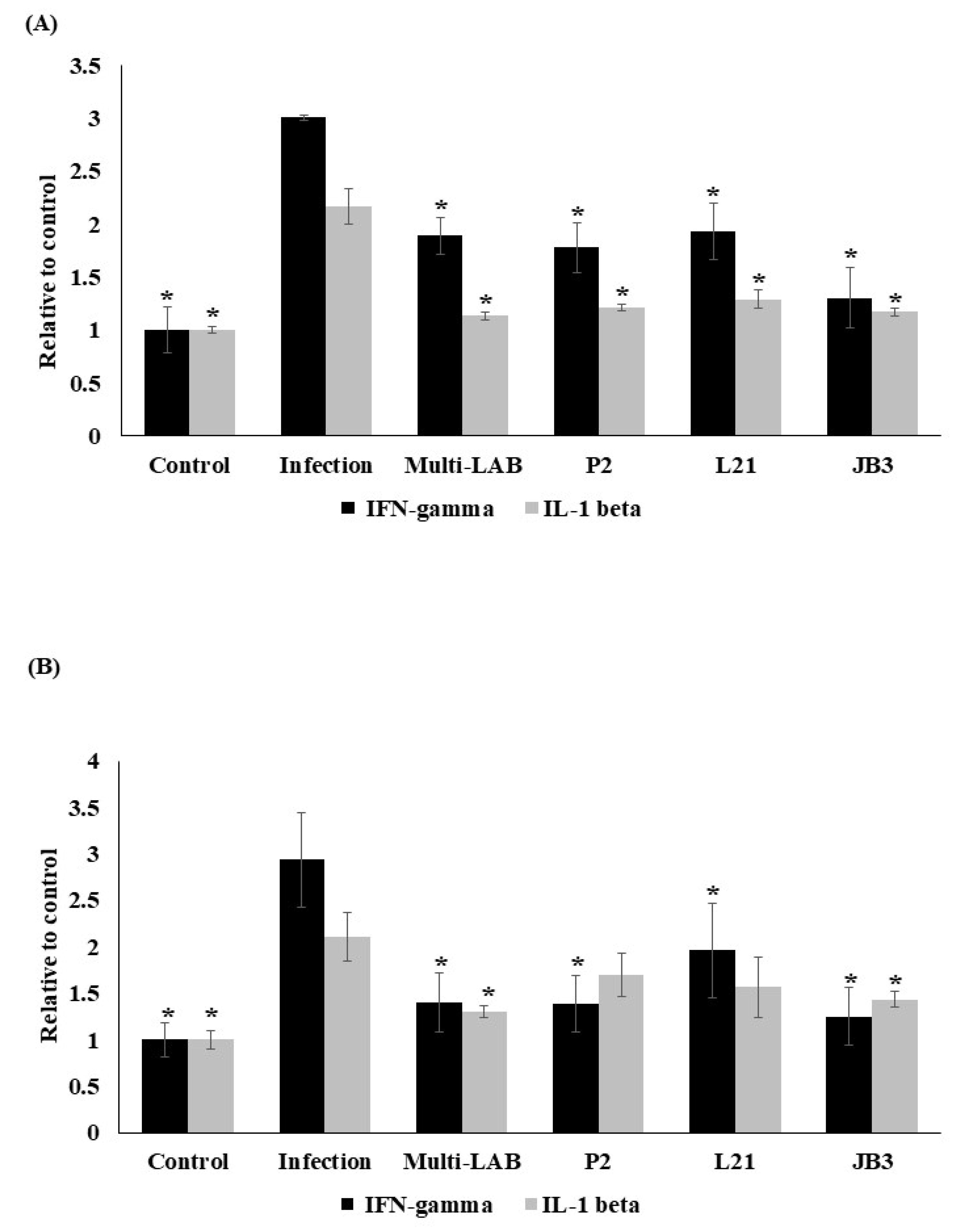
3.2. Effects of LAB Treatments on H. pylori-Induced Immune Response
3.3. Effects of LAB Treatments on H. pylori-Induced Inflammation
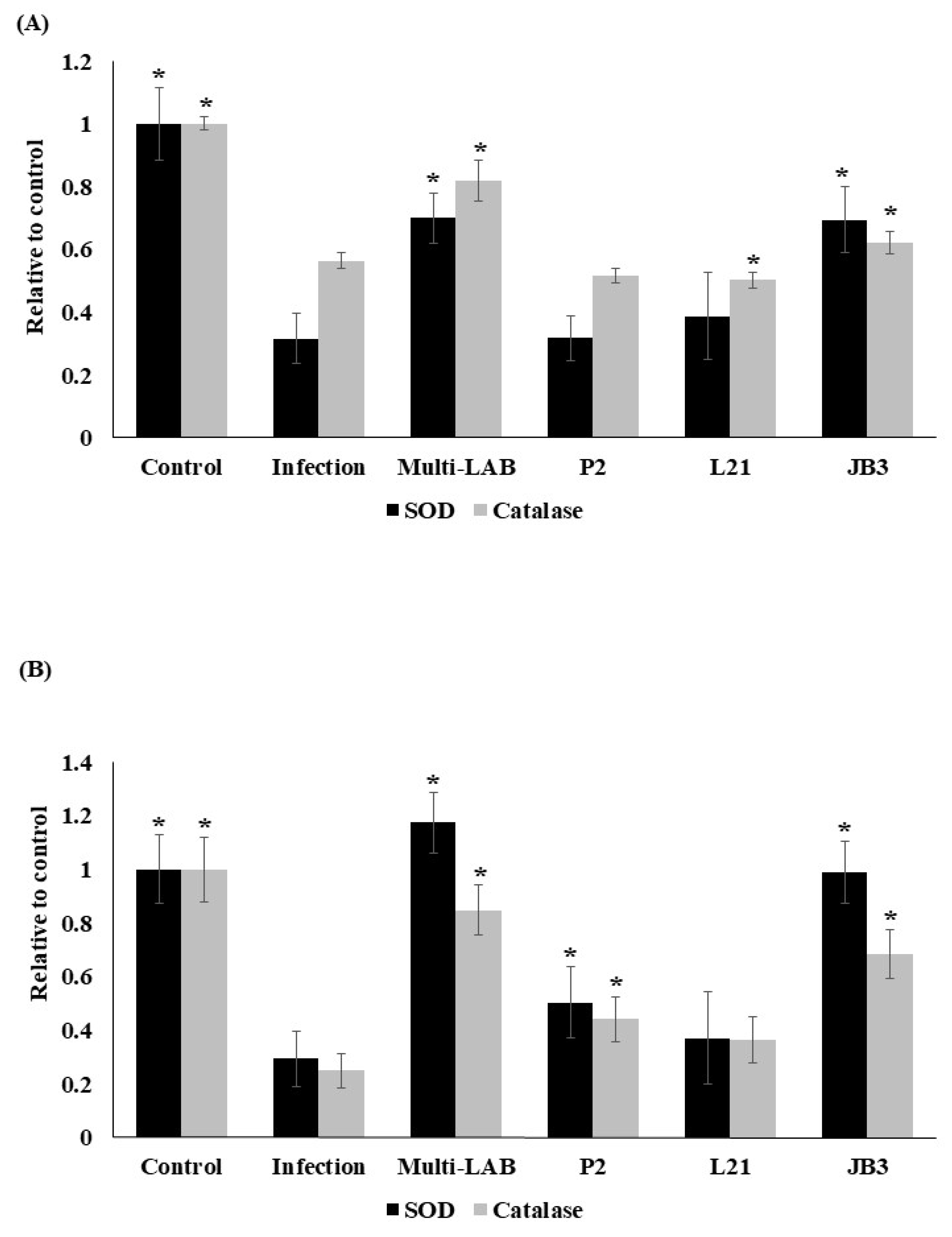
3.4. Effects of LAB Treatments on H. pylori-Induced Oxidative stress
3.5. Effects of LAB Treatments on H. pylori-Infection Mediated Serum Concentrations of Ammo Acids and Fatty Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sjomina, O.; Pavlova, J.; Niv, Y.; Leja, M. Epidemiology of Helicobacter pylori infection. Helicobacter 2018, 23 (Suppl. 1), e12514. [Google Scholar] [CrossRef] [Green Version]
- Leja, M.; Grinberga-Derica, I.; Bilgilier, C.; Steininger, C. Review: Epidemiology of Helicobacter pylori infection. Helicobacter 2019, 24 (Suppl. 1), e12635. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, L.E.; Peek, R.M., Jr.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ables, A.Z.; Simon, I.; Melton, E.R. Update on Helicobacter pylori treatment. Am. Fam. Physician 2007, 75, 351–358. [Google Scholar] [PubMed]
- Alba, C.; Blanco, A.; Alarcon, T. Antibiotic resistance in Helicobacter pylori. Curr. Opin. Infect. Dis. 2017, 30, 489–497. [Google Scholar] [CrossRef]
- Goderska, K.; Agudo Pena, S.; Alarcon, T. Helicobacter pylori treatment: Antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef]
- Michetti, P.; Dorta, G.; Wiesel, P.H.; Brassart, D.; Verdu, E.; Herranz, M.; Felley, C.; Porta, N.; Rouvet, M.; Blum, A.L.; et al. Effect of whey-based culture supernatant of Lactobacillus acidophilus (johnsonii) La1 on Helicobacter pylori infection in humans. Digestion 1999, 60, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.E.; Su, C.H.; Yang, J.S.; Lu, C.C.; Hou, Y.C.; Wu, J.B.; Hsu, Y.M. Baicalin, Baicalein, and Lactobacillus Rhamnosus JB3 Alleviated Helicobacter pylori infections in vitro and in vivo. J. Food Sci. 2018, 83, 3118–3125. [Google Scholar] [CrossRef]
- Pantoflickova, D.; Corthesy-Theulaz, I.; Dorta, G.; Stolte, M.; Isler, P.; Rochat, F.; Enslen, M.; Blum, A.L. Favourable effect of regular intake of fermented milk containing Lactobacillus johnsonii on Helicobacter pylori associated gastritis. Aliment. Pharmacol. Ther. 2003, 18, 805–813. [Google Scholar] [CrossRef]
- Ren, W.; Rajendran, R.; Zhao, Y.; Tan, B.; Wu, G.; Bazer, F.W.; Zhu, G.; Peng, Y.; Huang, X.; Deng, J.; et al. Amino acids as mediators of metabolic cross talk between host and pathogen. Front. Immunol. 2018, 9, 319. [Google Scholar] [CrossRef]
- Lin, W.H.; Yu, B.; Jang, S.H.; Tsen, H.Y. Different probiotic properties for Lactobacillus fermentum strains isolated from swine and poultry. Anaerobe 2007, 13, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. Effects of multi-strain probiotics combined with gardeniae fructus on intestinal microbiota, metabolites, and morphology in broilers. J. Poult. Sci. 2019, 56, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, L.; Kelly, L.K.; Ayub, K.; Graham, D.Y.; Go, M.F. Genotypes of Helicobacter pylori obtained from gastric ulcer patients taking or not taking NSAIDs. Am. J. Gastroenterol. 1999, 94, 1502–1507. [Google Scholar] [CrossRef]
- Lin, W.D.; Wu, J.Y.; Lai, C.C.; Tsai, F.J.; Tsai, C.H.; Lin, S.P.; Niu, D.M. A pilot study of neonatal screening by electrospray ionization tandem mass spectrometry in Taiwan. Acta Paediatr. Taiwan 2001, 42, 224–230. [Google Scholar] [PubMed]
- Rashed, M.S.; Ozand, P.T.; Bucknall, M.P.; Little, D. Diagnosis of inborn errors of metabolism from blood spots by acylcarnitines and amino acids profiling using automated electrospray tandem mass spectrometry. Pediatr. Res. 1995, 38, 324–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Dhiman, M. Inflammasome activation and regulation during Helicobacter pylori pathogenesis. Microb. Pathog. 2018, 125, 468–474. [Google Scholar] [CrossRef]
- Kurpad, A.V. The requirements of protein & amino acid during acute & chronic infections. Indian J. Med. Res. 2006, 124, 129–148. [Google Scholar]
- Bang, C.S.; Baik, G.H. Time to learn from the past and prepare for the future in Helicobacter pylori eradication. Korean J. Intern. Med. 2015, 30, 789–791. [Google Scholar] [CrossRef]
- Feng, J.R.; Wang, F.; Qiu, X.; McFarland, L.V.; Chen, P.F.; Zhou, R.; Liu, J.; Zhao, Q.; Li, J. Efficacy and safety of probiotic-supplemented triple therapy for eradication of Helicobacter pylori in children: A systematic review and network meta-analysis. Eur. J. Clin. Pharmacol. 2017, 73, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Chen, S.; Yin, J.; Duan, J.; Li, T.; Liu, G.; Feng, Z.; Tan, B.; Yin, Y.; Wu, G. Dietary arginine supplementation of mice alters the microbial population and activates intestinal innate immunity. J. Nutr. 2014, 144, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Luke, J.D.; Kullas, A.L.; Kapilashrami, K.; Botbol, Y.; Koller, A.; Tonge, P.J.; Chen, E.I.; Macian, F.; van der Velden, A.W. Asparagine deprivation mediated by Salmonella asparaginase causes suppression of activation-induced T cell metabolic reprogramming. J. Leukoc. Biol. 2016, 99, 387–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Nagata, K.; Nagata, Y.; Sato, T.; Fujino, M.A.; Nakajima, K.; Tamura, T. L-Serine, D- and L-proline and alanine as respiratory substrates of Helicobacter pylori: Correlation between in vitro and in vivo amino acid levels. Microbiology 2003, 149, 2023–2030. [Google Scholar] [CrossRef]
- Jung, S.W.; Lee, S.W. The antibacterial effect of fatty acids on Helicobacter pylori infection. Korean J. Intern. Med. 2016, 31, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Khulusi, S.; Ahmed, H.A.; Patel, P.; Mendall, M.A.; Northfield, T.C. The effects of unsaturated fatty acids on Helicobacter pylori in vitro. J. Med. Microbiol. 1995, 42, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.Q.; O’Connor, C.J.; Roberton, A.M. Antibacterial actions of fatty acids and monoglycerides against Helicobacter pylori. FEMS Immunol. Med. Microbiol. 2003, 36, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated fatty acids and risk of coronary heart disease: Modulation by replacement nutrients. Curr. Atheroscler. Rep. 2010, 12, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Senyilmaz-Tiebe, D.; Pfaff, D.H.; Virtue, S.; Schwarz, K.V.; Fleming, T.; Altamura, S.; Muckenthaler, M.U.; Okun, J.G.; Vidal-Puig, A.; Nawroth, P.; et al. Dietary stearic acid regulates mitochondria in vivo in humans. Nat. Commun. 2018, 9, 3129. [Google Scholar] [CrossRef] [PubMed]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vazquez-Carrera, M. Palmitic and oleic acid: The yin and yang of fatty acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2018, 29, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Muir, J.G.; Gibson, P.R. Does butyrate protect from colorectal cancer? J. Gastroenterol. Hepatol. 2006, 21, 209–218. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Control | Infection | Multi-LAB | P2 | L21 | JB3 |
|---|---|---|---|---|---|---|
| Alanine | 387.27 ± 15.54 b | 170.69 ± 4.00 c | 385.90 ± 12.24 b | 452.97 ± 16.63 a | 358.01 ± 12.43 b | 370.54 ± 18.48 b |
| Arginine | 244.91 ± 20.99 a | 169.04 ± 8.65 c | 256.76 ± 11.04 a | 237.04 ± 10.01 a | 213.72 ± 9.35 a,b | 183.65 ± 15.56 b,c |
| Aspartate | 15.66 ± 2.14 a | 7.01 ± 0.86 c | 14.58 ± 0.23 a,b | 13.05 ± 1.21 a,b | 11.18 ± 0.77 b | 11.70 ± 0.93 b |
| Cysteine | 1.29 ± 0.16 a | 0.20 ± 0.14 b | 0.60 ± 0.26 b | 0.56 ± 0.27 b | 0.50 ± 0.24 b | 0.40 ± 0.12 b |
| Glutamine | 1761.05 ± 68.33 a | 1324.64 ± 65.06 c,d | 1316.64 ± 37.69 c,d | 1462.99 ± 42.32 b,c | 1565.80 ± 31.85 b | 1217.54 ± 67.22 d |
| Glutamate | 165.03 ± 12.18 a | 87.52 ± 3.80 c | 140.44 ± 4.06 b | 125.19 ± 5.97 b | 123.30 ± 4.47 b | 141.59 ± 6.07 b |
| Glycine | 310.79 ± 14.03 a | 260.17 ± 9.95 b | 327.63 ± 12.2 a | 309.92 ± 8.89 a | 324.10 ± 9.68 a | 317.36 ± 9.22 a |
| Leucine/Isoleucine | 268.83 ± 12.11 a | 183.40 ± 5.14 d | 191.54 ± 3.88 b,c | 162.92 ± 5.85 c | 181.73 ± 9.14 c | 207.68 ± 2.88 b |
| Methionine | 62.83 ± 3.87 a | 29.56 ± 1.18 c | 50.54 ± 3.07 b | 50.07 ± 1.37 b | 44.65 ± 1.36 b | 44.64 ± 2.20 b |
| Phenylalanine | 98.09 ± 3.98 a | 78.95 ± 2.88 c | 85.34 ± 2.17 b,c | 80.90 ± 2.28 b,c | 88.14 ± 2.78 b | 96.41 ± 2.43 a |
| Proline | 372.18 ± 18.59 a,b | 224.47 ± 10.47 d | 276.82 ± 10.56 c | 338.46 ± 11.66 b | 382.92 ± 20.11 a | 367.11 ± 11.49 a,b |
| Serine | 63.32 ± 2.44 a | 33.48 ± 1.69 d | 52.06 ± 0.87 b | 47.27 ± 1.55b c | 48.67 ± 1.19b c | 43.91 ± 1.77 c |
| Threonine | 83.03 ± 5.10 a | 42.15 ± 1.49 c | 56.60 ± 1.46 b | 58.89 ± 2.72 b | 59.91 ± 3.46 b | 57.42 ± 2.04 b |
| Tryptophan | 73.49 ± 3.20 a | 51.36 ± 3.64 c | 69.89 ± 4.23 a | 64.21 ± 4.30 a,b | 55.54 ± 2.67 b,c | 53.42 ± 2.63 c |
| Fatty Acid | Control | Infection | Multi-LAB | P2 | L21 | JB3 |
|---|---|---|---|---|---|---|
| Butyric acid (C4) | 0.86 ± 0.12 b,c | 0.81 ± 0.15 b,c | 1.71 ± 0.42 a | 1.49 ± 0.28 a,b | 0.64 ± 0.09 c | 1.06 ± 0.26 a,b,c |
| Valeric acid (C5) | 0.14 ± 0.04 b | 0.07 ± 0.01 b,c | 0.21 ± 0.02 a | 0.06 ± 0.03 c | 0.07 ± 0.02 b,c | 0.04 ± 0.01 c |
| Palmitic acid (C16:0) | 0.70 ± 0.08 a | 0.32 ± 0.04 b | 0.78 ± 0.10 a | 0.49 ± 0.02 b | 0.38 ± 0.02 b | 0.34 ± 0.06 b |
| Palmitoleic acid (C16:1) | 0.18 ± 0.03 a,b | 0.14 ± 0.02 b | 0.25 ± 0.05 a | 0.09 ± 0.02 b | 0.14 ± 0.01 b | 0.15 ± 0.04 b |
| Stearic acid (C18) | 0.11 ± 0.01 a,b | 0.09 ± 0.01 a,b | 0.16 ± 0.04 a | 0.08 ± 0.04 b | 0.06 ± 0.02 b | 0.05 ± 0.02 b |
| Oleic acid (C18:1) | 0.35 ± 0.05 b | 0.28 ± 0.02 b | 0.52 ± 0.03 a | 0.33 ± 0.03 b | 0.31 ± 0.04 b | 0.27 ± 0.04 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-C.; Huang, W.-C.; Su, C.-H.; Lin, W.-D.; Wu, W.-T.; Yu, B.; Hsu, Y.-M. Effects of Multi-Strain Probiotics on Immune Responses and Metabolic Balance in Helicobacter pylori-Infected Mice. Nutrients 2020, 12, 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082476
Lin C-C, Huang W-C, Su C-H, Lin W-D, Wu W-T, Yu B, Hsu Y-M. Effects of Multi-Strain Probiotics on Immune Responses and Metabolic Balance in Helicobacter pylori-Infected Mice. Nutrients. 2020; 12(8):2476. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082476
Chicago/Turabian StyleLin, Chun-Che, Wei-Chiao Huang, Chiu-Hsian Su, Wei-De Lin, Wen-Tzu Wu, Bi Yu, and Yuan-Man Hsu. 2020. "Effects of Multi-Strain Probiotics on Immune Responses and Metabolic Balance in Helicobacter pylori-Infected Mice" Nutrients 12, no. 8: 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082476