
Vitamin D Deficiency Cause Gender Specific Alterations of Renal Arterial Function in a Rodent Model

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Chronic Treatment of the Rats
2.4. Myography
2.5. Immunohistochemistry
2.6. Statistics
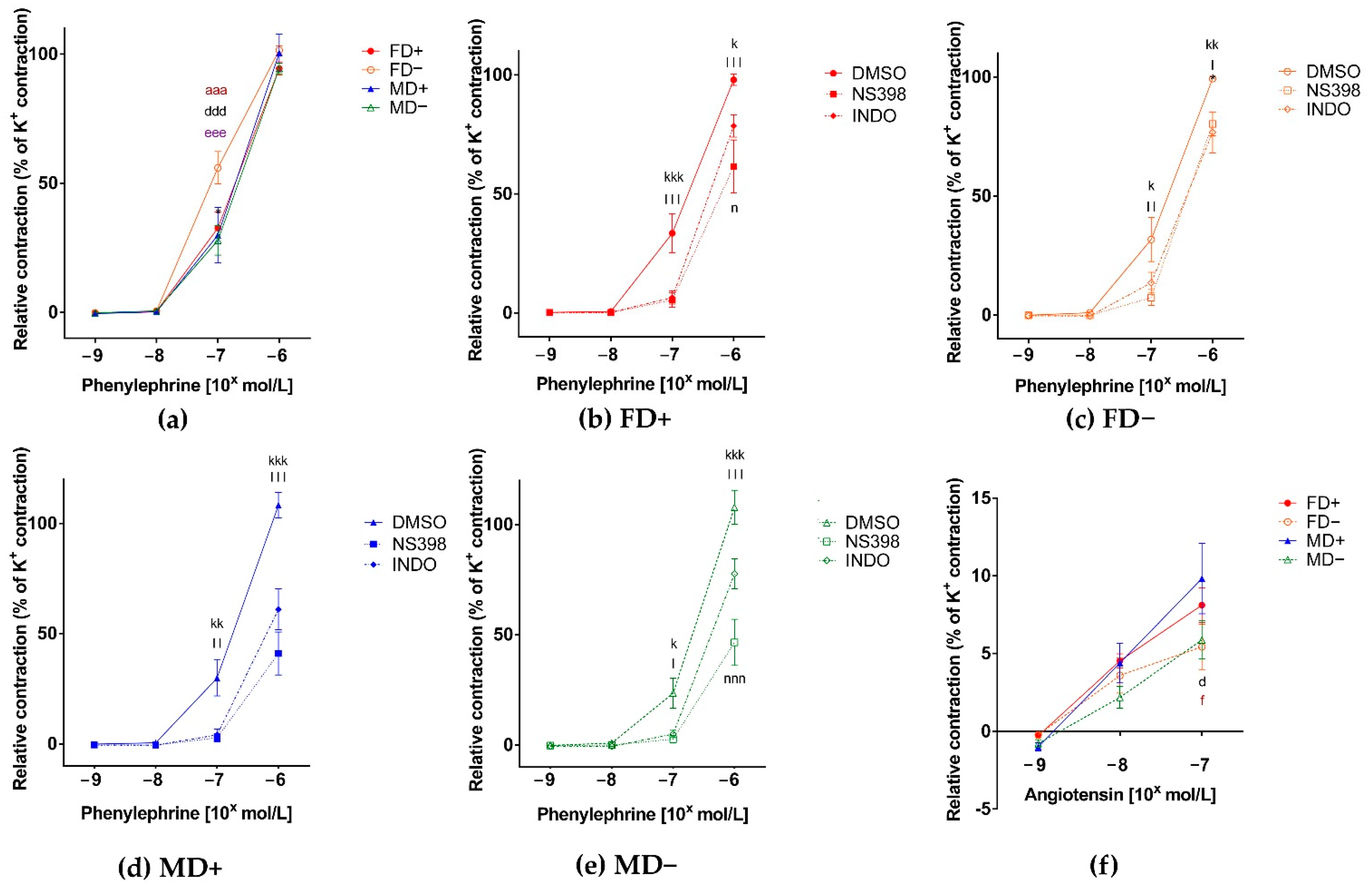
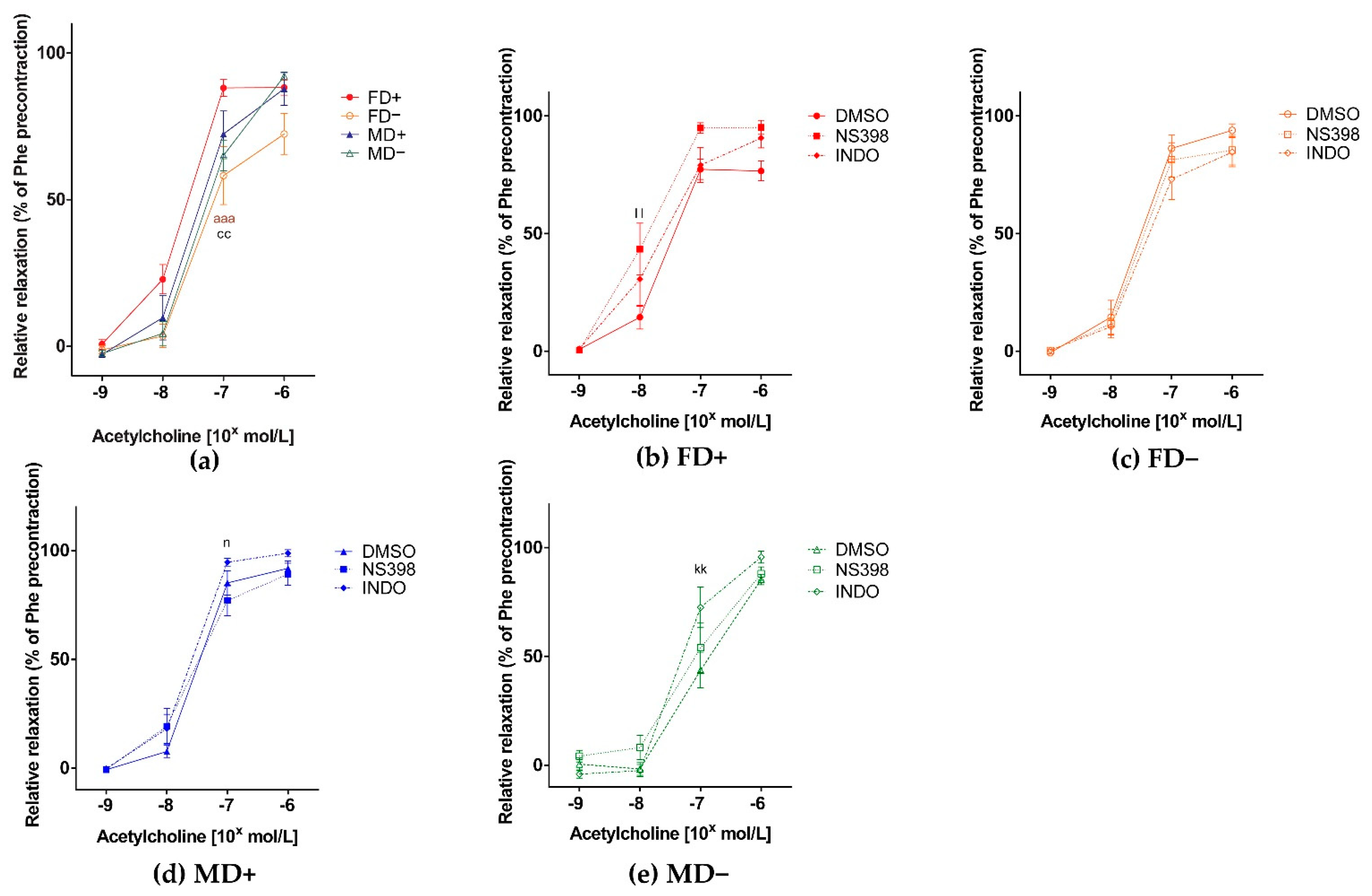
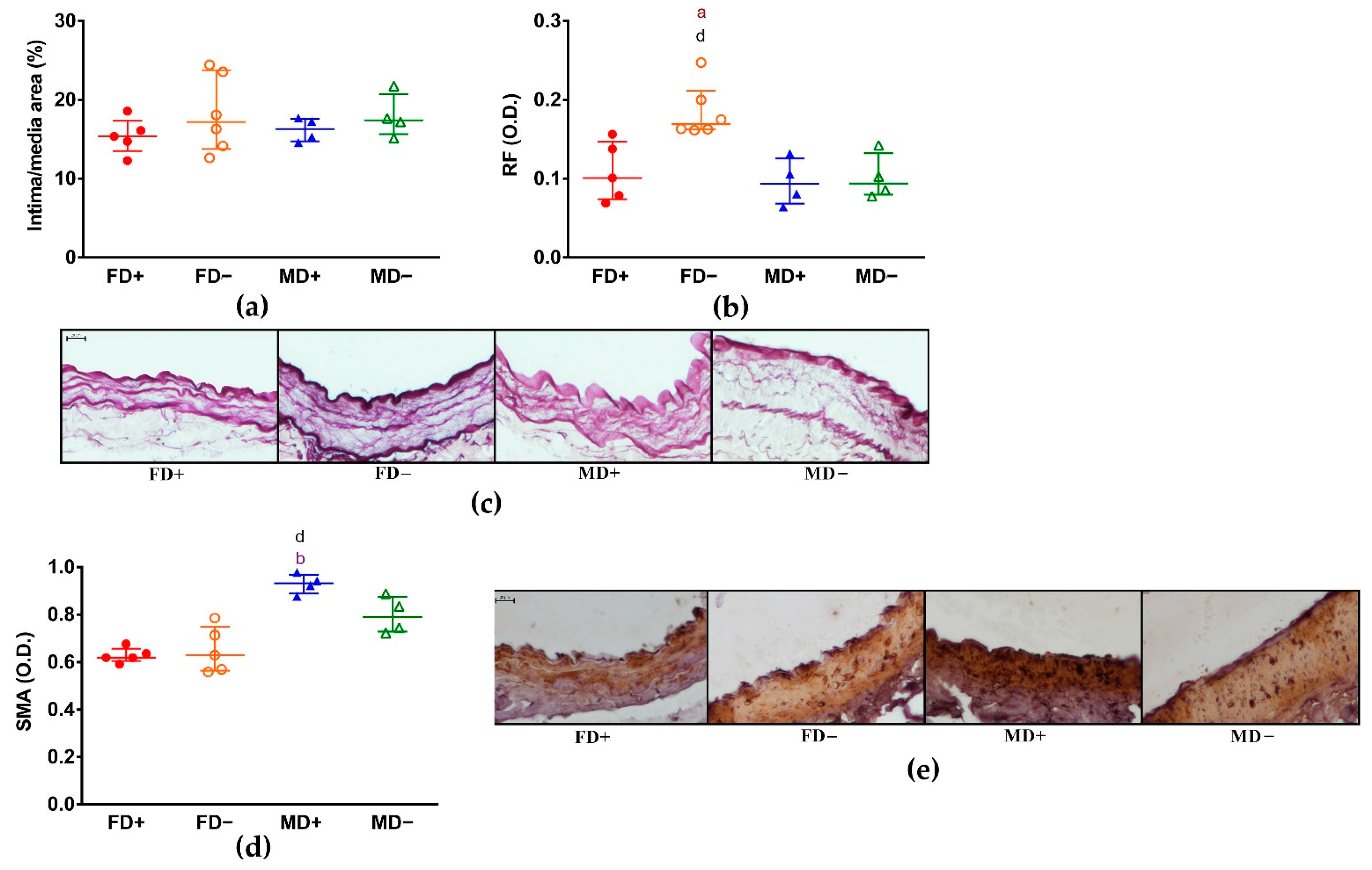
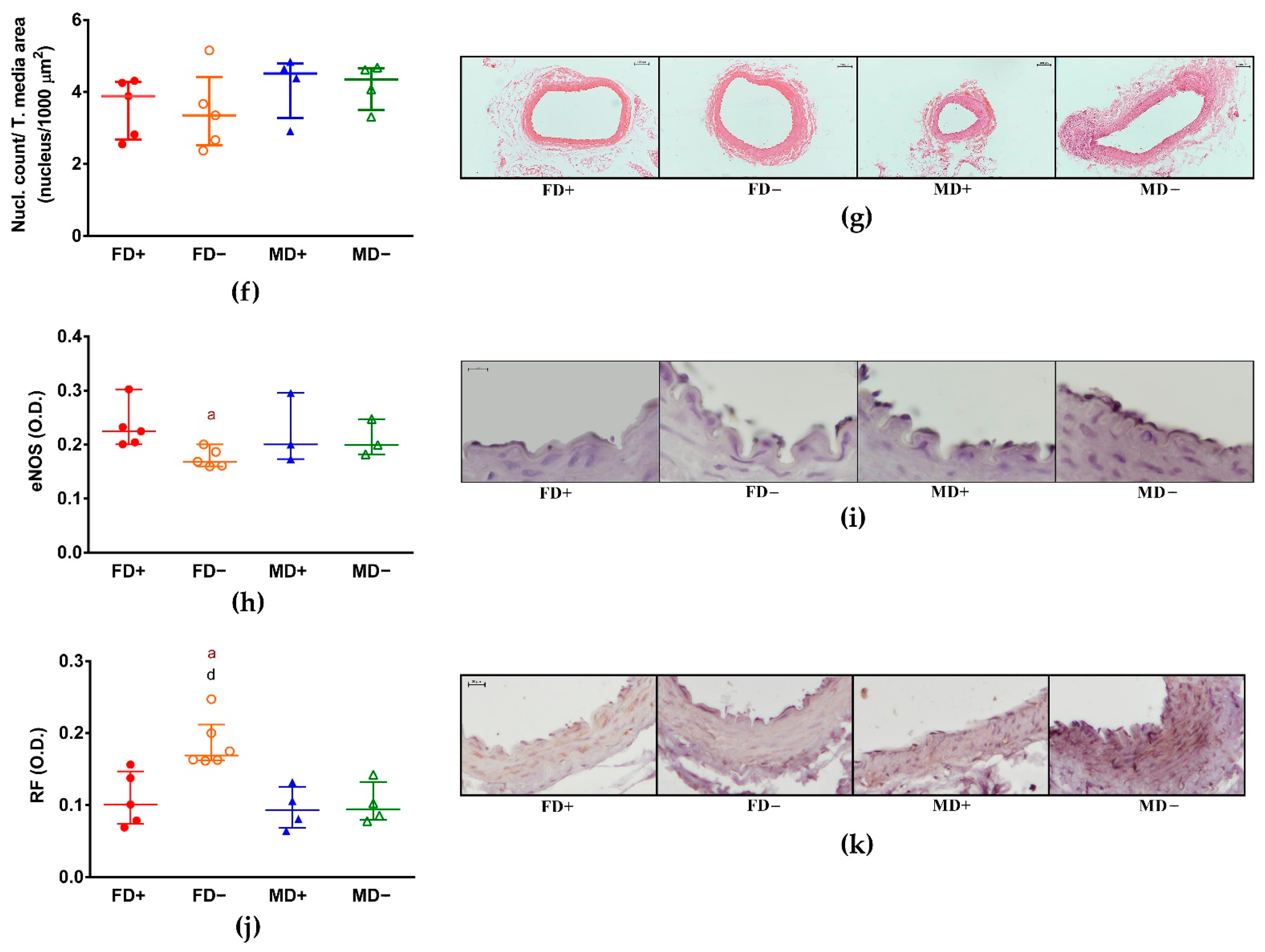
3. Results
3.1. Vascular Function of Renal Arteries
3.2. Histology of Renal Arteries
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Theodoratou, E.; Tzoulaki, I.; Zgaga, L.; Ioannidis, J.P.A. Vitamin D and multiple health outcomes: Umbrella review of systematic reviews and meta-analyses of observational studies and randomised trials. BMJ 2014, 348, g2035. [Google Scholar] [CrossRef] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.-M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- AlQuaiz, A.M.; Kazi, A.; Fouda, M.; Alyousefi, N. Age and gender differences in the prevalence and correlates of vitamin D deficiency. Arch. Osteoporos. 2018, 13, 49. [Google Scholar] [CrossRef]
- Sanghera, D.K.; Sapkota, B.R.; Aston, C.E.; Blackett, P.R. Vitamin D Status, Gender Differences, and Cardiometabolic Health Disparities. Ann. Nutr. Metab. 2017, 70, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, S.; Braun, J.; Bopp, M.; Faeh, D. Inverse association between circulating vitamin D and mortality—Dependent on sex and cause of death? Nutr. Metab. Cardiovasc. Dis. 2013, 23, 960–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, A.; Herrmann, S.M.; Textor, S.C. Chronic Renal Ischemia in Humans: Can Cell Therapy Repair the Kidney in Occlusive Renovascular Disease? Physiology 2015, 30, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Pál, É.; Hadjadj, L.; Fontányi, Z.; Monori-Kiss, A.; Mezei, Z.; Lippai, N.; Magyar, A.; Heinzlmann, A.; Karvaly, G.; Monos, E.; et al. Vitamin D deficiency causes inward hypertrophic remodeling and alters vascular reactivity of rat cerebral arterioles. PLoS ONE 2018, 13, e0192480. [Google Scholar] [CrossRef]
- Pál, É.; Hadjadj, L.; Fontányi, Z.; Monori-Kiss, A.; Lippai, N.; Horváth, E.M.; Magyar, A.; Monos, E.; Nádasy, G.L.; Benyó, Z.; et al. Gender, hyperandrogenism and vitamin D deficiency related functional and morphological alterations of rat cerebral arteries. PLoS ONE 2019, 14, e0216951. [Google Scholar] [CrossRef]
- Hadjadj, L.; Várbíró, S.; Horváth, E.M.; Monori-Kiss, A.; Pál, É.; Karvaly, G.B.; Heinzlmann, A.; Magyar, A.; Szabo, I.; Sziva, R.E.; et al. Insulin resistance in an animal model of polycystic ovary disease is aggravated by vitamin D deficiency: Vascular consequences. Diabetes Vasc. Dis. Res. 2018, 15, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Sziva, R.E.; Fontányi, Z.; Pál, É.; Hadjadj, L.; Monori-Kiss, A.; Horváth, E.M.; Benkő, R.; Magyar, A.; Heinzlmann, A.; Benyó, Z.; et al. Vitamin D Deficiency Induces Elevated Oxidative and Biomechanical Damage in Coronary Arterioles in Male Rats. Antioxidants 2020, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.S.; Blümel, J.E.; Arteaga, E.; Aedo, S.; Tapia, V.; Araos, A.; Sciaraffia, C.; Castelo-Branco, C. Gender differences in the prevalence of vitamin D deficiency in a southern Latin American country: A pilot study. Climacteric 2020, 23, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.K.; Hofsø, D.; Aasheim, E.T.; Tanbo, T.; Holven, K.B.; Andersen, L.F.; Røislien, J.; Hjelmesæth, J. Impact of gender on vitamin D deficiency in morbidly obese patients: A cross-sectional study. Eur. J. Clin. Nutr. 2011, 66, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Saki, F.; Kasaee, S.R.; Sadeghian, F.; Koohpeyma, F.; Omrani, G.R. Investigating the effect of testosterone by itself and in combination with letrozole on 1,25-dihydroxy vitamin D and FGF23 in male rats. J. Endocrinol. Investig. 2018, 42, 19–25. [Google Scholar] [CrossRef]
- Lee, S.R.; Park, M.-Y.; Yang, H.; Lee, G.-S.; An, B.-S.; Park, B.-K.; Jeung, E.-B.; Hong, E.-J. 5α-dihydrotestosterone reduces renal Cyp24a1 expression via suppression of progesterone receptor. J. Mol. Endocrinol. 2018, 60, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Wong, S.L.; Lau, C.W.; Lee, H.K.; Ng, C.F.; Zhang, L.; Yao, X.; Chen, Z.Y.; Vanhoutte, P.M.; Huang, Y. Calcitriol protects renovascular function in hypertension by down-regulating angiotensin II type 1 receptors and reducing oxidative stress. Eur. Hear. J. 2012, 33, 2980–2990. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, G.; Tucker, A.T.; Harwood, S.M.; Pearse, R.M.; Raftery, M.J.; Yaqoob, M.M. Ergocalciferol and Microcirculatory Function in Chronic Kidney Disease and Concomitant Vitamin D Deficiency: An Exploratory, Double Blind, Randomised Controlled Trial. PLoS ONE 2014, 9, e99461. [Google Scholar] [CrossRef]
- Arfian, N.; Kusuma, M.H.H.; Anggorowati, N.; Nugroho, D.B.; Jeffilano, A.; Suzuki, Y.; Ikeda, K.; Emoto, N. Vitamin D Upregulates Endothelin-1, ETBR, eNOS mRNA Expression and Attenuates Vascular Remodelling and Ischemia in Kidney Fibrosis Model in Mice Vitamin D Upregulates Endothelin-1, ETBR, eNOS mRNA Expression and Attenuates Vascular Remodelling and Ischemia in Kidney Fibrosis Model in Mice. Physiol. Res. 2018, 67, S137–S147. [Google Scholar] [CrossRef]
- Martínez-Miguel, P.; Valdivielso, J.M.; Medrano-Andrés, D.; Román-García, P.; Cano-Peñalver, J.L.; Rodríguez-Puyol, M.; Rodríguez-Puyol, D.; López-Ongil, S. The active form of vitamin D, calcitriol, induces a complex dual upregulation of endothelin and nitric oxide in cultured endothelial cells. Am. J. Physiol. Metab. 2014, 307, E1085–E1096. [Google Scholar] [CrossRef] [Green Version]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-H.; Meza, C.A.; Clarke, H.; Kim, J.-S.; Hickner, R.C. Vitamin D and Endothelial Function. Nutrients 2020, 12, 575. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, H. Vitamin D is a membrane antioxidant Ability to inhibit iron-dependent lipid peroxidation in liposomes compared to cholesterol, ergosterol and tamoxifen and relevance to anticancer action. FEBS Lett. 1993, 326, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Dawoud, H.; Malinski, T. Nanomedical studies of the restoration of nitric oxide/peroxynitrite balance in dysfunctional endothelium by 1,25-dihydroxy vitamin D3—Clinical implications for cardiovascular diseases. Int. J. Nanomed. 2018, 13, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, R.; Morimoto, I.; Morita, E.; Sugimoto, H.; Ito, Y.; Eto, S. Androgen receptors, 5 alpha-reductase activity and androgen-dependent proliferation of vascular smooth muscle cells. J. Steroid Biochem. Mol. Biol. 1994, 50, 169–174. [Google Scholar] [CrossRef]
- Dai-Do, D.; Espinosa, E.; Liu, G.; Rabelink, T.J.; Julmy, F.; Yang, Z.; Mahler, F.; Lüscher, T.F. 17 beta-estradiol inhibits proliferation and migration of human vascular smooth muscle cells: Similar effects in cells from postmenopausal females and in males. Cardiovasc. Res. 1996, 32, 980–985. [Google Scholar] [PubMed]
- Komukai, K.; Mochizuki, S.; Yoshimura, M. Gender and the renin-angiotensin-aldosterone system. Fundam. Clin. Pharmacol. 2010, 24, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Forman, J.P.; Williams, J.S.; Fisher, N.D. Plasma 25-Hydroxyvitamin D and Regulation of the Renin-Angiotensin System in Humans. Hypertension 2010, 55, 1283–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickenig, G.; Bäumer, A.T.; Grohè, C.; Kahlert, S.; Strehlow, K.; Rosenkranz, S.; Stäblein, A.; Beckers, F.; Smits, J.F.M.; Daemen, M.J.A.P.; et al. Estrogen Modulates AT1Receptor Gene Expression In Vitro and In Vivo. Circulation 1998, 97, 2197–2201. [Google Scholar] [CrossRef] [Green Version]
- Calkin, A.C.; Sudhir, K.; Honisett, S.; Williams, M.R.I.; Dawood, T.; Komesaroff, P.A. Rapid Potentiation of Endothelium-Dependent Vasodilation by Estradiol in Postmenopausal Women Is Mediated via Cyclooxygenase 2. J. Clin. Endocrinol. Metab. 2002, 87, 5072–5075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, M.; Deb, S.; Sebastian, S.; Okamura, K.; Bulun, S.E. Estrogen up-regulates cyclooxygenase-2 via estrogen receptor in human uterine microvascular endothelial cells. Fertil. Steril. 2004, 81, 1351–1356. [Google Scholar] [CrossRef]
- Vanhoutte, P.M.; Tang, E.H.C. Endothelium-dependent contractions: When a good guy turns bad! J. Physiol. 2008, 586, 5295–5304. [Google Scholar] [CrossRef]
- Parkington, H. Prostacyclin and endothelium-dependent hyperpolarization. Pharmacol. Res. 2004, 49, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Félétou, M.; Huang, Y.; Vanhoutte, P.M. Vasoconstrictor prostanoids. Pflügers Arch. Eur. J. Physiol. 2010, 459, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Rivero, J.; Cachofeiro, V.; Lahera, V.; Aras-López, R.; Márquez-Rodas, I.; Salaices, M.; Xavier, F.E.; Ferrer, M.; Balfagón, G. Participation of Prostacyclin in Endothelial Dysfunction Induced by Aldosterone in Normotensive and Hypertensive Rats. Hypertension 2005, 46, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norman, P.; Wysocki, S.; Lamawansa, M. The role of vitamin D3 in the aetiology of abdominal aortic aneurysms. Med. Hypotheses 1995, 45, 17–20. [Google Scholar] [CrossRef]
- Watson, K.E.; Abrolat, M.L.; Malone, L.L.; Hoeg, J.M.; Doherty, T.; Detrano, R.; Demer, L.L. Active serum vitamin D levels are inversely correlated with coronary calcification. Circulation 1997, 96, 1755–1760. [Google Scholar] [CrossRef]
- Barbarawi, M.; Kheiri, B.; Zayed, Y.; Barbarawi, O.; Dhillon, H.; Swaid, B.; Yelangi, A.; Sundus, S.; Bachuwa, G.; Alkotob, M.L.; et al. Vitamin D Supplementation and Cardiovascular Disease Risks in More Than 83 000 Individuals in 21 Randomized Clinical Trials. JAMA Cardiol. 2019, 4, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Karakas, M.; Thorand, B.; Zierer, A.; Huth, C.; Meisinger, C.; Roden, M.; Rottbauer, W.; Peters, A.; Koenig, W.; Herder, C. Low Levels of Serum 25-Hydroxyvitamin D Are Associated with Increased Risk of Myocardial Infarction, Especially in Women: Results from the MONICA/KORA Augsburg Case-Cohort Study. J. Clin. Endocrinol. Metab. 2013, 98, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somjen, D.; Kohen, F.; Amir-Zaltsman, Y.; Knoll, E.; Stern, N. Vitamin D analogs modulate the action of gonadal steroids in human vascular cells in vitro. Am. J. Hypertens. 2000, 13, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Somjen, D.; Katzburg, S.; Baz, M.; Stern, N.; Posner, G.H. Modulation of the response to estradiol-17β of rat vascular tissues by a non calcemic vitamin D analog. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 339–341. [Google Scholar] [CrossRef]
- Somjen, D.; Knoll, E.; Sharon, O.; Many, A.; Stern, N. Interaction between the effects of the selective estrogen modulator femarelle and a vitamin D analog in human umbilical artery vascular smooth muscle cells. J. Steroid Biochem. Mol. Biol. 2017, 174, 9–13. [Google Scholar] [CrossRef]
- Godoy, A.S.; Chung, I.; Montecinos, V.P.; Buttyan, R.; Johnson, C.S.; Smith, G.J. Role of androgen and vitamin D receptors in endothelial cells from benign and malignant human prostate. Am. J. Physiol. Metab. 2013, 304, E1131–E1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krohn, K.; Haffner, D.; Himmele, R.; Klaus, G.; Mehls, O.; Schaefer, F. 1, 25 (OH) 2 D 3 and Dihydrotestosterone Interact to Regulate Proliferation and Differentiation of Epiphyseal Chondrocytes. Calcif. Tissue Int. 2003, 73, 400–410. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sipos, M.; Péterffy, B.; Sziva, R.E.; Magyar, P.; Hadjadj, L.; Bányai, B.; Süli, A.; Soltész-Katona, E.; Gerszi, D.; Kiss, J.; et al. Vitamin D Deficiency Cause Gender Specific Alterations of Renal Arterial Function in a Rodent Model. Nutrients 2021, 13, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020704
Sipos M, Péterffy B, Sziva RE, Magyar P, Hadjadj L, Bányai B, Süli A, Soltész-Katona E, Gerszi D, Kiss J, et al. Vitamin D Deficiency Cause Gender Specific Alterations of Renal Arterial Function in a Rodent Model. Nutrients. 2021; 13(2):704. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020704
Chicago/Turabian StyleSipos, Miklós, Borbála Péterffy, Réka Eszter Sziva, Péter Magyar, Leila Hadjadj, Bálint Bányai, Anita Süli, Eszter Soltész-Katona, Dóra Gerszi, Judit Kiss, and et al. 2021. "Vitamin D Deficiency Cause Gender Specific Alterations of Renal Arterial Function in a Rodent Model" Nutrients 13, no. 2: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020704