Childhood Malnutrition and Association of Lean Mass with Metabolome and Hormone Profile in Later Life

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Setting
2.2. Participants
2.3. Variables
2.4. Data Sources and Measurement
2.5. Study Size
2.6. Data Analysis
2.6.1. Individual Metabolites and Hormones
2.6.2. Path Analysis
2.6.3. Predictive Model for Lean Mass Index Using Metabolome and Hormonal Profile
3. Results
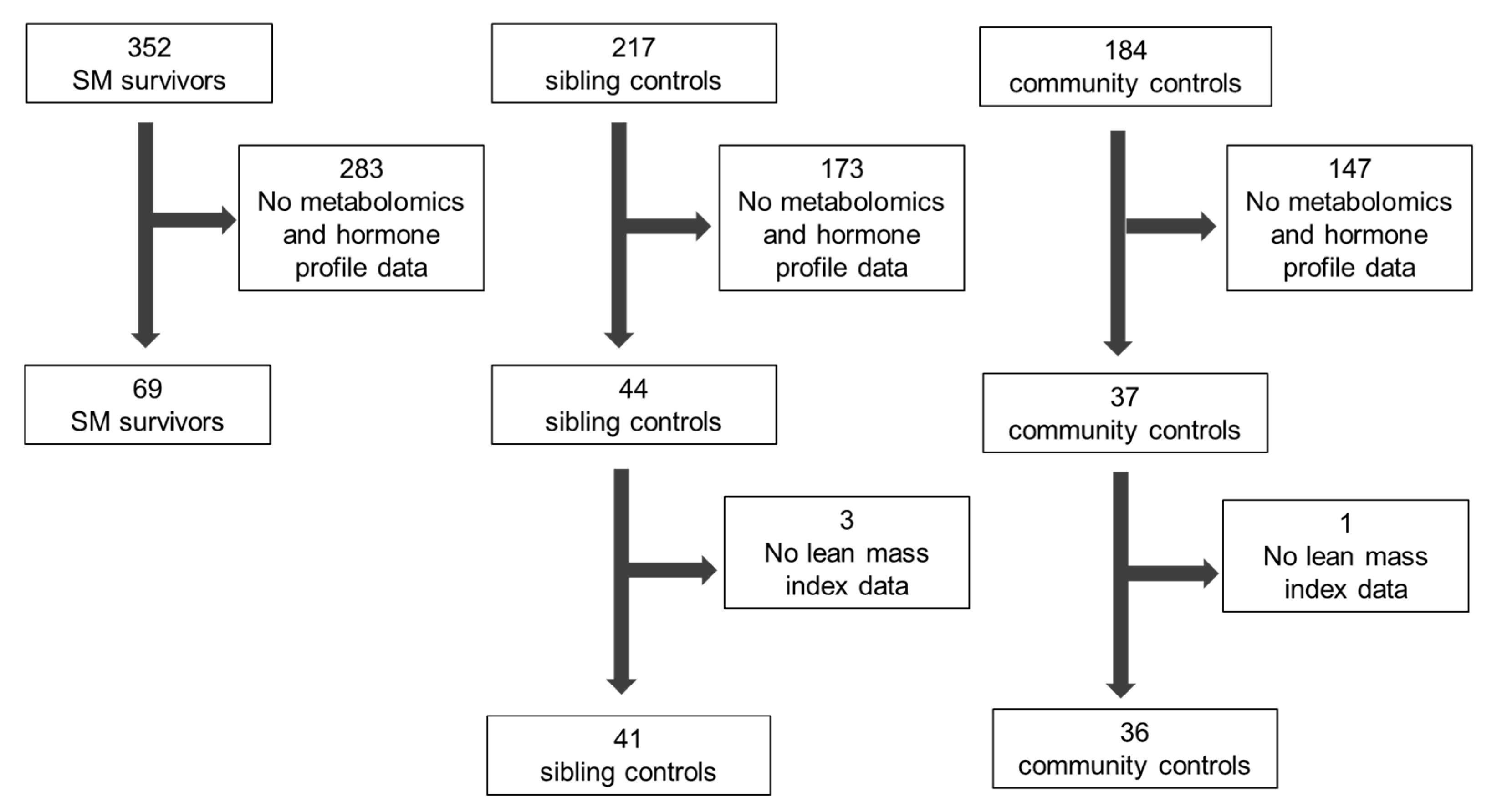
3.1. Study Participants
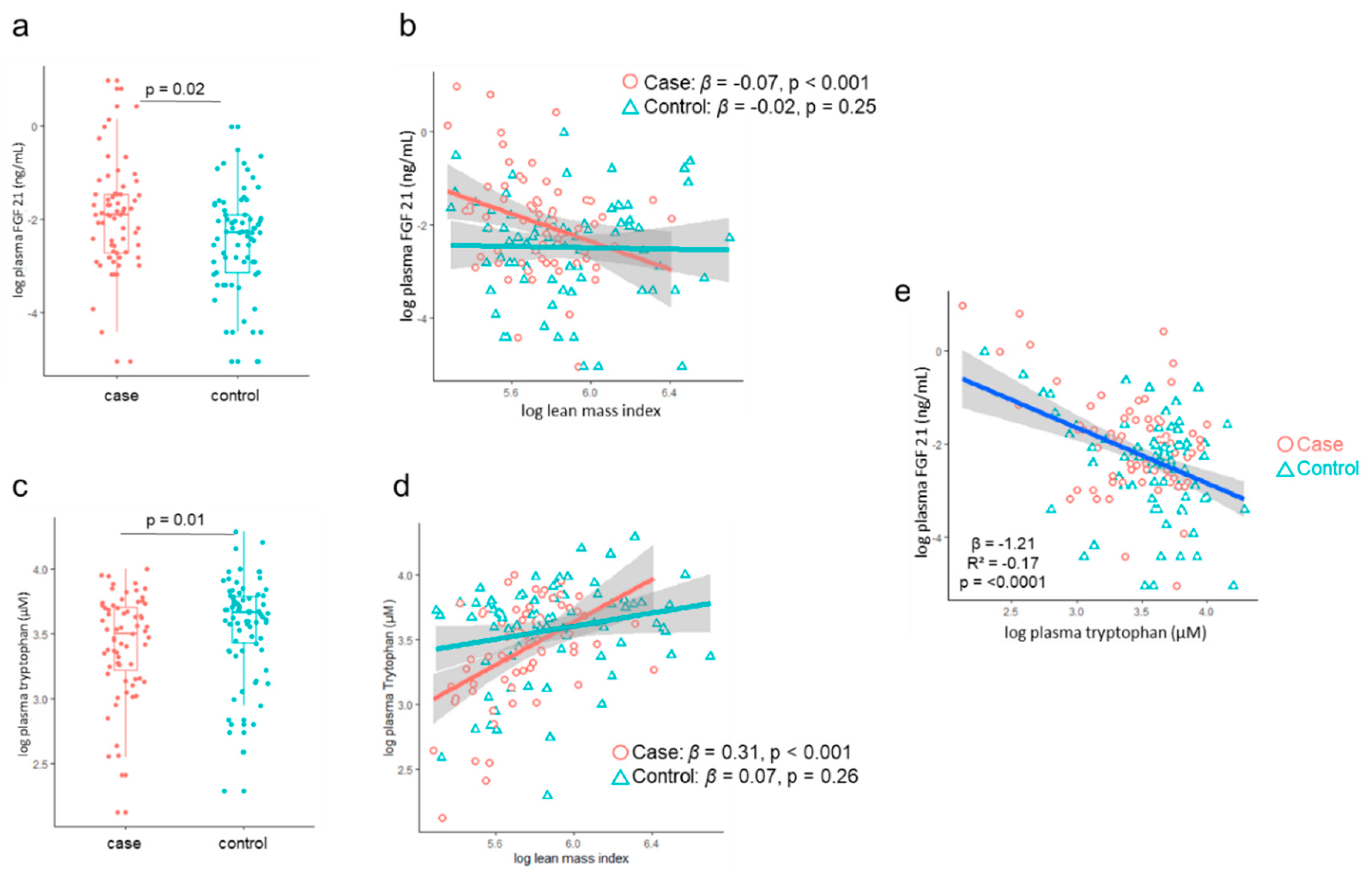
3.2. LMI Association with Metabolome and Hormone Profile
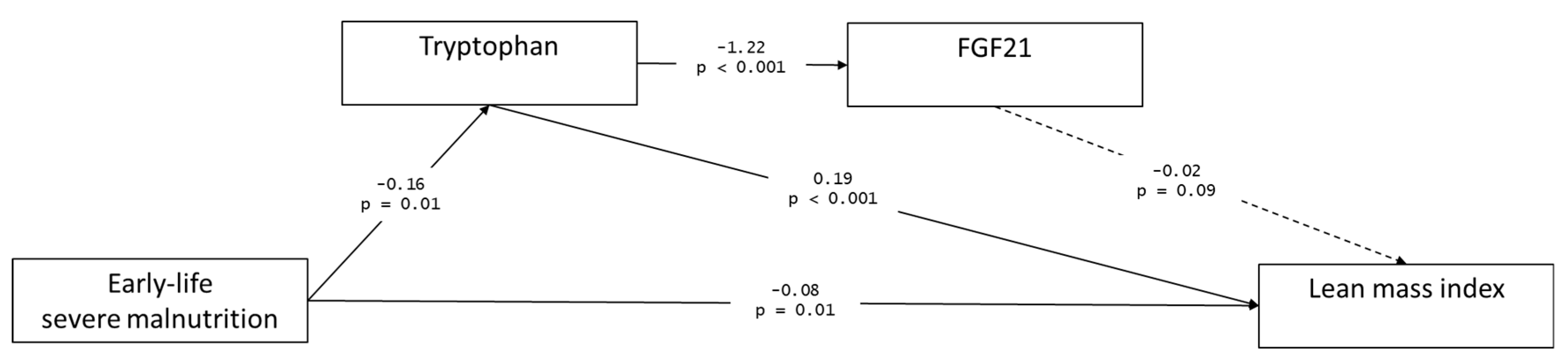
3.3. Path Analysis
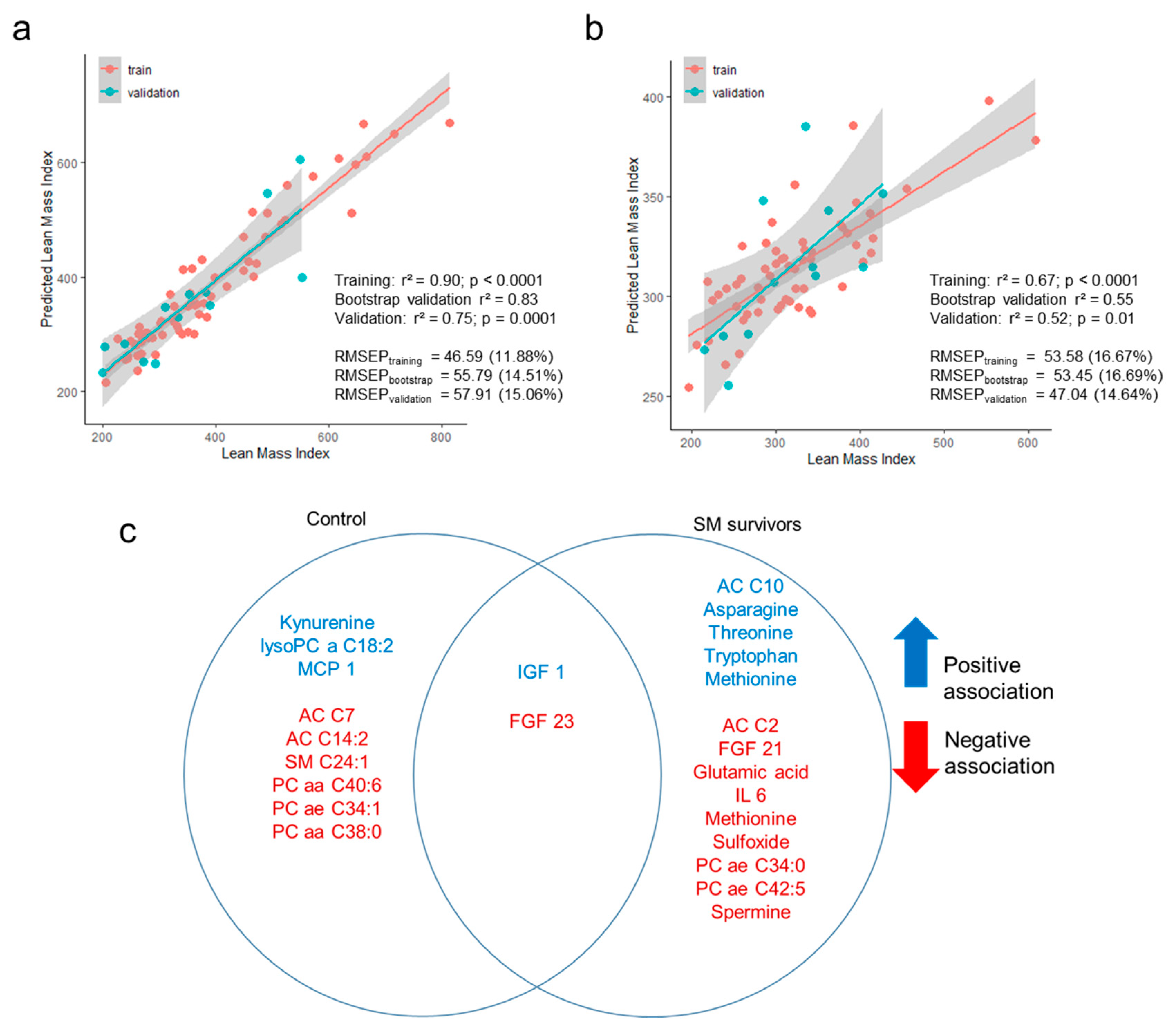
3.4. Prediction of Lean Mass Index Using Metabolome and Hormonal Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Nutrition Report. Available online: https://globalnutritionreport.org/reports/2020-global-nutrition-report/ (accessed on 19 May 2020).
- Bhutta, Z.A.; Berkley, J.A.; Bandsma, R.H.J.; Kerac, M.; Trehan, I.; Briend, A. Severe childhood malnutrition. Nat. Rev. Dis. Primers 2017, 3, 17067. [Google Scholar] [CrossRef] [PubMed]
- Kerac, M.; McGrath, M.; Connell, N.; Compala, C.; Moore, W.; Bailey, J.; Bandsma, R.; Berkley, J.; Briend, A.; Collins, S.; et al. Severe Malnutrition: Thinking deeply, communicating simply. BMJ Glob. Health 2020, in press. [Google Scholar]
- Lelijveld, N.; Seal, A.; Wells, J.C.; Kirkby, J.; Opondo, C.; Chimwezi, E.; Bunn, J.; Bandsma, R.; Heyderman, R.S.; Nyirenda, M.J.; et al. Chronic disease outcomes after severe acute malnutrition in Malawian children (ChroSAM): A cohort study. Lancet Glob. Health 2016, 4, e654–e662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis-Emmanuel, P.M.; Thompson, D.S.; Barnett, A.T.; Osmond, C.; Byrne, C.D.; Hanson, M.A.; Gluckman, P.D.; Forrester, T.E.; Boyne, M.S. Glucose metabolism in adult survivors of severe acute malnutrition. J. Clin. Endocrinol. Metab. 2014, 99, 2233–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennant, I.A.; Barnett, A.T.; Thompson, D.S.; Kips, J.; Boyne, M.S.; Chung, E.E.; Chung, A.P.; Osmond, C.; Hanson, M.A.; Gluckman, P.D.; et al. Impaired cardiovascular structure and function in adult survivors of severe acute malnutrition. Hypertension 2014, 64, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Grey, K.; Gonzales, G.B.; Abera, M.; Lelijveld, N.; Thompson, D.; Berhane, M.; Abdissa, A.; Girma, T.; Kerac, M. Severe malnutrition or famine exposure in childhood and cardiometabolic non-communicable disease later in life: A systematic review. under review.
- Wells, J.C.; Sawaya, A.L.; Wibaek, R.; Mwangome, M.; Poullas, M.S.; Yajnik, C.S.; Demaio, A. The double burden of malnutrition: Aetiological pathways and consequences for health. Lancet 2020, 395, 75–88. [Google Scholar] [CrossRef]
- Wells, J.C.K. The capacity-load model of non-communicable disease risk: Understanding the effects of child malnutrition, ethnicity and the social determinants of health. Eur. J. Clin. Nutr. 2018, 72, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Montagnese, C.; Nutile, T.; Marphatia, A.A.; Grijalva-Eternod, C.S.; Siervo, M.; Ciullo, M.; Wells, J.C. Body composition, leg length and blood pressure in a rural Italian population: A test of the capacity-load model. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1204–1212. [Google Scholar] [CrossRef]
- Unni, U.S.; Ramakrishnan, G.; Raj, T.; Kishore, R.P.; Thomas, T.; Vaz, M.; Kurpad, A.V. Muscle mass and functional correlates of insulin sensitivity in lean young Indian men. Eur. J. Clin. Nutr. 2009, 63, 1206–1212. [Google Scholar] [CrossRef]
- Wells, J.C.K. Body composition and susceptibility to type 2 diabetes: An evolutionary perspective. Eur. J. Clin. Nutr. 2017, 71, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Tang, L.; Chen, J.; Pei, J.; Chen, P.; Fang, Z.; Zhou, S.; Hu, X. Association of predicted lean body mass and fat mass with cardiovascular events in patients with type 2 diabetes mellitus. Can. Med. Assoc. J. 2019, 191, E1042–E1048. [Google Scholar] [CrossRef] [Green Version]
- Medina-Inojosa, J.R.; Somers, V.K.; Thomas, R.J.; Jean, N.; Jenkins, S.M.; Gomez-Ibarra, M.A.; Supervia, M.; Lopez-Jimenez, F. Association Between Adiposity and Lean Mass with Long-Term Cardiovascular Events in Patients With Coronary Artery Disease: No Paradox. J. Am. Heart Assocc. 2018, 7, e007505. [Google Scholar] [CrossRef] [Green Version]
- Hioki, H.; Miura, T.; Motoki, H.; Kobayashi, H.; Kobayashi, M.; Nakajima, H.; Kimura, H.; Mawatari, E.; Akanuma, H.; Sato, T.; et al. Lean body mass index prognostic value for cardiovascular events in patients with coronary artery disease. Heart Asia 2015, 7, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikanthan, P.; Horwich, T.B.; Tseng, C.H. Relation of Muscle Mass and Fat Mass to Cardiovascular Disease Mortality. Am. J. Cardiol. 2016, 117, 1355–1360. [Google Scholar] [CrossRef]
- Lavie, C.J.; De Schutter, A.; Patel, D.A.; Romero-Corral, A.; Artham, S.M.; Milani, R.V. Body composition and survival in stable coronary heart disease: Impact of lean mass index and body fat in the “obesity paradox”. J. Am. Coll. Cardiol. 2012, 60, 1374–1380. [Google Scholar] [CrossRef] [Green Version]
- Bourdon, C.; Lelijveld, N.; Thompson, D.; Dalvi, P.S.; Gonzales, G.B.; Wang, D.; Alipour, M.; Wine, E.; Chimwezi, E.; Wells, J.C.; et al. Metabolomics in plasma of Malawian children 7 years after surviving severe acute malnutrition: “ChroSAM” a cohort study. EBioMedicine 2019, 45, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.C.K.; Williams, J.E.; Fewtrell, M.; Singhal, A.; Lucas, A.; Cole, T.J. A simplified approach to analysing bio-electrical impedance data in epidemiological surveys. Int. J. Obes. 2007, 31, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Lee, T.; Maydeu-Olivares, A. Understanding the Model Size Effect on SEM Fit Indices. Educ. Psychol. Meas. 2018, 79, 310–334. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.H.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 22. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- Ninomiya, S.; Nakamura, N.; Nakamura, H.; Mizutani, T.; Kaneda, Y.; Yamaguchi, K.; Matsumoto, T.; Kitagawa, J.; Kanemura, N.; Shiraki, M.; et al. Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy. Nutrients 2020, 12, 978. [Google Scholar] [CrossRef] [Green Version]
- Cortamira, N.O.; Seve, B.; Lebreton, Y.; Ganier, P. Effect of dietary tryptophan on muscle, liver and whole-body protein synthesis in weaned piglets: Relationship to plasma insulin. Br. J. Nutr. 1991, 66, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.D.; Smith, T.K.; Bayley, H.S. A Role for Tryptophan in Regulation of Protein Synthesis in Porcine Muscle. J. Nutr. 1988, 118, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.A.; Moore, S.C.; Playdon, M.; Meirelles, O.; Newman, A.B.; Milijkovic, I.; Kritchevsky, S.B.; Schwartz, A.; Goodpaster, B.H.; Sampson, J.; et al. Metabolites Associated with Lean Mass and Adiposity in Older Black Men. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2017, 72, 1352–1359. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.M.; Maratos-Flier, E. Understanding the Physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.M.; Laeger, T.; Dehner, M.; Albarado, D.C.; Clarke, B.; Wanders, D.; Burke, S.J.; Collier, J.J.; Qualls-Creekmore, E.; Solon-Biet, S.M.; et al. FGF21 Signals Protein Status to the Brain and Adaptively Regulates Food Choice and Metabolism. Cell Rep. 2019, 27, 2934–2947.e3. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, Y.; Saito, K.; Nakazawa, K.; Konishi, M.; Itoh, N.; Hakuno, F.; Takahashi, S.; Kato, H.; Takenaka, A. Rapid increase in fibroblast growth factor 21 in protein malnutrition and its impact on growth and lipid metabolism. Br. J. Nutr. 2015, 114, 1410–1418. [Google Scholar] [CrossRef] [Green Version]
- Zapata, R.C.; Singh, A.; Pezeshki, A.; Chelikani, P.K. Tryptophan restriction partially recapitulates the age-dependent effects of total amino acid restriction on energy balance in diet-induced obese rats. J. Nutr. Biochem. 2019, 65, 115–127. [Google Scholar] [CrossRef]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig 2014, 124, 3913–3922. [Google Scholar] [CrossRef] [Green Version]
- Yap, Y.W.; Rusu, P.M.; Chan, A.Y.; Fam, B.C.; Jungmann, A.; Solon-Biet, S.M.; Barlow, C.K.; Creek, D.J.; Huang, C.; Schittenhelm, R.B.; et al. Restriction of essential amino acids dictates the systemic metabolic response to dietary protein dilution. Nat. Commun. 2020, 11, 2894. [Google Scholar] [CrossRef]
- Wells, J.C.; Fewtrell, M.S. Measuring body composition. Arch. Dis. Child. 2006, 91, 612–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerac, M.; Bunn, J.; Seal, A.; Thindwa, M.; Tomkins, A.; Sadler, K.; Bahwere, P.; Collins, S. Probiotics and prebiotics for severe acute malnutrition (PRONUT study): A double-blind efficacy randomised controlled trial in Malawi. Lancet 2009, 374, 136–144. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SM Survivors | Controls | Mean Difference ° | p | |
|---|---|---|---|---|
| n | 69 | 77 | ||
| Mean age in years (SD) * | 9.63 (1.63) | 9.95 (2.31) | −0.32 | 0.33 |
| n girls (%) | 30 (43%) | 34 (44%) | 1 | |
| n HIV status (%) | <0.001 | |||
| Negative | 44 (64%) | 46 (60%) | ||
| Positive | 20 (29%) | 4 (5%) | ||
| Unknown | 5 (7%) | 27 (35%) | ||
| Mean mid-upper arm circumference in cm (SD) * | 17.12 (1.64) | 18.15(2.30) | −1.03 | 0.002 |
| Mean height in cm (SD) * | 125.51 (8.90) | 129.57 (13.73) | −4.06 | 0.038 |
| Mean length-for-height z-score (SD) * | −1.68 (1.21) | −1.37 (1.00) | −0.31 | 0.090 |
| Mean weight-for-age z-score (SD) * | −1.44 (0.93) | −1.2 (0.88) | −0.24 | 0.210 |
| Mean body-mass index in kg/m2 (SD) * | 15.25 (1.33) | 15.77 (1.60) | −0.52 | 0.037 |
| Mean lean mass index in (1/Z) × 1000 (SD) * | 319.97 (75.36) | 378.87 (132.61) | −58.9 | 0.001 |
| Feature | Partial Coefficient * | p ° | Partial r2 □ | R2 ▪ | p ° Interaction § |
|---|---|---|---|---|---|
| Hormones | |||||
| IGF1 | 0.125 | <0.001 | 0.271 | 0.667 | - |
| FGF21 | −0.054 | 0.027 | 0.069 | 0.574 | 0.007 |
| Lipids | |||||
| AC C18 | −0.054 | 0.021 | 0.073 | 0.576 | - |
| AC C18:1 | −0.050 | 0.046 | 0.058 | 0.569 | - |
| PC ae C34:1 | −0.065 | 0.004 | 0.107 | 0.592 | - |
| PC aa C32:0 | −0.060 | 0.005 | 0.095 | 0.586 | - |
| PC aa C34:1 | −0.062 | 0.006 | 0.096 | 0.587 | - |
| PC aa C36:1 | −0.064 | 0.006 | 0.095 | 0.586 | - |
| PC aa C32:1 | −0.058 | 0.012 | 0.082 | 0.580 | - |
| PC ae C34:0 | −0.051 | 0.027 | 0.068 | 0.574 | - |
| PC ae C38:5 | −0.052 | 0.027 | 0.067 | 0.573 | - |
| Amino acids | |||||
| Tryptophan | 0.079 | <0.001 | 0.150 | 0.611 | 0.003 |
| Tyrosine | 0.057 | 0.012 | 0.082 | 0.580 | - |
| Citrulline | 0.052 | 0.040 | 0.061 | 0.571 | - |
| Biogenic amine | |||||
| Creatinine | 0.064 | 0.004 | 0.100 | 0.589 | - |
| Ratio and sums | |||||
| Kynurenine:Tryptophan | −0.07 | <0.001 | 0.003 | 0.590 | - |
| Sum of aromatic amino acids | 0.048 | 0.040 | 0.059 | 0.569 | - |
| Sum of lysine, threonine and tryptophan | 0.073 | 0.001 | 0.122 | 0.598 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzales, G.B.; Lelijveld, N.; Bourdon, C.; Chimwezi, E.; Nyirenda, M.J.; Wells, J.C.; Kerac, M.; Bandsma, R.H.J. Childhood Malnutrition and Association of Lean Mass with Metabolome and Hormone Profile in Later Life. Nutrients 2020, 12, 3593. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113593
Gonzales GB, Lelijveld N, Bourdon C, Chimwezi E, Nyirenda MJ, Wells JC, Kerac M, Bandsma RHJ. Childhood Malnutrition and Association of Lean Mass with Metabolome and Hormone Profile in Later Life. Nutrients. 2020; 12(11):3593. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113593
Chicago/Turabian StyleGonzales, Gerard Bryan, Natasha Lelijveld, Celine Bourdon, Emmanuel Chimwezi, Moffat J. Nyirenda, Jonathan C. Wells, Marko Kerac, and Robert H. J. Bandsma. 2020. "Childhood Malnutrition and Association of Lean Mass with Metabolome and Hormone Profile in Later Life" Nutrients 12, no. 11: 3593. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113593