
Asprosin—A Fasting-Induced, Glucogenic, and Orexigenic Adipokine as a New Promising Player. Will It Be a New Factor in the Treatment of Obesity, Diabetes, or Infertility? A Review of the Literature


Abstract
:1. Introduction
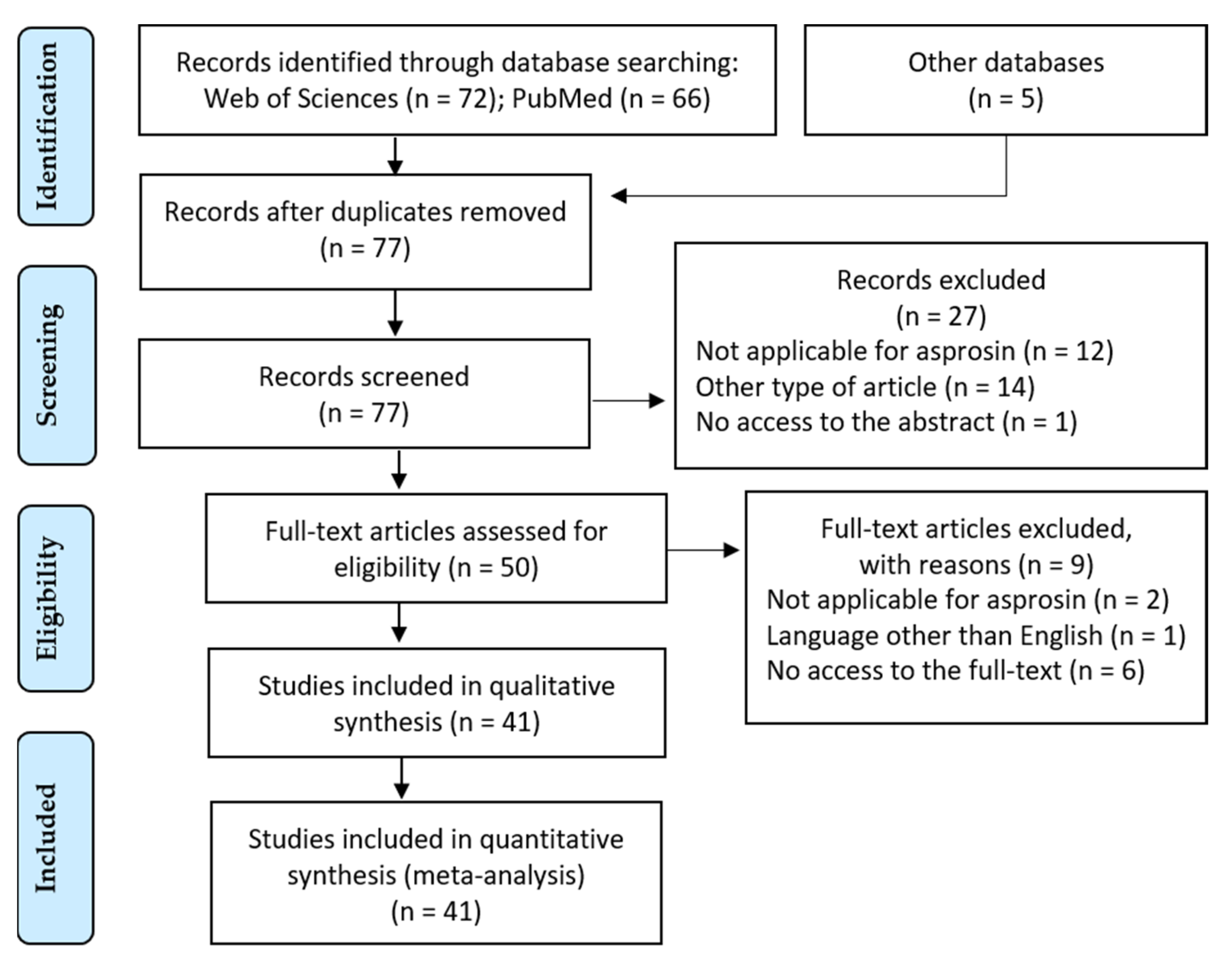
2. Materials and Methods
3. Results
4. Asprosin Discovery and Structure
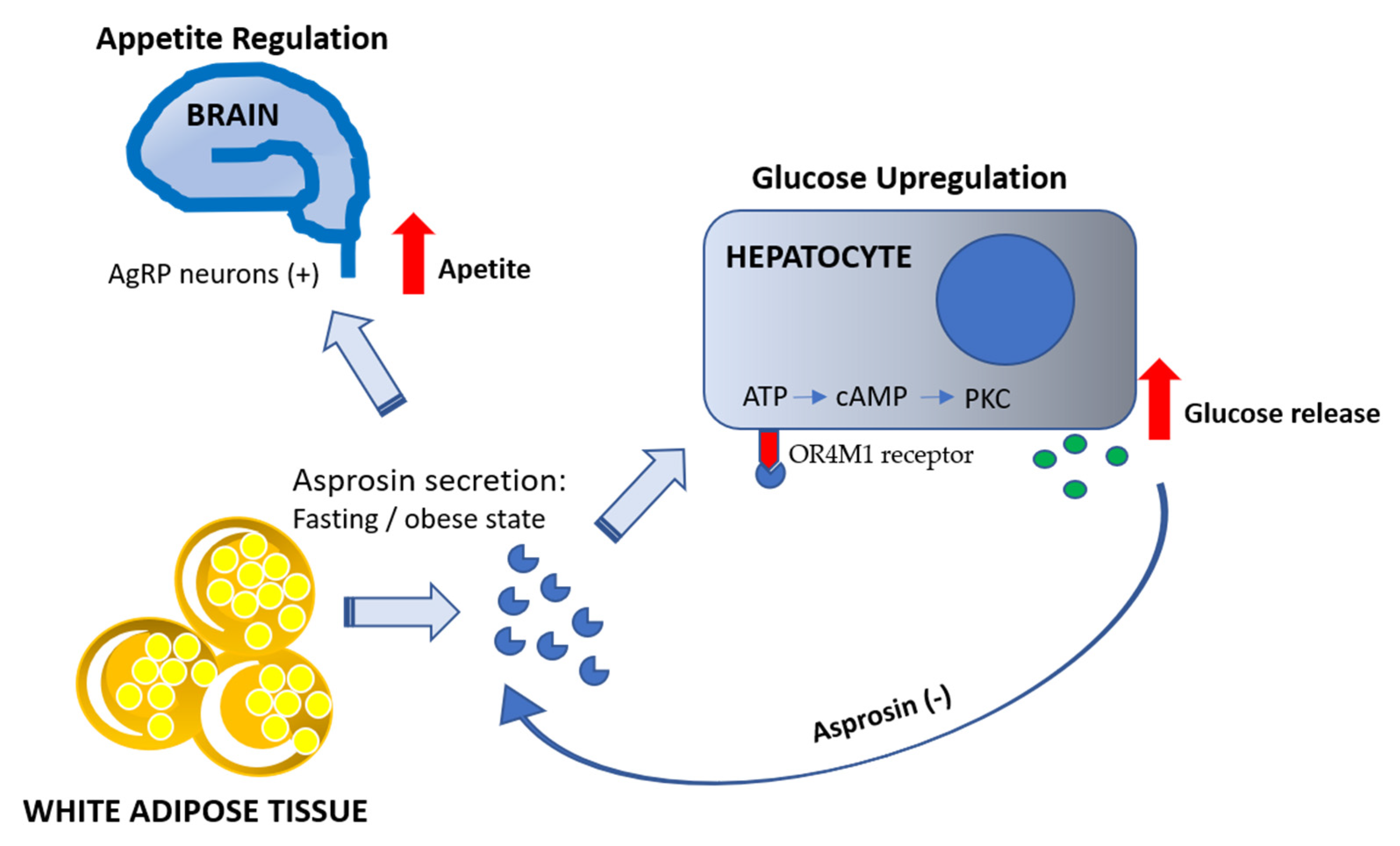
5. Glucogenic Action of Asprosin
6. Asprosin and Appetite
7. Asprosin and Insulin Resistance, Diabetes, and Obesity
8. Asprosin in Myocardial and Microvascular Damage
9. Asprosin and Male/Female Fertility
10. Asprosin and Polycystic Ovary Syndrome
11. Physical Activity and Asprosin Production
12. Research on the Potential of Asprosin as a Drug—Promising Research Directions
13. Conclusions
Funding
Conflicts of Interest
References
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Korta, P.; Pocheć, E.; Mazur-Biały, A. Irisin as a Multifunctional Protein: Implications for Health and Certain Diseases. Medicina 2019, 55, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münzberg, H.; Morrison, C.D. Structure, production and signaling of leptin. Metabolism 2015, 64, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the past two decades. J. Mol. Cell. Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef]
- Romere, C.; Duerrschmid, C.; Bournat, J.; Constable, P.; Jain, M.; Xia, F.; Saha, P.K.; Del Solar, M.; Zhu, B.; York, B.; et al. Asprosin, a fasting-induced glucogenic protein hormone. Cell 2016, 165, 566–579. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.G.; Xie, W.; Chopra, A.R. Energy Regulation Mechanism and Therapeutic Potential of Asprosin. Diabetes 2020, 69, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Duerrschmid, C.; He, Y.; Wang, C.; Li, C.; Bournat, J.C.; Romere, C.; Saha, P.K.; Lee, M.E.; Phillips, K.J.; Jain, M.; et al. Asprosin is a centrally acting orexigenic hormone. Nat. Med. 2017, 23, 1444–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Yao, B.; Yang, Q.; Deng, J.; Song, Y.; Sui, T.; Zhou, L.; Yao, H.; Xu, Y.; Ouyang, H.; et al. Truncated C-terminus of fibrillin-1 induces Marfanoid-progeroid-lipodystrophy (MPL) syndrome in rabbit (biologists.org). Dis. Model Mech. 2018, 11, dmm031542. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Duan, L.; Gong, F.; Zhang, Y.; Deng, K.; Yao, Y.; Wang, L.; Pan, H.; Zhu, H. Serum Levels of Asprosin, a Novel Adipokine, Are Significantly Lowered in Patients with Acromegaly. Int. J. Endocrinol. 2020. [Google Scholar] [CrossRef]
- Wiecek, M.; Szymura, J.; Maciejczyk, M.; Kantorowicz, M.; Szygula, Z. Acute anaerobic exercise affects the secretion of asprosin, irisin, and other cytokines–a comparison between sexes. Front Physiol. 2018, 9, 1782. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Shan, H.; Chen, L.; Long, A.; Zhang, Y.; Liu, Y.; Jia, L.; Wei, F.; Han, J.; Li, T.; et al. OLFR734 Mediates Glucose Metabolism as a Receptor of Asprosin. Cell Metabol. 2019, 30, 319–328. [Google Scholar] [CrossRef]
- Ramnanan, C.J.; Edgerton, D.S.; Kraft, G.; Cherrington, A.D. Physiologic action of glucagon on liver glucose metabolism. Diabetes Obes. Metab. 2011, 13, 118–125. [Google Scholar] [CrossRef]
- Gropp, E.; Shanabrough, M.; Borok, E.; Xu, A.W.; Janoschek, R.; Buch, T.; Plum, N.; Hampel, B.; Weisman, A.; Barsh, G.S.; et al. Agouti-related peptide-expressing neurons are mandatory for feeding. Nat. Neurosci. 2015, 8, 1289–1291. [Google Scholar] [CrossRef]
- Morton, G.J.; Meek, T.H.; Schwartz, M.W. Neurobiology of food intake in health and disease. Nat. Rev. Neurosci. 2014, 15, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Beutler, L.R.; Knight, Z.A. A spotlight on appetite. Neuron 2018, 97, 739–741. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xu, Y.; Zheng, Y.; Kang, Q.; Lou, Z.; Liu, Q.; Chen, H.; Ji, Y.; Guo, L.; Chen, C.; et al. Increased plasma asprosin levels in patients with drug-naive anorexia nervosa. Eat Weight Disord. 2020. [Google Scholar] [CrossRef]
- Du, C.; Wang, C.; Guan, X.; Li, J.; Du, X.; Xu, Z.; Zheng, Z. Asprosin is associated with anorexia and body fat mass in cancer patients. Supportive Care Cancer 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Lin, T.A.; Liu, K.H.; Liao, C.H.; Liu, Y.Y.; Wu, V.C.; Wen, M.S.; Yeh, T.S. Serum asprosin levels and bariatric surgery outcomes in obese adults. Int. J. Obes. (Lond.) 2019, 43, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Ugur, K.; Aydin, S. Saliva and Blood Asprosin Hormone Concentration Associated with Obesity. Int. J. Endocrinol. 2019, 2521096. [Google Scholar] [CrossRef]
- Wang, Y.; Qu, H.; Xiong, X.; Qiu, Y.; Liao, Y.; Chen, Y.; Zheng, Y.; Zheng, H. Plasma Asprosin Concentrations Are Increased in Individuals with Glucose Dysregulation and Correlated with Insulin Resistance and First-Phase Insulin Secretion. Mediat. Inflamm. 2018, 2018, 9471583. [Google Scholar] [CrossRef] [Green Version]
- Alan, M.; Gurlek, B.; Yilmaz, A.; Aksit, M.; Aslanipour, B.; Gulhan, I.; Mechmet, C.; Taner, C.E. Asprosin: A novel peptide hormone related to insulin resistance in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2019, 35, 220–223. [Google Scholar] [CrossRef]
- Groener, J.B.; Valkanou, A.; Kender, Z.; Pfeiffenberger, J.; Kihm, L.; Fleming, T.; Nawroth, P.P.; Kopf, S. Asprosin response in hypoglycemia is not related to hypoglycemia unawareness but rather to insulin resistance in type 1 diabetes. PLoS ONE 2019, 14, e0222771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, C.; Zhou, N.; Fu, Y.; Cheng, X. Circulating asprosin concentrations are increased in type 2 diabetes mellitus and independently associated with fasting glucose and triglyceride. Clin. Chim. Acta 2017, 489, 183–188. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, H.; Ma, X.; Wu, H. Increased serum level and impaired response to glucose fluctuation of asprosin is associated with type 2 diabetes mellitus. J. Diabetes Investig. 2019, 11, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Naiemian, S.; Naeemipour, M.; Zarei, M.; Najafi, M.L.; Gohari, A.; Behroozikhah, M.R.; Heydari, H.; Miri, M. Serum concentration of asprosin in new-onset type 2 diabetes. Diabetol. Metab. Syndr. 2020, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Kocaman, N.; Kuloğlu, T. Expression of asprosin in rat hepatic, renal, heart, gastric, testicular and brain tissues and its changes in a streptozotocin-induced diabetes mellitus model. Tissue Cell 2020, 66, 101397. [Google Scholar] [CrossRef]
- Lee, T.; Yun, S.; Jeong, J.H.; Jung, T.W. Asprosin impairs insulin secretion in response to glucose and viability through TLR4/JNK-mediated inflammation. Mol. Cell. Endocrinol. 2019, 486, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.W.; Kim, H.C.; Kim, H.U.; Park, T.; Park, J.; Kim, U.; Jeong, J.H. Asprosin attenuates insulin signaling pathway through PKCδ-activated ER stress and inflammation in skeletal muscle. J. Cell Physiol. 2019, 234, 20888–20899. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhao, L.; Guo, C.; Yang, L.; Wang, D.; Li, Y.; Xia, H.; Wang, C.; Cai, Z.; Li, L.; et al. Higher Serum Asprosin Level is Associated with Urinary Albumin Excretion and Renal Function in Type 2 Diabetes. Diabetes Metabol. Syndr. Obes. 2020, 13, 4341–4351. [Google Scholar] [CrossRef]
- Ke, F.; Xue, G.; Jiang, X.; Li, F.; Lai, X.; Zhang, M.; Shen, Y.; Gao, L. Combination of asprosin and adiponectin as a novel marker for diagnosing non-alcoholic fatty liver disease. Cytokine 2020, 134, 155184. [Google Scholar] [CrossRef]
- Zhong, L.; Long, Y.; Wang, S.; Lian, R.; Deng, L.; Ye, Z.; Wang, Z.; Liu, B. Continuous elevation of plasma asprosin in pregnant women complicated with gestational diabetes mellitus: A nested case-control study. Placenta 2020, 93, 17–22. [Google Scholar] [CrossRef]
- Baykus, Y.; Yavuzkir, S.; Ustebay, S.; Ugur, K.; Deniz, R.; Aydin, S. Asprosin in umbilical cord of newborns and maternal blood of gestational diabetes, preeclampsia, severe preeclampsia, intrauterine growth retardation and macrosemic fetus. Peptides 2019, 120, 170132. [Google Scholar] [CrossRef]
- Sünnetçi, S.E.; Hatipoğlu, H.U. Increased serum circulating asprosin levels in children with obesity. Pediatr. Int. 2020, 62, 467–476. [Google Scholar] [CrossRef]
- Janoschek, R.; Hoffmann, T.; Morcos, Y.A.T.; Sengle, G.; Dötsch, J.; Hucklenbruch-Rother, E. Asprosin in pregnancy and childhood. Mol. Cell. Pediatr. 2020, 7, 1–5. [Google Scholar] [CrossRef]
- Zhang, Z.; Tan, Y.; Zhu, L.; Zhang, B.; Feng, P.; Gao, E.; Hu, C.; Wang, X.; Yi, W.; Sun, Y. Asprosin improves the survival of mesenchymal stromal cells in myocardial infarction by inhibiting apoptosis via the activated ERK1/2-SOD2 pathway. Life Sci. 2019, 231, 116554. [Google Scholar] [CrossRef]
- Wen, M.S.; Wang, C.Y.; Yeh, J.K.; Chen, C.C.; Tsai, M.L.; Ho, M.Y.; Hung, K.C.; Hsieh, I.C. The role of Asprosin in patients with dilated cardiomyopathy. BMC Cardiovasc. Disord. 2020, 20, 402. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Qiu, C.M.; Hou, J.N.; Wei, X.Y.; Xiang, C.X.; Pei, H.F. Study of the role and mechanism of asprosin/spartin pathway in cardiac microvascular endothelial injury induced by diabete mellitus. Sichuan Da Xue Xue Bao Yi Xue Ban 2019, 50, 827–834. (In Chinese) [Google Scholar]
- Wei, F.; Long, A.; Wang, Y. The Asprosin-OLFR734 hormonal signaling axis modulates male fertility. Cell Discov. 2019, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liao, M.; Shen, R.; Zhang, L.; Hu, H.; Wu, J.; Wang, X.; Qu, H.; Guo, S.; Long, M.; et al. Plasma asprosin levels are associated with glucose metabolism, lipid, and sex hormone profiles in females with metabolic-related diseases. Mediat. Inflamm. 2018, 2018, 7375294. [Google Scholar] [CrossRef] [Green Version]
- Cooper, T.G.; Noonan, E.; von Eckardstein, S.; Auger, J.; Gordon Baker, H.W.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization reference values for human semen characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef]
- Irvine, D.S. Epidemiology and aetiology of male infertility. Hum. Reprod. 1998, 13 (Suppl. 1), 33–44. [Google Scholar] [CrossRef] [Green Version]
- Maylem, E.R.S.; Spicer, L.J.; Batalha, I.; Schutz, L.F. Discovery of a possible role of asprosin in ovarian follicular function. J. Mol. Endocrinol. 2021, 66, 25–34. [Google Scholar] [CrossRef]
- Leonard, A.N.; Shill, A.L.; Thackray, A.E.; Stensel, D.J.; Bishop, N.C. Fasted plasma asprosin concentrations are associated with menstrual cycle phase, oral contraceptive use and training status in healthy women. Eur. J. Appl. Physiol. 2020, 1–9. [Google Scholar] [CrossRef]
- Shah, R. Emerging Topics in Cardiometabolic and Psychologic Sequelae, Pathogenesis, and Treatment of Polycystic Ovarian Syndrome: A Review. Children 2019, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.L.; Huang, S.Y.; Hsu, Y.C.; Chin, T.H.; Soong, Y.K. The serum level of irisin, but not asprosin, is abnormal in polycystic ovary syndrome patients. Sci. Rep. 2019, 9, 6447. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.R.; Seo, D.Y.; Kim, T.N.; Park, S.H.; Kwak, H.-B.; Ko, K.S.; Rhee, B.D.; Han, J. Aerobic Exercise Training Decreases Hepatic Asprosin in Diabetic Rats. J. Clin. Med. 2019, 8, 666. [Google Scholar] [CrossRef] [Green Version]
- Nakhaei, H.; Mogharnasi, M.; Fanaei, H. Effect of swimming training on levels of asprosin, lipid profile, glucose and insulin resistance in rats with metabolic syndrome. Obes. Med. 2019, 15, 100111. [Google Scholar] [CrossRef]
- Ceylan, H.İ.; Saygın, Ö.; Özel Türkcü, Ü. Assessment of acute aerobic exercise in the morning versus evening on asprosin, spexin, lipocalin-2, and insulin level in overweight/obese versus normal weight adult men. Chronobiol. Int. 2020, 37, 1252–1268. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, H.İ.; Saygın, Ö. An investigation of the relationship between new fasting hormone asprosin, obesity and acute–chronic exercise: Current systematic review. Arch. Physiol. Biochem. 2020, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wiecek, M.; Szymura, J.; Sproull, J.; Szygula, Z. Decreased Blood Asprosin in Hyperglycemic Menopausal Women as a Result of Whole-Body Cryotherapy Regardless of Metabolic Syndrome. J. Clin. Med. 2019, 8, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, A.; Moore, D.D. Methods for Stimulation of Appetite and Increase in Weight by Administration of Asprosin. U.S. Patent Application No. 20,190,282,664, 19 September 2019. Available online: http://patents.justia.com/20190282664 (accessed on 31 January 2021).
- Chopra, A.; Moore, D.D. Asprosin, a Fast-Induced Glucogenic Protein Hormone. U.S. Patent Application No. 20,190,144,536, 16 May 2019. Available online: http://patents.justia.com/20190144536 (accessed on 31 January 2021).
- CN107261115A—Asprosin is Used for the Application for Preparing Treatment Ischemic Heart Medicine—Patent Application no. CN107261115, 7 July 2020. Available online: http://patents.google.com/cn10721115a/en (accessed on 31 January 2021). (In Chinese).



{kind=link}
{kind=link}
| Thematic Threads | Description | References |
|---|---|---|
| Asprosin discovery | Asprosin structure | [7,8] |
| Asprosin action | Olfr734 as an asprosin receptor | [12] |
| FBN1 gene mutation reduces asprosin production | [7,9,10] | |
| Starvation induces asprosin production by white adipose tissue | [7] | |
| Obesity state induces asprosin production by white adipose tissue | [13] | |
| Mechanism of asprosin action via cAMP/PKC activation | [7] | |
| Liver as a target organ for asprosin action—induction of glucose release | [7,9] | |
| Appetite regulation | Asprosin activates orexigenic agouti-related peptide (AgRP) neurons leading to appetite stimulation by the Gαs-cAMP-PKA axis | [9] |
| Anti-asprosin antibodies efficiently suppress AgRP neurons activation and food intake by insulin-resistant obese mice | [9] | |
| Elevated asprosin level in diseases state | in patients with obesity | [20,21,35,36] |
| in patients with insulin resistance | [22,23] | |
| in patients with diabetes mellitus type 1 | [24] | |
| in patients with diabetes mellitus type 2 | [25,26,27] | |
| in patients with anorexia nervosa | [18] | |
| in pregnant women with gestational diabetes mellitus | [33] | |
| in pregnant women with pre-eclampsia | [34] | |
| Decreased asprosin level in diseases state | patients with neonatal progeroid syndrome (NPS) | [7,8] |
| in oncological patients with anorexia | [19] | |
| in women with intrauterine growth restriction | [34] | |
| in patients with acromegaly | [11] | |
| In mice with streptozocin-induces diabetes | [28] | |
| Pathophysiology of asprosin action | Asprosin induces pancreatic beta-cell dysfunction and impairs insulin release | [29] |
| Asprosin impairs muscle cell sensitivity to insulin | [30] | |
| Cardioprotection | Asprosin improves the effectiveness of mesenchymal stromal cells (MSCs) use in myocardial infarction (MI) treatment | [37] |
| Asprosin protects cardiomyocytes through hypoxia-induced apoptosis | [38] | |
| Asprosin protects cardiac microvascular endothelial cells against damage caused by high glucose concentration | [39] | |
| Fertility | Asprosin improves progressive sperm motility and fertility potential | [40] |
| Asprosin regulates the function of ovarian follicles | [44] | |
| Asprosin level changes in the course of menstrual cycle | [45] | |
| Physical activity in asprosin reduction | 8-week aerobic exercise program reduces asprosin in mice with type 1 diabetes mellitus | [48] |
| Intermittent and continuous swimming training decrease asprosin levels in rats with metabolic syndrome | [49] | |
| Acute aerobic exercise reduces asprosin level in healthy and obese men | [50] | |
| Short-term high-intensive anaerobic exercise increases asprosin level in healthy women | [12] | |
| Whole-body cryotherapy reduces asprosin level in menopausal women | [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur-Bialy, A.I. Asprosin—A Fasting-Induced, Glucogenic, and Orexigenic Adipokine as a New Promising Player. Will It Be a New Factor in the Treatment of Obesity, Diabetes, or Infertility? A Review of the Literature. Nutrients 2021, 13, 620. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020620
Mazur-Bialy AI. Asprosin—A Fasting-Induced, Glucogenic, and Orexigenic Adipokine as a New Promising Player. Will It Be a New Factor in the Treatment of Obesity, Diabetes, or Infertility? A Review of the Literature. Nutrients. 2021; 13(2):620. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020620
Chicago/Turabian StyleMazur-Bialy, Agnieszka Irena. 2021. "Asprosin—A Fasting-Induced, Glucogenic, and Orexigenic Adipokine as a New Promising Player. Will It Be a New Factor in the Treatment of Obesity, Diabetes, or Infertility? A Review of the Literature" Nutrients 13, no. 2: 620. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020620