
The Prebiotic Effect of Australian Seaweeds on Commensal Bacteria and Short Chain Fatty Acid Production in a Simulated Gut Model


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Seaweed Biomass
2.3. Proximate Analysis
Oxygen Radical Absorbance Capacity
2.4. Polysaccharide Extraction
2.5. Polyphenol Extraction
2.5.1. Phlorotannins from Brown Seaweeds
2.5.2. Polyphenols from Red and Green Seaweeds
2.6. Simulated Gastric Digestion
2.7. Simulated Anaerobic Digestion
2.7.1. Preparation of Basal Fermentation Medium
2.7.2. Fresh Faecal Inoculum
2.7.3. Anaerobic Fermentation
2.8. Short Chain Fatty Acid Quantification
2.9. Bacterial 16S rRNA Sequencing
2.10. Bioinformatics Methods
2.11. Statistical Analysis
3. Results
3.1. Compositional Content
3.2. Simulated Gastric Digestion
3.3. Short Chain Fatty Acid Production
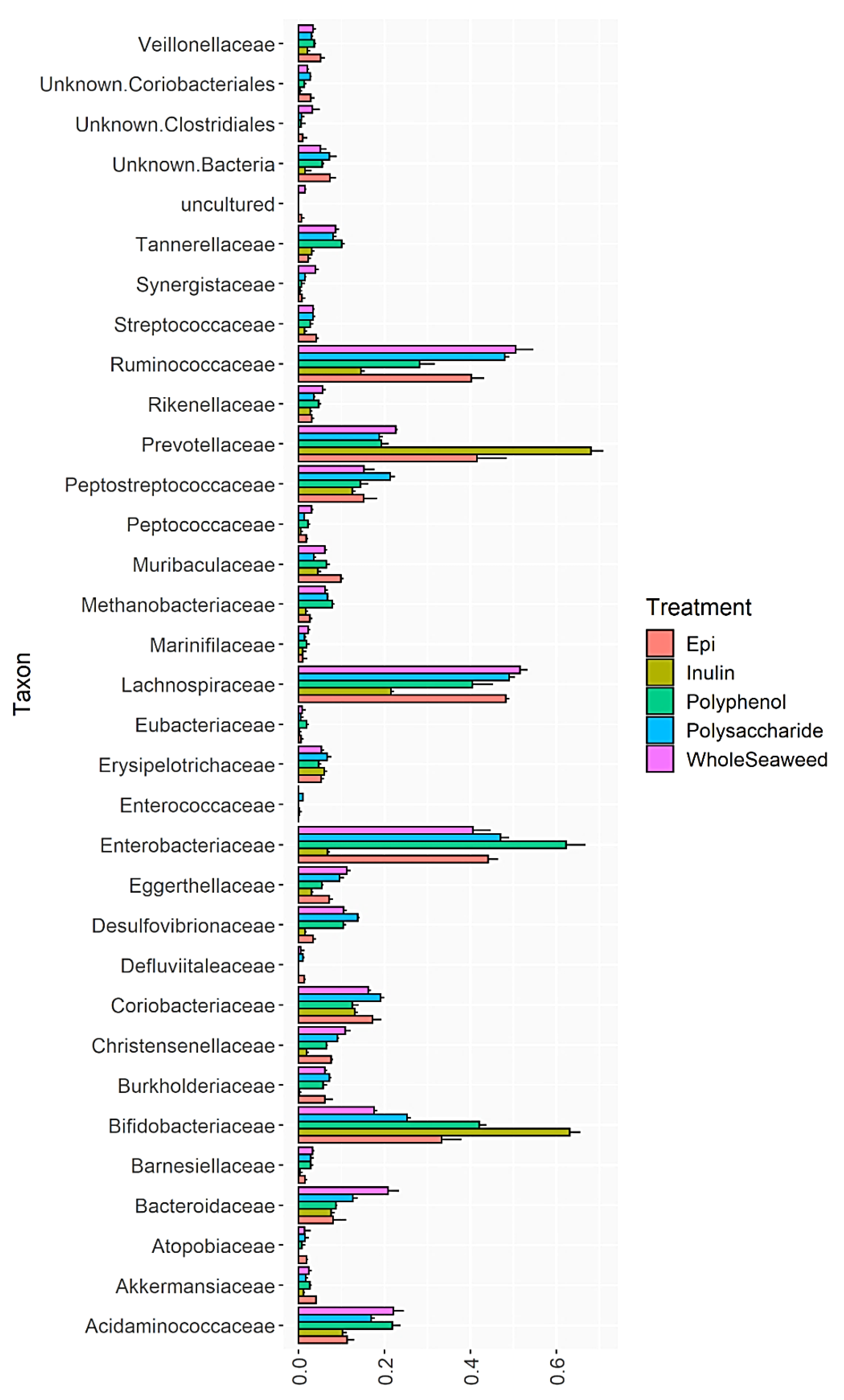
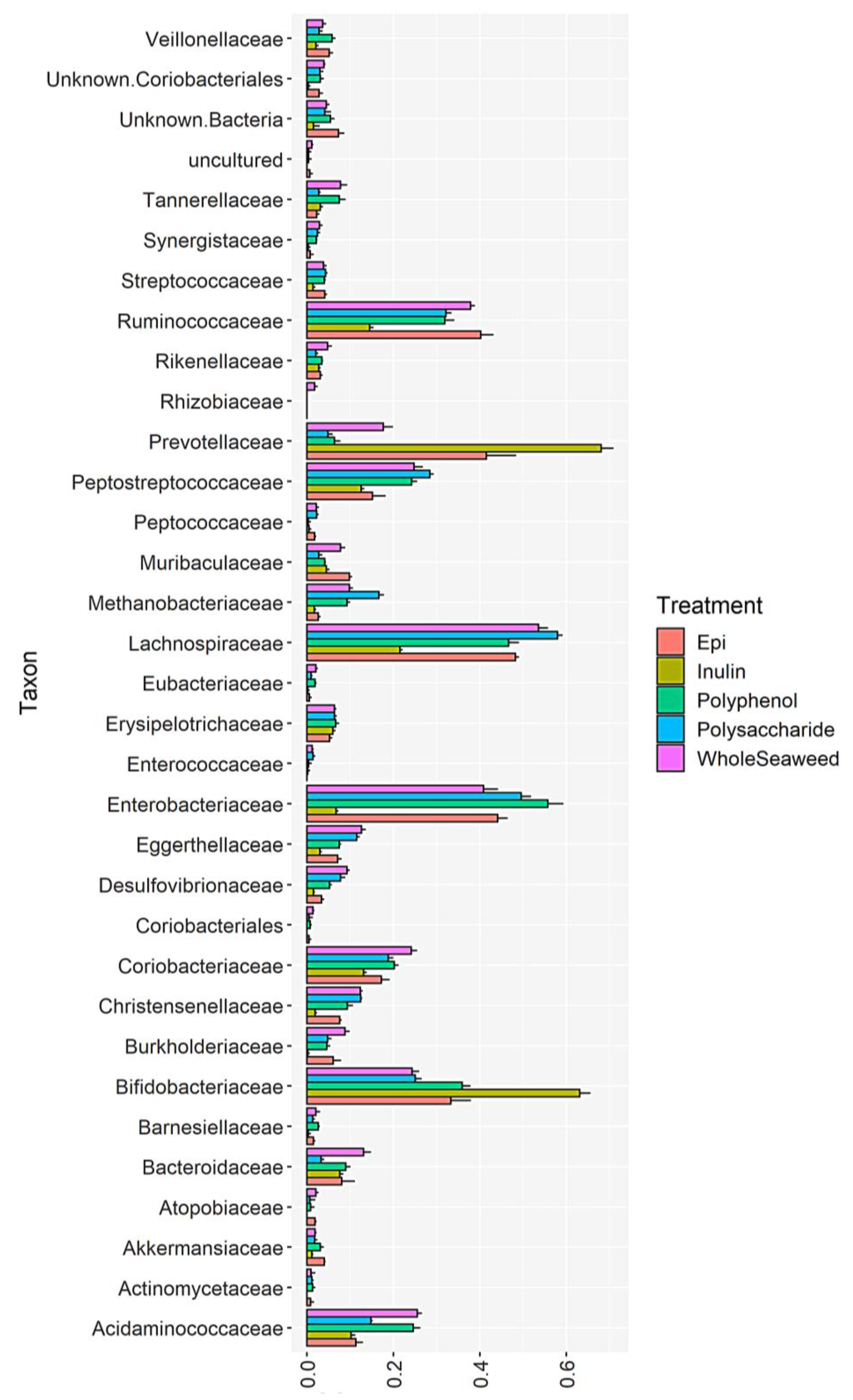
3.4. Bacterial Abundance

{kind=link}
{kind=link}
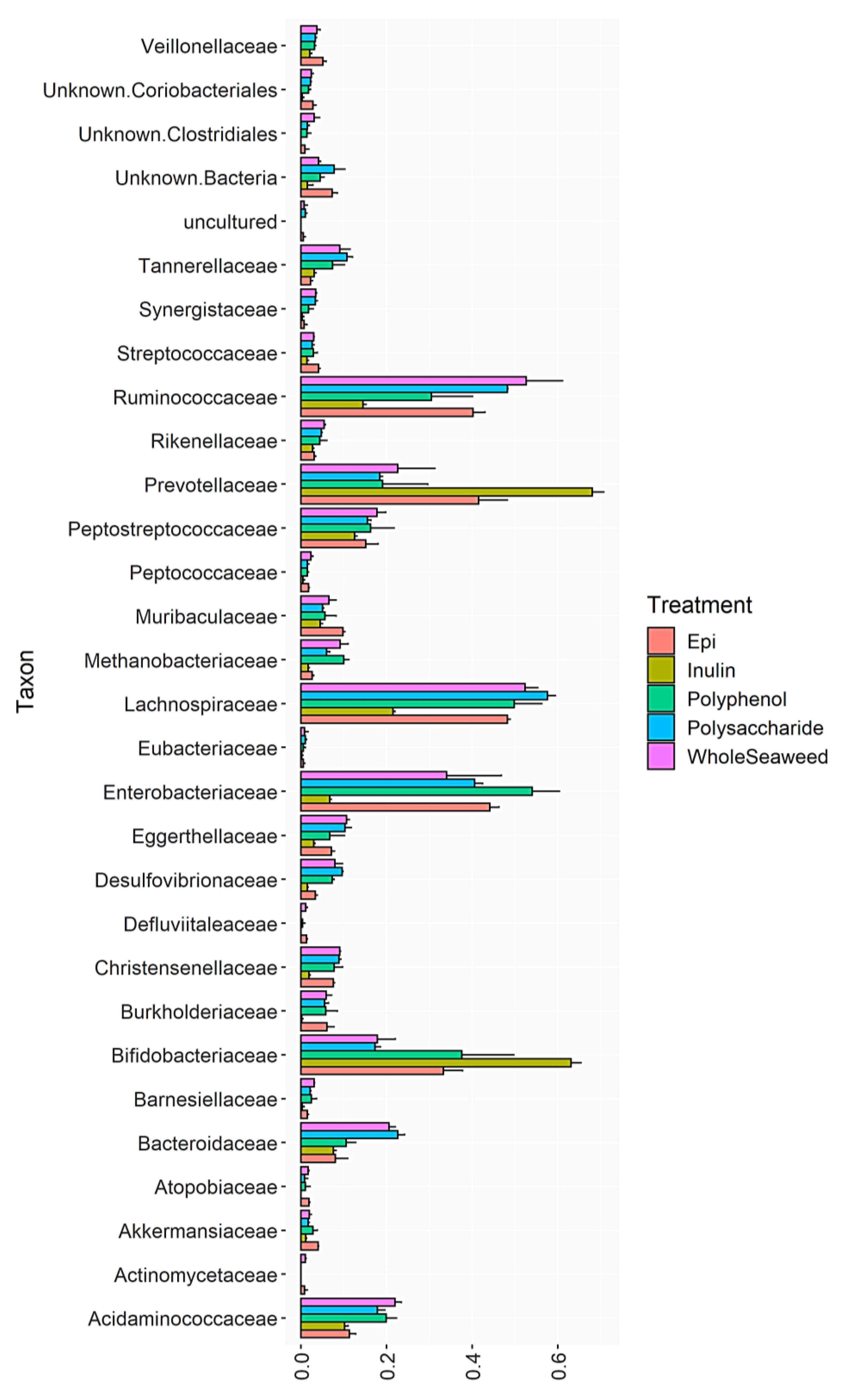{kind=link}
{kind=link}
| Phylum | Order | Genus | Species | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gram + | Gram - | Gram + | Gram - | / | Gram + | Gram + | Gram - | Gram + | Gram - | Gram + | Gram + | ||
| Substrate | Firmicutes | Bacteroidetes | Actinobacteria | Proteobacteria | Unclassified Bacterial Phylum | Lactobacillales | Faecalibacteria | Akkermansia | Roseburia | Barnesiella | Butyricicoccus | B. hydrogenotrophica | Shannon Diversity Index |
| Inulin | 32.50 | 68.71 | 64.48 | 6.91 | 1.47 | 1.44 | 10.63 | 1.15 | 3.08 | 0.26 | 3.04 | 8.67 × 10−17 | 3.38 |
| EGCG | 67.13 | 43.16 | 38.32 | 44.67 | 7.24 | 4.07 | 24.03 | 4.00 | 18.14 | 1.52 | 6.72 | 0.41 | 6.43 |
| P. comosa WH | ↑† 79.42 (p = 5.95 × 10−10) | ↓† 33.20 (p = 4.97 × 10−7) | ↓† 26.47 (p = 8.50 × 10−9) | ↑† 42.32 (p = 6.79 × 10−8) | ↑† 5.03 (p = 0.0132) | ↑† 3.36 (p = 9.7 × 10−4) | ↑† 20.54 (p = 2.4 × 10−6) | ↑† 2.34 NS | ↑† 12.27 (p = 2.08 × 10−7) | ↑† 3.23 (p = 0.0017) | ↑† 7.46 (p = 0.0004) | ↑† 3.32 (p = 0.00087) | ↑† 6.46 (p = 5.45 × 10−14) |
| P. comosa PS | ↑† 75.73 (p = 2.63 × 10−3) | ↓† 24.70 (p = 1.56 × 10−7) | ↓† 33.18 (p = 2.26 × 10−8) | ↑† 49.47 (p = 4.32 × 10−11) | ↑† 7.18 (p = 0.0030) | ↑† 3.50 (p = 0.0050) | ↑† 13.22 (p = 0.0126) | ↑† 1.70 NS | ↑† 7.06 (p = 6.21 × 10−5) | ↑† 2.76 (p = 0.0080) | ↑† 6.81 (p = 7.70 × 10−5) | ↓† 4.34 × 10−17 NS | ↑† 5.77 (p = 4.02 × 10−9) |
| P. comosa PP | ↓§ 57.35 (p = 0.0092) | ↓§ 24.87 (p = 0.0013) | ↑§ 44.19 NS | ↑§ 63.31 (p = 6.66 × 10−5) | ↓§ 5.53 NS | ↓§ 2.68 (p = 0.0195) | ↓§ 19.06 (p = 0.0470) | ↓§ 2.62 NS | ↓§ 5.51 (p = 2.80 × 10−5) | ↑§ 2.80 (p = 0.0340) | ↓§ 4.29 (p = 0.0126) | ↑§ 0.48 NS | ↓§ 4.79 (p = 4.26 × 10−6) |
| E. radiata WH | ↑† 81.55 (p = 6.97 × 10−8) | ↓† 33.41 (p = 5.94 × 10−5) | ↓† 25.92 (p = 1.57 × 10−6) | ↑† 35.67 (p = 0.0017) | ↑† 4.09 (p = 0.0137) | ↑† 3.03 (p = 0.0107) | ↑† 22.30 (p = 0.0019) | ↑† 1.97 NS | ↑† 11.17 (p = 0.0002) | ↑† 3.06 (p = 0.0007) | ↑† 7.42 (p = 0.0005) | ↑† 3.27 (p = 0.00092) | ↑† 6.25 (p = 2.84 × 10−5) |
| E. radiata PS | ↑† 80.44 (p = 0.0000) | ↓† 31.87 (p = 9.01 × 10−7) | ↓† 25.0 (p = 9.46 × 10−9) | ↑† 42.02 (p = 1.37 × 10−9) | ↑† 7.71 (p = 0.0035) | ↑† 2.57 (p = 0.0496) | ↑† 20.91 (p = 7.03 × 10−6) | ↑† 1.70 NS | ↑† 12.45 (p = 2.11 × 10−6) | ↑† 2.10 (p = 0.0111) | ↑† 8.39 (p = 5.44 × 10−5) | ↑† 3.34 (p = 7.83 × 10−5) | ↑† 6.46 (p = 7.69 × 10−14) |
| E. radiata PP | ↓§ 65.53 NS | ↓§ 24.53 (p = 0.0285) | ↑§ 41.31 NS | ↑§ 57.81 (p = 0.0279) | ↓§ 4.45 (p = 0.0359) | ↓§ 2.92 NS | ↓§ 15.13 (p = 0.0004) | ↓§ 2.82 NS | ↓§ 6.76 (p = 5.53 × 10−5) | ↑§ 2.39 (p = 0.0464) | ↓§ 5.95 NS | ↑§ 2.40 (p = 0.04372) | ↓§ 5.25 (p = 0.0008) |
| U. ohnoi WH | ↑† 77.73 (p = 0.00) | ↓† 25.16 (p = 1.5 × 10−6) | ↓† 36.78 (p = 9.31 × 10−8) | ↑† 42.79 (p = 1.06 × 10−8) | ↑† 4.51 (p = 5.26 × 103) | ↑† 4.01 (p = 0.0013) | ↑† 17.31 (p = 0.0003) | ↑† 1.82 NS | ↑† 9.57 (p = 1.08 × 10−5) | ↑† 2.01 (p = 0.0491) | ↑† 8.49 (p = 2.35 × 10−6) | ↑† 4.35 (p = 0.00013) | ↑† 6.38 (p = 3.56 × 10−13) |
| U. ohnoi PS | ↑† 77.28 (p = 0.00) | ↓† 0.740 (p = 4.14 × 10−8) | ↓† 33.54 (p = 3.20 × 10−8) | ↑† 50.38 (p = 2.30 × 10−10) | ↑† 4.12 (p = 0.0363) | ↑† 4.52 (p = 4.92 × 10−5) | ↑† 13.62 (p = 0.0313) | ↑† 1.77 NS | ↑† 8.80 (p = 3.82 × 10−5) | ↑† 1.31 (p = 0.0500) | ↑† 7.00 (p = 0.0003) | ↑† 3.32 (p = 7.02 × 10−5) | ↑† 5.78 (p = 2.89 × 10−9) |
| U. ohnoi PP | ↑§ 68.80 NS | ↓§ 14.63 (p = 6.47 × 10−5) | ↑§ 42.00 NS | ↑§ 56.07 (p = 0.0007) | ↓§ 5.41 NS | ↓§ 4.06 NS | ↓§ 21.89 NS | ↓§ 3.03 NS | ↓§ 6.81 (p = 7.40 × 10−8) | ↑§ 2.64 (p = 0.0061) | ↑§ 6.92 NS | ↓§ 2.32 × 10−17 (p = 3.26 × 10−6) | ↓§ 5.22 (p = 1.15 × 10−5) |



4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of Human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Vliet, M.J.; Tissing, W.J.E.; Dun, C.A.J.; Meessen, N.E.L.; Kamps, W.A.; de Bont, E.S.J.M.; Harmsen, H.J.M. Chemotherapy treatment in pediatric patients with acute myeloid leukemia receiving antimicrobial prophylaxis leads to a relative increase of colonization with potentially pathogenic bacteria in the gut. Clin. Infect. Dis. 2009, 49, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Portune, K.J.; Benítez-Páez, A.; Del Pulgar, E.M.G.; Cerrudo, V.; Sanz, Y. Gut microbiota, diet, and obesity-related disorders—The good, the bad, and the future challenges. Mol. Nutr. Food Res. 2017, 61, 1600252. [Google Scholar] [CrossRef] [Green Version]
- Sanz, Y.; De Palma, G. Gut microbiota and probiotics in modulation of epithelium and gut-associated lymphoid tissue function. Int. Rev. Immunol. 2009, 28, 397–413. [Google Scholar] [CrossRef] [Green Version]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Ellis, J.L.; Karl, J.P.; Oliverio, A.M.; Fu, X.; Soares, J.W.; Wolfe, B.E.; Hernandez, C.J.; Mason, J.B.; Booth, S.L. Dietary vitamin K is remodeled by gut microbiota and influences community composition. Gut Microbes 2021, 13, 1887721. [Google Scholar] [CrossRef]
- Stecher, B.; Hardt, W.-D. The role of microbiota in infectious disease. Trends Microbiol. 2008, 16, 107–114. [Google Scholar] [CrossRef]
- Khan, R.; Petersen, F.C.; Shekhar, S. Commensal bacteria: An emerging player in defense against respiratory pathogens. Front. Immunol. 2019, 10, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özgün, C.O.U.; Knut, R.; Dzung, B.D. Modulation of the gut microbiota by prebiotic fibres and bacteriocins. Microb. Ecol. Health Dis. 2017, 28, 1348886. [Google Scholar] [CrossRef] [Green Version]
- Hütt, P.; Shchepetova, J.; Lõivukene, K.; Kullisaar, T.; Mikelsaar, M. Antagonistic activity of probiotic Lactobacilli and Bifidobacteria against entero- and uropathogens. J. Appl. Microbiol. 2006, 100, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littman, D.R.; Pamer, E.G. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011, 10, 311–323. [Google Scholar] [CrossRef] [Green Version]
- James, K.R.; Gomes, T.; Elmentaite, R.; Kumar, N.; Gulliver, E.L.; King, H.W.; Stares, M.D.; Bareham, B.R.; Ferdinand, J.R.; Petrova, V.N.; et al. Distinct microbial and immune niches of the human colon. Nat. Immunol. 2020, 21, 343–353. [Google Scholar] [CrossRef]
- Jackson, M.A.; Verdi, S.; Maxan, M.-E.; Shin, C.M.; Zierer, J.; Bowyer, R.C.; Martin, T.; Williams, F.M.; Menni, C.; Bell, J.T. Gut microbiota associations with common diseases and prescription medications in a population-based cohort. Nat. Commun. 2018, 9, 2655. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Elson, C.O. Adaptive immune education by gut microbiota antigens. Immunology 2018, 154, 28–37. [Google Scholar] [CrossRef]
- Negi, S.; Das, D.K.; Pahari, S.; Nadeem, S.; Agrewala, J.N. Potential role of gut microbiota in induction and regulation of innate immune memory. Front. Immunol. 2019, 10, 2441. [Google Scholar] [CrossRef] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Čaja, F.; Stakheev, D.; Chernyavskiy, O.; Křížan, J.; Dvořák, J.; Rossmann, P.; Štěpánková, R.; Makovický, P.; Makovický, P.; Kozáková, H. Immune activation by microbiome shapes the colon mucosa: Comparison between healthy rat mucosa under conventional and germ-free conditions. J. Immunotoxicol. 2021, 18, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Amit-Romach, E.; Uni, Z.; Reifen, R. Therapeutic potential of two probiotics in inflammatory bowel disease as observed in the trinitrobenzene sulfonic acid model of colitis. Dis. Colon Rectum 2008, 51, 1828. [Google Scholar] [CrossRef] [PubMed]
- Lamousé-Smith, E.S.; Tzeng, A.; Starnbach, M.N. The intestinal flora is required to support antibody responses to systemic immunization in infant and germ free mice. PLoS ONE 2011, 6, e27662. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Chen, L.-L.; Lu, R.-Q.; Ma, W.-W.; Xiao, R. Alteration of intestinal microbiota composition in oral sensitized C3H/HeJ mice is associated with changes in dendritic cells and t cells in mesenteric lymph nodes. Front. Immunol. 2021, 12, 631494. [Google Scholar] [CrossRef]
- Ganal, S.C.; Sanos, S.L.; Kallfass, C.; Oberle, K.; Johner, C.; Kirschning, C.; Lienenklaus, S.; Weiss, S.; Staeheli, P.; Aichele, P. Priming of natural killer cells by nonmucosal mononuclear phagocytes requires instructive signals from commensal microbiota. Immunity 2012, 37, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.L.; Bittinger, K.; Lee, J.-J.; Friedman, E.S.; Mattei, L.M.; Graham, K.; Zhang, D.; Bush, J.; Balamuth, F.; McGowan, F.X., Jr.; et al. Intestinal microbiome diversity in pediatric sepsis: A conceptual framework for intestinal dysbiosis to influence immunometabolic function. Crit. Care Explor. 2021, 3, e0360. [Google Scholar] [CrossRef]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; classification, chemical properties, and future perspective applications in fields of pharmacology and biological medicine (A review of current applications and upcoming potentialities). J. Polym. Environ. 2021, 29, 2359–2371. [Google Scholar] [CrossRef]
- De Borba Gurpilhares, D.; Cinelli, L.P.; Simas, N.K.; Pessoa, A., Jr.; Sette, L.D. Marine prebiotics: Polysaccharides and oligosaccharides obtained by using microbial enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Sánchez-Muniz, F.J. Dietary fibre from edible seaweeds: Chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef]
- Lupton, J.R.; Betteridge, V.A.; Pijls, L.T.J. Codex final definition of dietary fibre: Issues of implementation. Qual. Assur. Saf. Crop. 2009, 1, 206–212. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed components as potential modulators of the gut microbiota. Mar. Drugs 2021, 19, 358. [Google Scholar] [CrossRef] [PubMed]
- Tannock, G.W.; Liu, Y. Guided dietary fibre intake as a means of directing short-chain fatty acid production by the gut microbiota. J. R. Soc. N. Z. 2020, 50, 434–455. [Google Scholar] [CrossRef]
- Roediger, W. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 1980, 21, 793–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritzhaupt, A.; Wood, I.S.; Ellis, A.; Hosie, K.B.; Shirazi-Beechey, S.P. Identification and characterization of a monocarboxylate transporter (MCT1) in pig and human colon: Its potential to transport l-lactate as well as butyrate. J. Physiol. 1998, 513, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Zambell, K.L.; Fitch, M.D.; Fleming, S.E. Acetate and butyrate are the major substrates for de novo lipogenesis in rat colonic epithelial cells. J. Nutr. 2003, 133, 3509–3515. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Tsujikawa, T.; Fujiyama, Y. Role of dietary fiber and short-chain fatty acids in the colon. Curr. Pharm. Des. 2003, 9, 347–358. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [Green Version]
- Docampo, M.D.; Stein-Thoeringer, C.K.; Lazrak, A.; Burgos da Silva, M.D.; Cross, J.; van den Brink, M.R.M. Expression of the butyrate/niacin receptor, GPR109a on T cells plays an important role in a mouse model of graft versus host disease. Blood 2018, 132, 61. [Google Scholar] [CrossRef]
- Ikeda-Ohtsubo, W.; López Nadal, A.; Zaccaria, E.; Iha, M.; Kitazawa, H.; Kleerebezem, M.; Brugman, S. Intestinal microbiota and immune modulation in Zebrafish by fucoidan from Okinawa mozuku (Cladosiphon okamuranus). Front. Nutr. 2020, 7, 67. [Google Scholar] [CrossRef]
- Hui, Y.; Tamez-Hidalgo, P.; Cieplak, T.; Satessa, G.D.; Kot, W.; Søren, S.K.; Nielsen, M.O.; Nielsen, D.S.; Krych, L. Supplementation of a lacto-fermented rapeseed-seaweed blend promotes gut microbial- and gut immune-modulation in weaner piglets. J. Anim. Sci. Biotechnol. 2020, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/AKT signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Gueven, N.; Spring, K.J.; Holmes, S.; Ahuja, K.; Eri, R.; Park, A.Y.; Fitton, J.H. Micro RNA expression after ingestion of fucoidan; a clinical study. Mar. Drugs 2020, 18, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terada, A.; Hara, H.; Mitsuoka, T. Effect of dietary alginate on the faecal microbiota and faecal metabolic activity in humans. Microb. Ecol. Health Dis. 1995, 8, 259–266. [Google Scholar] [CrossRef]
- Al Hinai, E.A.; Kullamethee, P.; Rowland, I.R.; Swann, J.; Walton, G.E.; Commane, D.M. Modelling the role of microbial p-cresol in colorectal genotoxicity. Gut Microbes 2019, 10, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Glorieux, G.; Gryp, T.; Perna, A. Gut-derived metabolites and their role in immune dysfunction in chronic kidney disease. Toxins 2020, 12, 245. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Kandasamy, S.; Zhang, J.; Kirby, C.W.; Karakach, T.; Hafting, J.; Critchley, A.T.; Evans, F.; Prithiviraj, B. Prebiotic effects of diet supplemented with the cultivated red seaweed Chondrus crispus or with fructo-oligo-saccharide on host immunity, colonic microbiota and gut microbial metabolites. BMC Complement. Altern. Med. 2015, 15, 279. [Google Scholar] [CrossRef]
- Du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the red seaweed Sarconema filiforme attenuate symptoms of diet-induced metabolic syndrome in rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Zhou, R.; Wang, Y.; Zhang, M.; Liu, K.; Ma, C. Beneficial effects of sulfated polysaccharides from the red seaweed Gelidium pacificum Okamura on mice with antibiotic-associated diarrhea. Food Funct. 2020, 11, 4625–4637. [Google Scholar] [CrossRef]
- Takahashi, M.; Takahashi, K.; Abe, S.; Yamada, K.; Suzuki, M.; Masahisa, M.; Endo, M.; Abe, K.; Inoue, R.; Hoshi, H. Improvement of psoriasis by alteration of the gut environment by oral administration of fucoidan from Cladosiphon okamuranus. Mar. Drugs 2020, 18, 154. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.; Sweeney, T.; Bahar, B.; O’Doherty, J. Effect of maternal seaweed extract supplementation on suckling piglet growth, humoral immunity, selected microflora, and immune response after an ex vivo lipopolysaccharide challenge. J. Anim. Sci. 2012, 90, 505–514. [Google Scholar] [CrossRef]
- Wang, Y.; Han, F.; Hu, B.; Li, J.; Yu, W. In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nutr. Res. 2006, 26, 597–603. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–598. [Google Scholar] [CrossRef]
- Poole, J.; Diop, A.; Rainville, L.C.; Barnabé, S. Bioextracting polyphenols from the brown seaweed Ascophyllum nodosum from Québec’s north shore coastline. Ind. Biotechnol. 2019, 15, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Wekre, M.E.; Kåsin, K.; Underhaug, J.; Holmelid, B.; Jordheim, M. Quantification of polyphenols in seaweeds: A case study of Ulva Intestinalis. Antioxid. 2019, 8, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.J.; Liu, R.; Xue, B.; Luo, J.; Gao, L.; Wang, Y.; Ou, S.; Li, S.; Peng, X. Impact and consequences of polyphenols and fructooligosaccharide interplay on gut microbiota in rats. Food Funct. 2017, 8, 1925–1932. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Conlon, M.A.; Vuaran, M.S.; Franco, C.M.M.; Zhang, W. Polysaccharide and phlorotannin-enriched extracts of the brown seaweed Ecklonia radiata influence human gut microbiota and fermentation in vitro. J. Appl. Phycol. 2017, 29, 2407–2416. [Google Scholar] [CrossRef]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal relationship between diet-induced gut microbiota changes and diabetes: A novel strategy to transplant Faecalibacterium prausnitzii in preventing diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-rich extracts from brown macroalgae Lessonia trabeculata attenuate hyperglycemia and modulate gut microbiota in high-fat diet and streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef]
- Lin, G.; Liu, X.; Yan, X.; Liu, D.; Yang, C.; Liu, B.; Huang, Y.; Zhao, C. Role of green macroalgae Enteromorpha prolifera polyphenols in the modulation of gene expression and intestinal microflora profiles in type 2 diabetic mice. Int. J. Mol. Sci. 2019, 20, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecerf, J.M.; Dépeint, F.; Clerc, E.; Dugenet, Y.; Niamba, C.N.; Rhazi, L.; Cayzeele, A.; Abdelnour, G.; Jaruga, A.; Younes, H.; et al. Xylo-oligosaccharide (XOS) in combination with inulin modulates both the intestinal environment and immune status in healthy subjects, while XOS alone only shows prebiotic properties. Br. J. Nutr. 2012, 108, 1847–1858. [Google Scholar] [CrossRef]
- Reimer, R.A.; Soto-Vaca, A.; Nicolucci, A.C.; Mayengbam, S.; Park, H.; Madsen, K.L.; Menon, R.; Vaughan, E.E. Effect of chicory inulin-type fructan–containing snack bars on the human gut microbiota in low dietary fiber consumers in a randomized crossover trial. Am. J. Clin. Nutr. 2020, 111, 1286–1296. [Google Scholar] [CrossRef]
- Holscher, H.D.; Bauer, L.L.; Gourineni, V.; Pelkman, C.L.; Fahey, G.C., Jr.; Swanson, K.S. Agave inulin supplementation affects the fecal microbiota of healthy adults participating in a randomized, double-blind, placebo-controlled, crossover trial. J. Nutr. 2015, 145, 2025–2032. [Google Scholar] [CrossRef] [Green Version]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2010, 93, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Yamakoshi, J.; Tokutake, S.; Kikuchi, M.; Kubota, Y.; Konishi, H.; Mitsuoka, T. Effect of proanthocyanidin-rich extract from grape seeds on human fecal flora and fecal odor. Microb. Ecol. Health Dis. 2001, 13, 25–31. [Google Scholar]
- ISO 6496:1999; Animal Feeding Stuffs—Determination of Moisture and Other Volatile Matter Content. International Organization for Standardization: Geneva, Switzerland, 1999. Available online: https://www.iso.org/standard/12871.html (accessed on 11 February 2022).
- ISO 2171:2007; Cereals, Pulses and By-Products—Determination of Ash Yield by Incineration. International Organization for Standardization: Geneva, Switzerland, 2007. Available online: https://www.iso.org/standard/37264.html. (accessed on 6 February 2022).
- AOAC Method 986.15-1988; Arsenic, Cadmium, Lead, Selenium and Zinc in Human and Pet Foods—Multielement Method. Association of Official Analytical Chemists: Rockville, MD, USA, 1996. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&products_id=203. (accessed on 2 January 2022).
- AOAC Method 974.14. 974.14-1976; Mercury in Fish—Alternative Digestion Method. Association of Official Analytical Chemists: Rockville, MD, USA, 1996. Available online: http://www.aoacofficialmethod.org/index.php?main_page=products_all&disp_order=3&page=127. (accessed on 8 February 2022).
- USEPA Method 6010D (SW-846); Inductively Coupled Plasma-Atomic Emission Spectrometry—Revision 4. United States Environmental Protection Agency: Washington, DC, USA, 2014. Available online: https://www.epa.gov/esam/epa-method-6010d-sw-846-inductively-coupled-plasma-atomic-emission-spectrometry. (accessed on 2 January 2022).
- USEPA Method 6020B (SW-846); Inductively Coupled Plasma-Mass Spectrometry—Revision 2. United States Environmental Protection Agency: Washington, DC, USA, 2014. Available online: https://www.epa.gov/esam/epa-method-6020b-sw-846-inductively-coupled-plasma-mass-spectrometry. (accessed on 2 January 2022).
- AOAC Method 992.23-1992(1998); Crude Protein in Cereal Grains and Oilseeds—Generic Combustion Method. Association of Official Analytical Chemists: Rockville, MD, USA, 1998. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&products_id=1635 (accessed on 14 February 2022).
- Biancarosa, I.; Espe, M.; Bruckner, C.G.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E.J. Amino acid composition, protein content, and nitrogen-to-protein conversion factors of 21 seaweed species from Norwegian waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- AOAC Method 991.43-1994; Total Dietary Fiber in Foods—Enzymatic-Gravimetric Method MES-TRIS Buffer. Association of Official Analytical Chemists: Rockville, MD, USA, 1994. Available online: http://www.eoma.aoac.org/methods/info.asp?ID=26991 (accessed on 19 January 2022).
- AOAC Method 991.1313-1999; Total Dietary Fiber (Determined as Neutral Sugar Residues, Uronic Acid Residues, And Klason Lignin)—Gas Chromatographic-Colorimetric-Gravimetric Method. Association of Official Analytical Chemists: Rockville, MD, USA, 1999. Available online: http://www.eoma.aoac.org/methods/info.asp?ID=26991 (accessed on 14 February 2022).
- AOAC Method 2017.13-2017; Total Phenolic Content in Extracts—Analyte, Phenolic Content—Analytical Technique, Folin-C (Folin and Ciocalteu) Colorimetric Method. Association of Official Analytical Chemists: Rockville, MD, USA, 2018. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&cPath=1&products_id=300 (accessed on 19 January 2022).
- Ou, B.; Chang, T.; Huang, D.; Prior, R.L. Determination of total antioxidant capacity by oxygen radical absorbance capacity (ORAC) using fluorescein as the fluorescence probe: AOAC first action 2012.23. J. AOAC Int. 2013, 96, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Dore, C.M.P.G.; das C. Faustino Alves, M.G.; Will, L.S.E.P.; Costa, T.G.; Sabry, D.A.; de Souza Rêgo, L.A.R.; Accardo, C.M.; Rocha, H.A.O.; Filgueira, L.G.A.; Leite, E.L. A sulfated polysaccharide, fucans, isolated from brown algae Sargassum vulgare with anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Sousa, C.; Silva Lí, R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.H.S.; Jacobsen, C. Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem. 2013, 138, 1670. [Google Scholar] [CrossRef]
- Bird, A.R.; Usher, S.; May, B.; Topping, D.L.; Morell, M.K. Resistant Starch: Measurement, Intakes, and Dietary Targets. In Dietary Fiber and Health; Cho, S., Almeida, N., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Chapter 5; pp. 60–75. [Google Scholar] [CrossRef]
- Zhou, Z.; Cao, X.; Zhou, J.Y.H. Effect of resistant starch structure on short-chain fatty acids production by human gut microbiota fermentation in vitro. Starch Stärke 2013, 65, 509–516. [Google Scholar] [CrossRef]
- Watson, E.-J.; Giles, J.; Scherer, B.L.; Blatchford, P. Human faecal collection methods demonstrate a bias in microbiome composition by cell wall structure. Sci. Rep. 2019, 9, 16831. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Dillon, M.R.; Zhang, Y.; Rideout, J.R.; Bolyen, E.; Li, H.; Albert, P.S.; Caporaso, J.G. q2-longitudinal: Longitudinal and paired-sample analyses of microbiome data. mSystems 2018, 3, e00219-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Allen-Vercoe, E.; Petrof, E.O. Fecal microbiota transplantation: In perspective. Ther. Adv. Gastroenterol. 2016, 9, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaila, A.A. Investigating the Anti-Methanogenic Properties of Select Species of Seaweed in New Zealand. Master’s Thesis, The University of Waikato, Hamilton, New Zealand, 2020. Available online: https://hdl.handle.net/10289/13946 (accessed on 22 December 2021).
- Charoensiddhi, S. Process Development for Functional Food Ingredients with Gut Health Benefits from The Brown Seaweed Ecklonia radiata. Ph.D. Thesis, College of Medicine and Public Health, Flinders University, Adelaide, South Australia, 2017. Available online: https://flex.flinders.edu.au/file/75b7665e-6946-419f-bf28-1cdd6cc45da4/1/Thesis_Suvimol_Final%20approval.pdf. (accessed on 10 January 2022).
- EC. European Commission HaCPD-GSCoF, Opinion of the Scientific Committee on Food on the Tolerable Upper Level of Intake of Iodine; European Commission: Brussels, Belgium, 2002; Available online: https://apps.who.int/iris/bitstream/handle/10665/338064/WHO-HEP-ECH-WSH-2020.5-eng.pdf. (accessed on 1 January 2022).
- Magnusson, M.; Glasson, C.R.; Vucko, M.J.; Angell, A.; Neoh, T.L.; de Nys, R. Enrichment processes for the production of high-protein feed from the green seaweed Ulva Ohnoi. Algal Res. 2019, 41, 101555. [Google Scholar] [CrossRef]
- García-Casal, M.N.; Ramirez, J.; Leets, I.; Pereira, A.C.; Quiroga, M.F. Antioxidant capacity, polyphenol content and iron bioavailability from algae (Ulva sp., Sargassum sp. and Porphyra sp.) in human subjects. Brit. J. Nutr. 2008, 101, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, P.D. Biogeographical variation in brown algal polyphenolics and other secondary metabolites: Comparison between temperate Australasia and North America. Oecologia 1989, 78, 373–382. [Google Scholar] [CrossRef]
- Skrzypczyk, V.M.; Hermon, K.M.; Norambuena, F.; Turchini, G.M.; Keast, R.; Bellgrove, A. Is Australian seaweed worth eating? Nutritional and sensorial properties of wild-harvested Australian versus commercially available seaweeds. J. Appl. Phycol. 2019, 31, 709–724. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Lee, O.H.; Yoon, K.Y.; Kim, K.J.; You, S.; Lee, B.Y. Seaweed extracts as a potential tool for the attenuation of oxidative damage in obesity-related pathologies. J. Phycol. 2011, 47, 548–556. [Google Scholar] [CrossRef]
- Fernandez-Lazaro, C.I.; Martínez-González, M.Á.; Aguilera-Buenosvinos, I.; Gea, A.; Ruiz-Canela, M.; Romanos-Nanclares, A.; Toledo, E. Dietary antioxidant vitamins and minerals and breast cancer risk: Prospective results from the SUN cohort. Antioxidants 2021, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food. Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Gorinstein, S.; Böhm, V.; Schaich, K.M.; Özyürek, M.; Güçlü, K. Methods of measurement and evaluation of natural antioxidant capacity/activity (IUPAC Technical Report). Pure Appl. Chem. 2013, 85, 957–998. [Google Scholar] [CrossRef] [Green Version]
- Charoensiddhi, S.; Franco, C.; Su, P.; Zhang, W. Improved antioxidant activities of brown seaweed Ecklonia radiata extracts prepared by microwave-assisted enzymatic extraction. J. Appl. Phycol. 2015, 27, 2049–2058. [Google Scholar] [CrossRef]
- Sáez, M.I.; Suárez, M.D.; Alarcón, F.J.; Martínez, T.F. Assessing the potential of algae extracts for extending the shelf life of rainbow trout (Oncorhynchus mykiss) fillets. Foods 2021, 10, 910. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Islam, G. Seaweed antioxidants as novel ingredients for better health and food quality: Bangladesh prospective. Proc. Pak. Acad. Sci. 2014, 51, 215–233. Available online: https://paspk.org/wp-content/uploads/proceedings/51,%20No.3/a7383af9Seaweed.pdf. (accessed on 19 December 2021).
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Ar Gall, E. Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Van Hees, D.H.; Olsen, Y.S.; Wernberg, T.; Van Alstyne, K.L.; Kendrick, G.A. Phenolic concentrations of brown seaweeds and relationships to nearshore environmental gradients in Western Australia. Mar. Biol. 2017, 164, 74. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, M.; Yuen, A.K.L.; Zhang, R.; Wright, J.T.; Taylor, R.B.; Maschmeyer, T.; de Nys, R. A comparative assessment of microwave assisted (MAE) and conventional solid-liquid (SLE) techniques for the extraction of phloroglucinol from brown seaweed. Algal Res. 2017, 23, 28–36. [Google Scholar] [CrossRef]
- Ivanova, V.; Rouseva, R.; Kolarova, M.; Serkedjieva, J.; Rachev, R.; Manolova, N. Isolation of a polysaccharide with antiviral effect from Ulva lactuca. Prep. Biochem. 1994, 24, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Charoensiddhi, S.; Lorbeer, A.J.; Lahnstein, J.; Bulone, V.; Franco, C.M.M.; Zhang, W. Enzyme-assisted extraction of carbohydrates from the brown alga Ecklonia radiata: Effect of enzyme type, pH and buffer on sugar yield and molecular weight profiles. Process. Biochem. 2016, 51, 1503–1510. [Google Scholar] [CrossRef]
- Lorbeer, A. Advanced and Integrated Technologies for the Valorization of South Australia’s Unique Seaweed Resources. Ph.D. Thesis, Department of Medical Biotechnology, Flinders University, Adelaide, South Australia, 2016. Available online: https://flex.flinders.edu.au/file/16f1e83d-71c0-427d-bcb8-ee40b0408e95/1/ThesisLorbeer2016.pdf (accessed on 4 January 2022).
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.; Jeon, K.; Moon, S.; Lee, K.; Kim, W.-K.; Jeong, H.; Cha, K.H.; Lim, M.Y.; Kang, W.; Kweon, M.-N.; et al. Roseburia spp. Abundance Associates with Alcohol Consumption in Humans and Its Administration Ameliorates Alcoholic Fatty Liver in Mice. Cell Host Microbe 2020, 27, 25–40.e6. [Google Scholar] [CrossRef]
- Boesmans, L.; Valles-Colomer, M.; Wang, J.; Eeckhaut, V.; Falony, G.; Ducatelle, R.; Immerseel, F.V.; Raes, J.; Verbeke, K.; Cotter, P.D. Butyrate producers as potential next-generation probiotics: Safety assessment of the administration of Butyricicoccus pullicaecorum to healthy volunteers. mSystems 2018, 3, e00094-e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. Available online: https://www.jpp.krakow.pl/journal/archive/10_12/pdf/497_10_12_article.pdf. (accessed on 11 April 2022).
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Dell’Anno, M.; Sotira, S.; Rebucci, R.; Reggi, S.; Castiglioni, B.; Rossi, L. In vitro evaluation of antimicrobial and antioxidant activities of algal extracts. Ital. J. Anim. Sci. 2020, 19, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Z.; Bach, S.; McAllister, T. Sensitivity of Escherichia coli to seaweed (Ascophyllum nodosum) phlorotannins and terrestrial tannins. Asian-Australas. J. Anim. Sci. 2009, 22, 238–245. [Google Scholar] [CrossRef]
- Buitimea-Cantúa, N.E.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Phenolic–protein interactions: Effects on food properties and health benefits. J. Med. Food 2018, 21, 188–198. [Google Scholar] [CrossRef]
- Tang, J.; Wang, W.; Chu, W. Antimicrobial and anti-quorum sensing activities of phlorotannins from seaweed (Hizikia fusiforme). Front. Cell. Infect. Microbiol. 2020, 10, 586750. [Google Scholar] [CrossRef] [PubMed]
- Ford, L.; Stratakos, A.C.; Theodoridou, K.; Dick, J.T.A.; Sheldrake, G.N.; Linton, M.; Corcionivoschi, N.; Walsh, P.J. Polyphenols from brown seaweeds as a potential antimicrobial agent in animal feeds. ACS Omega 2020, 5, 9093–9103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, T.P.N.; Troise, A.D.; Nijsse, B.; Roviello, G.N.; Fogliano, V.; de Vos, W.M. Intestinimonas-like bacteria are important butyrate producers that utilize Nε-fructosyllysine and lysine in formula-fed infants and adults. J. Funct. Foods 2020, 70, 103974. [Google Scholar] [CrossRef]
- Mehta, B.M.; Deeth, H.C. Blocked lysine in dairy products: Formation, occurrence, analysis, and nutritional implications. Compr. Rev. Food Sci. Food Saf. 2016, 15, 206–218. [Google Scholar] [CrossRef]
- Ubeda, C.; Bucci, V.; Caballero, S.; Djukovic, A.; Toussaint, N.C.; Equinda, M.; Lipuma, L.; Ling, L.; Gobourne, A.; No, D.; et al. Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect. Immun. 2013, 81, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Daillère, R.; Vétizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.M.; Flament, C.; Lepage, P.; Roberti, M.P.; et al. Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, A.; Aoe, S.; Watanabe, S.; Hisada, T.; Mochizuki, J.; Mizuno, S.; Hoshi, T.; Kodama, S. The nested study on the intestinal microbiota in GENKI study with special reference to the effect of brown rice eating. J. Obes. Chronic Dis. 2019, 3, 1–13. [Google Scholar] [CrossRef]
- Tan, P.L.; Kim, S.H. Probiotics: Emerging functional ingredients for healthy aging and age-related diseases. In Probiotic Beverages; Panda, S.K., Russell, I., Kellershohn, J., Eds.; Academic Press: London, UK, 2021; Chapter 10; pp. 175–212. [Google Scholar] [CrossRef]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef]
- Kriaa, A.; Bourgin, M.; Potiron, A.; Mkaouar, H.; Jablaoui, A.; Gérard, P.; Maguin, E.; Rhimi, M. Microbial impact on cholesterol and bile acid metabolism: Current status and future prospects. J. Lipid Res. 2019, 60, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia—A new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20–76 years of age. Npj Biofilms Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Geerlings, S.Y.; Kostopoulos, I.; de Vos, W.M.; Belzer, C. Akkermansia muciniphila in the human gastrointestinal tract: When, where, and how? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila adheres to enterocytes and strengthens the integrity of the epithelial cell layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Derrien, M.; Isolauri, E.; de Vos, W.M.; Salminen, S. Intestinal integrity and Akkermansia muciniphila, a mucin-degrading member of the intestinal microbiota present in infants, adults, and the elderly. Appl. Environ. Microbiol. 2007, 73, 7767–7770. [Google Scholar] [CrossRef] [Green Version]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Demirci, M.; Tokman, H.B.; Taner, Z.; Keskin, F.E.; Çağatay, P.; Bakar, Y.O.; Özyazar, M.; Kiraz, N.; Kocazeybek, B.S. Bacteroidetes and Firmicutes levels in gut microbiota and effects of hosts TLR2/TLR4 gene expression levels in adult type 1 diabetes patients in Istanbul, Turkey. J. Diabetes Complicat. 2020, 34, 107449. [Google Scholar] [CrossRef]
- Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366–380. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Dima, S.; Jafari, S.M. Bioavailability and bioaccessibility of food bioactive compounds; overview and assessment by in vitro methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2862–2884. [Google Scholar] [CrossRef] [PubMed]
- Déjean, G.; Tamura, K.; Cabrera, A.; Jain, N.; Pudlo, N.A.; Pereira, G.; Viborg, A.H.; Van Petegem, F.; Martens, E.C.; Brumer, H. Synergy between cell surface glycosidases and glycan-binding proteins dictates the utilization of specific beta(1,3)-glucans by human gut Bacteroides. mBio 2020, 11, e00095-e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, S.; Touvrey-Loiodice, M.; Poulet, L.; Drouillard, S.; Vincentelli, R.; Henrissat, B.; Skjåk-Bræk, G.; Helbert, W. Ancient acquisition of “alginate utilization loci” by human gut microbiota. Sci. Rep. 2018, 8, 8075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, F.; Barbeyron, T.; Tonon, T.; Génicot, S.; Czjzek, M.; Michel, G. Characterization of the first alginolytic operons in a marine bacterium: From their emergence in marine Flavobacteriia to their independent transfers to marine Proteobacteria and human gut Bacteroides. Environ. Microbiol. 2012, 14, 2379–2394. [Google Scholar] [CrossRef]
- Kabisch, A.; Otto, A.; König, S.; Becher, D.; Albrecht, D.; Schüler, M.; Teeling, H.; Amann, R.I.; Schweder, T. Functional characterization of polysaccharide utilization loci in the marine Bacteroidetes ‘Gramella forsetii’ KT0803. ISME J. 2014, 8, 1492–1502. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Xu, H.; Wei, C.; Jiang, T.; Qin, S.; Zhang, W.; Cao, Y.; Hu, C.; Zhang, F.; Qiao, D.; et al. Horizontal transfer of a novel soil agarase gene from marine bacteria to soil bacteria via human microbiota. Sci. Rep. 2016, 6, 34103. [Google Scholar] [CrossRef]
- Hehemann, J.-H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef]
- Pudlo, N.A.; Pereira, G.V.; Parnami, J.; Cid, M.; Markert, S.; Tingley, J.P.; Unfried, F.; Ali, A.; Campbell, A.; Urs, K.; et al. Extensive transfer of genes for edible seaweed digestion from marine to human gut bacteria. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pluvinage, B.; Grondin, J.M.; Amundsen, C.; Klassen, L.; Moote, P.E.; Xiao, Y.; Thomas, D.; Pudlo, N.A.; Anele, A.; Martens, E.C.; et al. Molecular basis of an agarose metabolic pathway acquired by a human intestinal symbiont. Nat. Commun. 2018, 9, 1043. [Google Scholar] [CrossRef]
- Kearney, S.M.; Gibbons, S.M.; Erdman, S.E.; Alm, E.J. Orthogonal dietary niche enables reversible engraftment of a gut bacterial commensal. Cell Rep. 2018, 24, 1842–1851. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, E.S.; DeLoache, W.C.; Pruss, K.M.; Whitaker, W.R.; Sonnenburg, J.L. An exclusive metabolic niche enables strain engraftment in the gut microbiota. Nature 2018, 557, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A common factor in human diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Ceccarani, C.; Bassanini, G.; Montanari, C.; Casiraghi, M.C.; Ottaviano, E.; Morace, G.; Biasucci, G.; Paci, S.; Borghi, E.; Verduci, E. Proteobacteria overgrowth and butyrate-producing taxa depletion in the gut microbiota of glycogen storage disease type 1 patients. Metabolites 2020, 10, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Reddy, C.R. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Salaün, S.; Kervarec, N.; Potin, P.; Haras, D.; Piotto, M.; La Barre, S. Whole-cell spectroscopy is a convenient tool to assist molecular identification of cultivatable marine bacteria and to investigate their adaptive metabolism. Talanta 2010, 80, 1758–1770. [Google Scholar] [CrossRef]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Murphy, E.F.; Cotter, P.D.; Hogan, A.; Sullivan, O.; Joyce, A.; Fouhy, F.; Clarke, S.F.; Marques, T.M.; Toole, P.W.; Stanton, C.; et al. Divergent metabolic outcomes arising from targeted manipulation of the gut microbiota in diet-induced obesity. Gut 2013, 62, 220. [Google Scholar] [CrossRef]
- Li, J.; Si, H.; Du, H.; Guo, H.; Dai, H.; Xu, S.; Wan, J. Comparison of gut microbiota structure and Actinobacteria abundances in healthy young adults and elderly subjects: A pilot study. BMC Microbiol. 2021, 21, 13. [Google Scholar] [CrossRef]
- Vázquez-Rodríguez, B.; Santos-Zea, L.; Heredia-Olea, E.; Acevedo-Pacheco, L.; Santacruz, A.; Gutiérrez-Uribe, J.A.; Cruz-Suárez, L.E. Effects of phlorotannin and polysaccharide fractions of brown seaweed Silvetia compressa on human gut microbiota composition using an in vitro colonic model. J. Funct. Foods 2021, 84, 104596. [Google Scholar] [CrossRef]
- Igwe, E.O.; Charlton, K.E.; Probst, Y.C.; Kent, K.; Netzel, M.E. A systematic literature review of the effect of anthocyanins on gut microbiota populations. J. Hum. Nutr. Diet. 2019, 32, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaci, N.; Borrel, G.; Tottey, W.; O’Toole, P.W.; Brugère, J.-F. Archaea and the human gut: New beginning of an old story. World J. Gastroenterol. 2014, 20, 16062–16078. [Google Scholar] [CrossRef] [PubMed]
- Nkamga, V.D.; Henrissat, B.; Drancourt, M. Archaea: Essential inhabitants of the human digestive microbiota. Hum. Microbiome J. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Camara, A.; Konate, S.; Tidjani Alou, M.; Kodio, A.; Togo, A.H.; Cortaredona, S.; Henrissat, B.; Thera, M.A.; Doumbo, O.K.; Raoult, D.; et al. Clinical evidence of the role of Methanobrevibacter smithii in severe acute malnutrition. Sci. Rep. 2021, 11, 5426. [Google Scholar] [CrossRef]
- Abbott, D.W.; Aasen, I.M.; Beauchemin, K.A.; Grondahl, F.; Gruninger, R.; Hayes, M.; Huws, S.; Kenny, D.A.; Krizsan, S.J.; Kirwan, S.F.; et al. Seaweed and seaweed bioactives for mitigation of enteric methane: Challenges and opportunities. Animals 2020, 10, 2432. [Google Scholar] [CrossRef]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
| (% of DW) | ||||||
|---|---|---|---|---|---|---|
| P. comosa WH | E. radiata WH | U. ohnoi WH | P. comosa PS | E. radiata PS | U. ohnoi PS | |
| Moisture | 6.04 ± 0.77 | 5.59 ± 0.87 | 4.73 ± 1.02 | |||
| Ash | 18.06 ± 1.47 | 24.73 ± 1.58 | 15.61 ± 1.92 | |||
| Protein | 3.67 ± 0.56 | 6.02 ± 0.06 | 19.28 ± 0.05 | |||
| Lipids | 1.96 ± 0.14 | 2.91 ± 0.08 | 3.04 ± 0.32 | |||
| Insoluble fibre | 37.41 ± 0.72 | 27.82 ± 0.21 | 32.01 ± 0.34 | 50.49 ± 1.81 | 63.09 ± 1.74 | 59.61 ± 1.17 |
| Soluble fibre | 23.47 ± 0.60 | 13.46 ± 0.45 | 15.05 ± 0.48 | 31.37 ± 0.56 | 30.86 ± 0.96 | 28.32 ± 0.30 |
| Total polysaccharides * | 62.53 ± 1.38 | 58.85 ± 1.49 | 50.14 ± 0.87 | 83.71 ± 2.32 | 94.56 ± 2.07 | 91.35 ± 1.86 |
| P. comosa | E. radiata | U. ohnoi | |
|---|---|---|---|
| (mg PE/g) | (mg GAE/g) | ||
| WH seaweeds | 0.38 ± 0.02 | 0.55 ± 0.03 | 0.35 ± 0.02 |
| PP extracts | 4.33 ± 0.15 | 3.48 ± 0.27 | 2.46 ± 0.21 |
| Rhamnose | Fucose | Ribose | Arabinose | Xylose | Mannose | Galactose | Glucose | ||
|---|---|---|---|---|---|---|---|---|---|
| Soluble NNSP (% DW) | Total Soluble NNSP (% DW) | ||||||||
| P. comosa WH | 0.00 ± 0.00 | 3.31 ± 0.14 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.34 ± 0.07 | 0.59 ± 0.05 | 0.52 ± 0.03 | 0.25 ± 0.00 | 6.00 |
| P. comosa PS | 0.00 ± 0.00 | 5.05 ± 0.21 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.59 ± 0.13 | 1.09 ± 0.07 | 0.92 ± 0.08 | 0.52 ± 0.01 | 10.17 |
| E. radiata WH | 0.00 ± 0.00 | 0.98 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.21 ± 0.01 | 0.04 ± 0.00 | 0.31 ± 0.00 | 0.39 ± 0.00 | 1.93 |
| E. radiata PS | 0.00 ± 0.00 | 1.39 ± 0.08 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.35 ± 0.01 | 0.08 ± 0.00 | 0.59 ± 0.02 | 0.61 ± 0.03 | 3.02 |
| U. ohnoi WH | 0.58 ± 0.06 | 0.00 ± 0.00 | 0.09 ± 0.01 | 0.00 ± 0.00 | 0.10 ± 0.00 | 0.09 ± 0.01 | 0.02 ± 0.00 | 0.00 ± 0.00 | 0.89 |
| U. ohnoi PS | 1.12 ± 0.09 | 0.00 ± 0.00 | 0.16 ± 0.02 | 0.00 ± 0.00 | 0.17 ± 0.01 | 0.17 ± 0.02 | 0.04 ± 0.00 | 0.00 ± 0.00 | 1.66 |
| Insoluble NNSP (% DW) | Total insoluble NNSP (% DW) | ||||||||
| P. comosa WH | 0.00 ± 0.00 | 1.91 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.65 ± 0.03 | 0.16 ± 0.00 | 0.00 ± 0.00 | 3.36 ± 0.27 | 6.08 |
| P. comosa PS | 0.00 ± 0.00 | 3.32 ± 0.07 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.17 ± 0.14 | 0.28 ± 0.01 | 0.00 ± 0.00 | 6.14 ± 0.58 | 10.91 |
| E. radiata WH | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.60 ± 0.09 | 2.60 |
| E. radiata PS | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 4.72 ± 0.13 | 4.72 |
| U. ohnoi WH | 3.04 ± 0.09 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.20 ± 0.06 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.75 ± 0.06 | 6.99 |
| U. ohnoi PS | 4.01 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.28 ± 0.09 | 0.00 ± 0.00 | 0.00 ± 0.00 | 3.68 ± 0.17 | 9.98 |
| P. comosa | E. radiata | U. ohnoi | |
|---|---|---|---|
| Mineral | mg/g (DW) | ||
| Bromine | 0.35 | 0.34 | 0.06 |
| Calcium | 13.00 | 11.20 | 2.79 |
| Chloride | 47.7 | 101.0 | 47.5 |
| Chromium | 0.00031 | 0.0032 | 0.00043 |
| Copper | 0.00014 | 0.00093 | 0.013 |
| Iodine | 1.70 | 3.40 | 0.0029 |
| Iron | 0.023 | 0.88 | 0.19 |
| Magnesium | 7.22 | 5.76 | 32.00 |
| Manganese | 0.0095 | 0.0074 | 0.017 |
| Molybdenum | 0.00035 | 0.00031 | 0.00023 |
| Phosphorus | 1.46 | 1.45 | 2.17 |
| Potassium | 68.10 | 78.00 | 25.00 |
| Selenium | <0.005 | <0.005 | <0.005 |
| Sodium | 26.3 | 30.2 | 23.1 |
| Zinc | 0.029 | 0.019 | 0.041 |
| µmol TE/g (DW) | ||
|---|---|---|
| Whole | Polyphenol Extract | |
| P. comosa | 84.5 ± 2.32 | 224.7 ± 5.33 |
| E. radiata | 111.0 ± 2.46 | 345.4 ± 6.87 |
| U. ohnoi | 17.5 ± 1.05 | 59.1 ± 1.96 |
| Seaweed | Gastrically Digested Portion (% DW) |
|---|---|
| P. comosa WH | 20.28 ± 0.93 |
| E. radiata WH | 41.98 ± 1.84 |
| U. ohnoi WH | 26.90 ± 1.07 |
| P. comosa PS | 9.36 ± 0.88 |
| E. radiata PS | 11.77 ± 1.94 |
| U. ohnoi PS | 8.38 ± 1.53 |
| Substrate | Total SCFA | Acetic | Butyric | Propionic | iso-Butyric | iso-Valeric | Valeric | Caproic |
|---|---|---|---|---|---|---|---|---|
| Blank | 33.53 ± 2.03 b | 18.33 ± 1.29 b | 5.17 ± 0.37 a | 5.29 ± 0.19 b | 1.17 ± 0.11 a | 1.80 ± 0.05 b | 1.76 ± 0.02 a | 0.00 ± 0.00 a |
| Cellulose | 35.55 ± 1.45 b | 18.37 ± 1.02 b | 5.87 ± 0.19 a | 5.91 ± 0.10 b | 1.42 ± 0.02 a | 2.06 ± 0.05 b | 1.92 ± 0.05 a | 0.00 ± 0.00 a |
| Inulin | 71.05 ± 1.08 d | 50.33 ± 0.77 c | 7.32 ± 0.06 a | 13.00 ± 0.25 d | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.40 ± 0.01 a | 0.00 ± 0.00 a |
| EGCG | 7.76 ± 0.09 a | 5.72 ± 0.06 a | 0.99 ± 0.02 a | 1.06 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| P. comosa WH | 52.32 ± 0.87 bc | 32.68 ± 0.46 bc | 8.11 ± 0.28 a | 6.78 ± 0.08 b | 0.94 ± 0.03 a | 1.49 ± 0.02 b | 2.09 ± 0.00 a | 0.23 ± 0.00 a |
| P. comosa PS | 37.38 ± 0.74 b | 24.27 ± 0.23 b | 4.64 ± 0.12 a | 6.53 ± 0.07 b | 0.30 ± 0.26 a | 0.71 ± 0.03 ab | 0.71 ± 0.03 a | 0.22 ± 0.01 a |
| P. comosa PP | 49.50 ± 1.28 bc | 24.30 ± 0.48 b | 13.71 ± 0.58 ab | 9.97 ± 0.15 c | 0.00 ± 0.00 a | 0.45 ± 0.01 ab | 1.08 ± 0.05 a | 0.00 ± 0.00 a |
| E. radiata WH | 61.20 ±1.02 cd | 37.56 ± 0.75 bc | 9.45 ± 0.43 ab | 8.96 ± 0.34 c | 1.00 ± 0.09 a | 1.65 ± 0.14 b | 2.27 ± 0.01 a | 0.29 ± 0.01 a |
| E. radiata PS | 227.53 ± 5.39 g | 145.03 ± 4.38 g | 30.04 ± 0.40 c | 31.35 ± 0.25 f | 4.40 ± 0.10 b | 6.60 ± 0.13 c | 8.58 ± 0.07 c | 1.53 ± 0.06 c |
| E. radiata PP | 183.73 ± 20.06 f | 99.44 ± 1.62 e | 44.87 ± 11.22 d | 28.62 ± 2.10 f | 0.97 ± 1.68 a | 2.74 ± 1.59 b | 6.09 ± 1.79 bc | 0.99 ± 0.07 b |
| U. ohnoi WH | 208.68 ± 19.08 fg | 119.52 ± 12.28 f | 33.07 ± 1.87 c | 32.69 ± 1.99 f | 4.95 ± 0.77 b | 8.07 ± 0.89 d | 10.12 ± 0.83 c | 0.26 ± 0.45 a |
| U. ohnoi PS | 182.91 ± 3.47 f | 104.83 ± 2.39 e | 27.88 ± 0.37 c | 28.78 ± 0.25 f | 4.38 ± 0.14 b | 6.96 ± 0.16 c | 8.98 ± 0.11 c | 1.10 ± 0.06 b |
| U. ohnoi PP | 140.42 ± 2.93 e | 77.07 ± 1.58 d | 27.13 ± 0.13 c | 29.47 ± 0.89 f | 0.00 ± 0.00 a | 2.05 ± 0.09 b | 4.70 ± 0.23 b | 0.00 ± 0.00 a |
| WH | PS | PP | |
| P. comosa | 0.794/0.332 (2.391) | 0.757/0.247 (3.065) | 0.573/0.249 (2.301) |
| E. radiata | 0.816/0.334 (2.443) | 0.804/0.319 (2.520) | 0.655/0.245 (2.673) |
| U. ohnoi | 0.777/0.252 (3.083) | 0.773/0.074 (10.446) | 0.688/0.146 (4.712) |
| Controls | |||
| Inulin | 0.325/0.687 (0.473) | ||
| EGCG | 0.671/0.436 (1.539) | ||
| Cellulose | 0.778/0.241(3.228) | ||
| Blank | 0.763/0.219 (3.484) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shannon, E.; Conlon, M.; Hayes, M. The Prebiotic Effect of Australian Seaweeds on Commensal Bacteria and Short Chain Fatty Acid Production in a Simulated Gut Model. Nutrients 2022, 14, 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102163
Shannon E, Conlon M, Hayes M. The Prebiotic Effect of Australian Seaweeds on Commensal Bacteria and Short Chain Fatty Acid Production in a Simulated Gut Model. Nutrients. 2022; 14(10):2163. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102163
Chicago/Turabian StyleShannon, Emer, Michael Conlon, and Maria Hayes. 2022. "The Prebiotic Effect of Australian Seaweeds on Commensal Bacteria and Short Chain Fatty Acid Production in a Simulated Gut Model" Nutrients 14, no. 10: 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102163