1. Introduction
The prevalence of type 2 diabetes mellitus (DM) has increased over short periods and occurs at a relatively younger age and lower body-mass index in Asia [
1]. When people in Asia develop diabetes with a lower degree of obesity at younger ages, they suffer longer from diabetic complications and die sooner than people in other regions [
2]. In Taiwan, DM was the fifth leading cause of death in 2015, and its prevalence has been increasing [
3]. Complications of DM include macrovascular and microvascular complications such as cardiovascular disease, chronic kidney failure, and diabetic retinopathy [
4,
5,
6]. Those complications are attributed to hyperglycemia inducing oxidative stress. Hyperglycemia increases NADH and FADH
2 and inhibits the delivery of protons through complex III in electron transport chain, which leads to overproduction of reactive oxygen species (ROS) and oxidative stress [
7]. ROS are highly toxic to cell components, particularly to cell membranes composed of lipids. ROS destroy lipids on membranes and produce lipid peroxide which damages cells. ROS are metabolized by nonenzymatic and enzymatic mechanisms to eliminate oxidative stress [
8].
Melatonin (5-methoxy-
N-acetyltryptamine), which is one of the strongest antioxidants, is secreted with a daily rhythm by the pineal gland [
9]. The peak concentration is around 10 pg/mL (43 pmol/L) in blood and 3 pg/mL in the saliva [
10]. It is thought that melatonin may be useful in the management of several diseases, such as depression [
11], insomnia [
11,
12], obesity [
9,
13], diabetes [
13], cancer [
12], and immune [
14] and cardiac disorders [
15]. It can scavenge ROS by its antioxidant properties and increase antioxidant enzyme activities [
16]. A recent study indicated that animals which received a pinealectomy exhibited impaired glucose tolerance, insulin resistance, and diabetes, which were improved by treatment with melatonin [
17]. In high-cholesterol diet rat model, supplementation with melatonin could decrease plasma total cholesterol and liver cholesterol and triglyceride, and increase plasma high-density lipoprotein-cholesterol (HDL-C) [
18]. In Zucker diabetic fatty (ZDF) rats, supplementation with melatonin not only decreased elevated plasma leptin, insulin and high blood glucose, but increased low levels of adiponectin [
13]. In a rat model of streptozotocin (STZ)-induced DM, supplementation with melatonin decreased fasting blood glucose (FBG) and improved liver damage, and reduced the oxidative stress [
19].
To induce diabetes, it is common to use STZ in a rodent model. STZ, an antibiotic produced by
Streptomyces achromogens, is a 2-deoxy-
d-glucose derivative, and because its structure is like that of glucose, it can cross glucose transporter 2 on islets [
20]. The damage to pancreatic cells by STZ was proposed to occur by two mechanisms. One is that STZ is a strong alkylating agent, which directly alkylates DNA by •CH3 or CH
3+ when STZ decomposes. The other is that STZ generates ROS in diabetogenesis [
21]. However, Murata et al. indicated that diabetogenesis of STZ is mainly caused by alkylation of DNA by CH
3+ [
22]. A method of inducing type 2 DM with hyperglycemia and relatively low insulin levels can be produced by combining STZ and nicotinamide (NA) [
23,
24]. In an ICR mice model, the combination of STZ and NA resulted in moderate hyperglycemia and metabolic syndrome [
25].
The intervention with melatonin can improve FBG, insulin, adipokines, and the oxidative state in a DM rat model [
13,
19]. However, few studies have evaluated the intervention with melatonin in a DM mouse model, and how melatonin affects glucose levels in an STZ-induced diabetic model is still controversial. These experiments were carried out to determine the effects of different doses of melatonin on glucose tolerance, lipid profile, adipokines, and activities of antioxidant enzymes in mice with NA/STZ-induced hyperglycemia.
2. Materials and Methods
2.1. Chemicals
STZ and melatonin were purchased from Sigma Chemical (St. Louis, MO, USA).
2.2. Animals
Male ICR mice (28–30 g body weight (BW)) were obtained at an age of 8 weeks from BioLASCO Taiwan (Taipei, Taiwan). Mice were maintained on commercial standard chow and tap water ad libitum. Mice were housed in a temperature- (22 ± 1 °C) and humidity- (40–60%) controlled room with a 12-h dark/light cycle (lights on at 07:00). BWs were recorded weekly throughout the experiments. After 1 week of acclimatization, animals were randomly divided into five groups including control (C), diabetic (D), low-dosage (L), medium-dosage (M), and high-dosage (H) groups. Groups C and D received the vehicle (a 6% alcohol (w/v) aqueous solution) daily. Groups L, M, and H, respectively, received melatonin dissolved in a 6% alcohol solution at 10, 20, and 50 mg/kg of BW daily after inducing hyperglycemia immediately. The vehicle and melatonin solution were given by oral tube feeding between 17:00 and 19:00 daily for 6 weeks. At the end of the experiment, mice were sacrificed by anesthetization. In addition, six normal male ICR mice (35–45 g BW) were used to study the pharmacokinetics of melatonin. This study was approved by the Institutional Animal Care and Use Committee (IACUC) of the Institute for Experimental Medical Research, Taipei Medical University (Taipei, Taiwan; LAC-2016-0173).
2.3. Induction of Hyperglycemia
The D, L, M and H groups were intraperitoneally (i.p.) injected with STZ (50 mg/kg of BW) in 0.1 M citrate buffer (pH 4.2) on two consecutive days. NA (120 mg/kg of BW) in saline was i.p. injected 30 min before the STZ injection on the first day after overnight fasting. Group C was i.p. injected with citrate buffer. Seven days after the second i.p. injection, mice that exhibited an 8-h FBG level of ≥200 mg/dL were recognized as being hyperglycemic. The others that exhibited an FBG level of <200 mg/dL were injected with STZ and monitored until the FBG level reached ≥200 mg/dL. FBG was monitored with a glucometer (Dragon Pharmaceutical Co, New Taipei, Taiwan). After D, L, M and H groups were induced hyperglycemia, melatonin administration was carried out at the same time.
2.4. Oral Glucose Tolerance Test (OGTT)
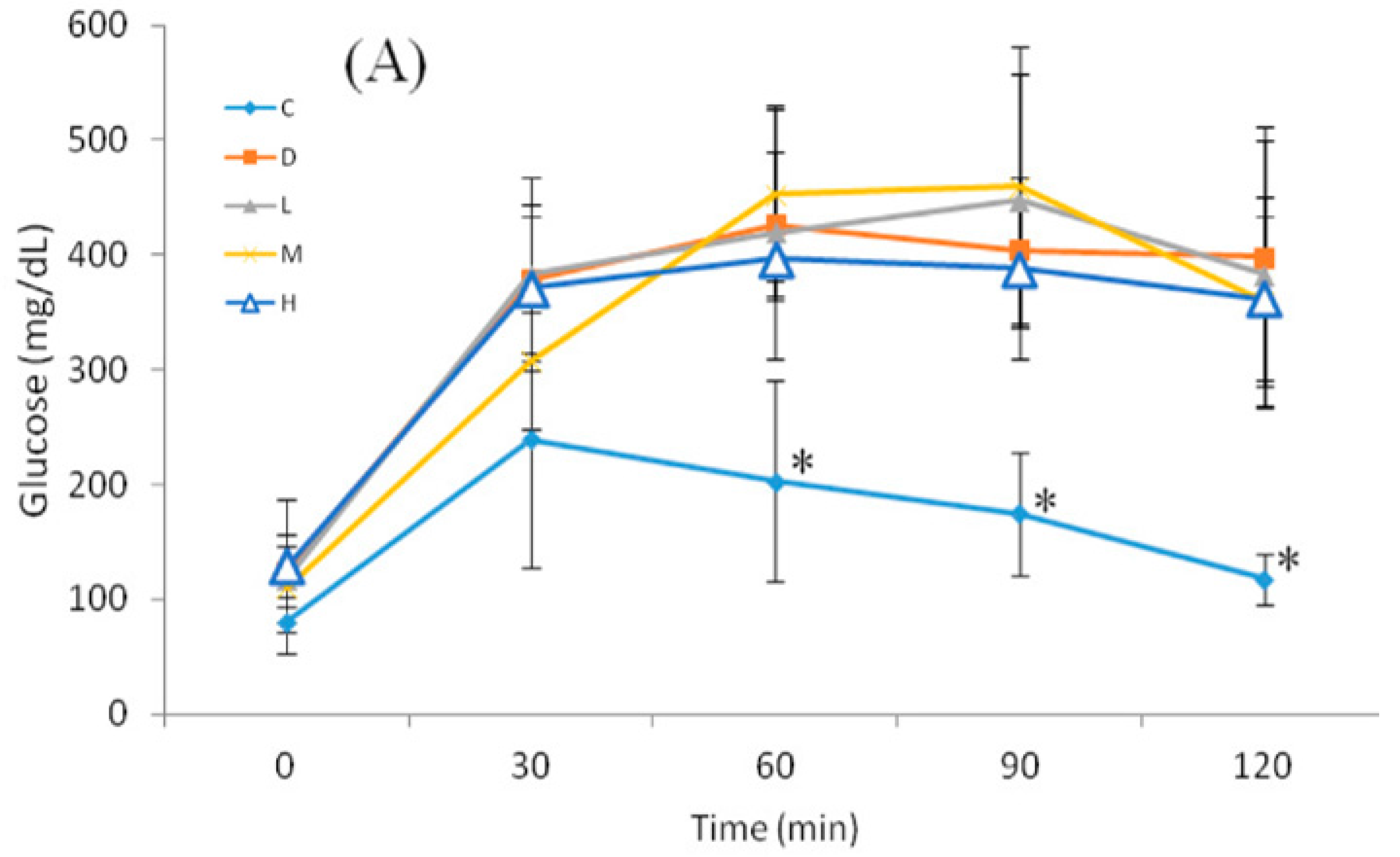
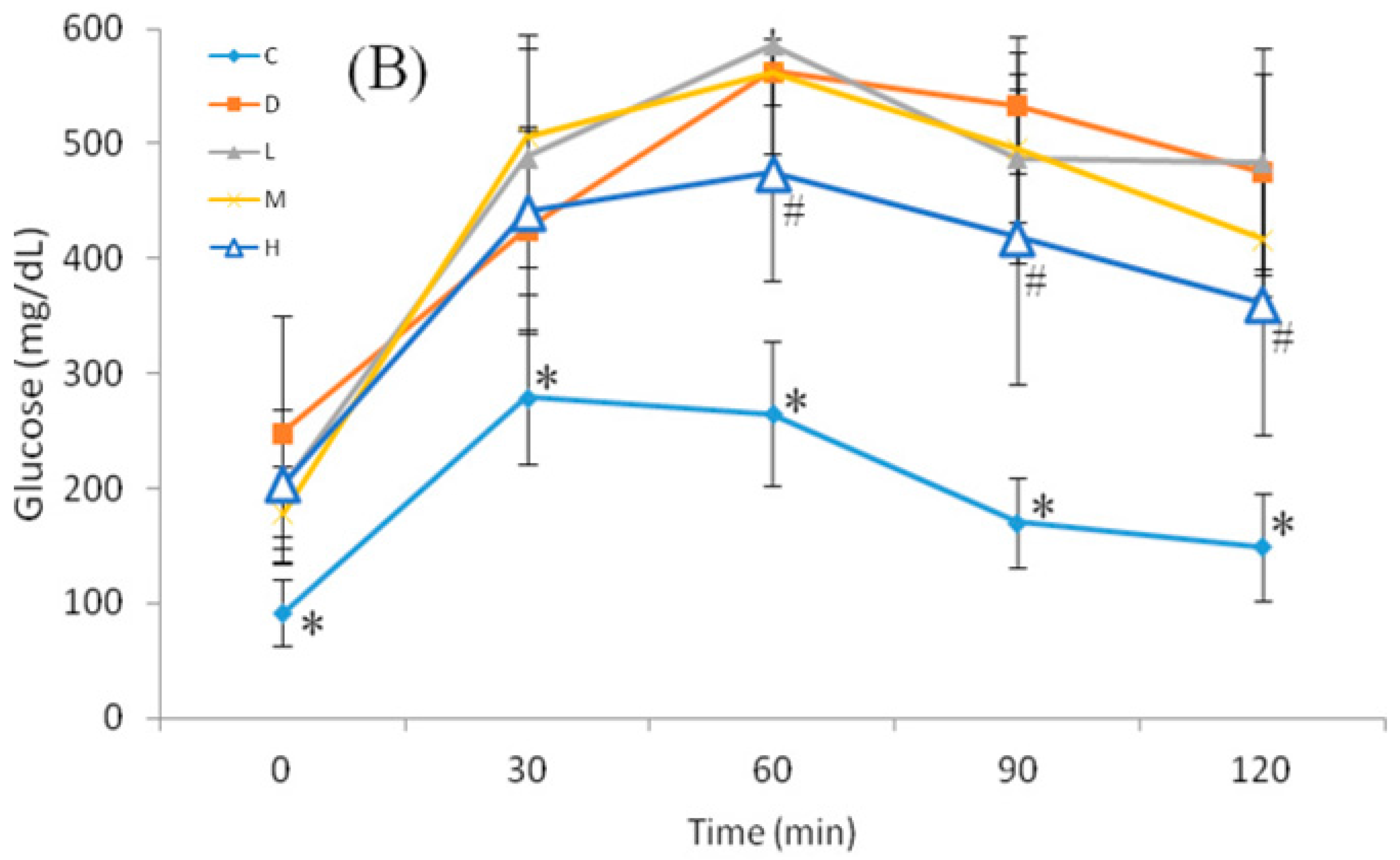
The OGTT was performed at 09:00, and 12-h FBG of mice was monitored at 0 min. The glucose load (2 g/kg of BW) was given as a bolus by gavage, and blood sugar was monitored at 30, 60, 90, and 120 min after glucose administration.
2.5. Pharmacokinetics of Melatonin
Six mice were starved for 4 h before being given melatonin by gavage. Melatonin was prepared as previously described. On Day 1, mice were tube fed melatonin at a dosage of 10 mg/kg of BW. Blood samples were collected from a tail vein at 10, 30, 60, and 120 min after melatonin administration. Blood samples were centrifuged at 1000× g for 30 min, and supernatants were collected to determine the melatonin concentration. On Day 5, the same six mice were administered melatonin at 50 mg/kg of BW and then followed the same steps as Day 1. The elimination rate constant (Ke) was calculated by (ln(Cp1)–ln(Cp2))/(t2–t1), where Cp1 is the melatonin concentration at t1, and Cp2 is the melatonin concentration at t2. The half-life (t1/2) was calculated by Ln(2)/Ke.
2.6. Biochemical Analysis
Plasma insulin was determined by a Mercodia mouse insulin enzyme-linked immunosorbent assay (ELISA) kit (Uppsala, Sweden). Plasma leptin and adiponectin were determined by an AssayMax mouse leptin and adiponectin ELISA kit (St. Charles, IL, USA). Plasma and serum melatonin were determined by a mouse melatonin ELISA kit (MyBioSource, Inc., San Diego, CA, USA). Plasma and liver total cholesterol and triglyceride were determined by a colorimetric assay kit. The homeostasis model assessment for insulin resistance (HOMA-IR) and quantitative insulin sensitivity check index (QUICKI) were calculated using the fasting glucose level (mg/dL) and insulin concentration (µU/mL). The HOMA-IR was calculated by (fasting blood sugar × insulin)/405. QUICKI was calculated by 1/(log(fasting blood sugar) + log(insulin)). Malondialdehyde (MDA) in liver homogenates and plasma was measured by a thiobarbituric acid-reactive substance (TBARS) assay kit from Cayman Chemical (Ann Arbor, MI, USA). Activities of superoxide dismutase (SOD) and glutathione peroxidase (GPx) in liver homogenates were measured by kits from Cayman Chemical. Total protein concentrations were determined by Bio-Rad protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
2.7. Statistical Analysis
Data were analyzed using PASW Statistics software (vers. 18.0, SPSS, Chicago, IL, USA). Comparisons between groups were made with an analysis of variance (ANOVA) followed by Duncan’s multiple-range test. A p value of <0.05 was considered significant.
4. Discussion
Melatonin is a biological modulator of mood, sleep, sexual behavior and circadian rhythm at physiological concentration in human [
16]. However, melatonin secretion decreases with age along with increased insulin resistance, but supplement of melatonin can relieve those symptoms [
16]. In a published cell experiment, 10 nM melatonin increased the ability of glucose intake in 3T3-L1 adipocytes [
26]. In this study, administration of melatonin at 50 mg/kg of BW increased and maintained serum melatonin levels ranging 1.46–11.67 µM, which was 1000 times than nM. It might indicate that administration of melatonin at 50 mg/kg can reach effective concentration of melatonin. After STZ induction of diabetes, rats exhibited high blood glucose and low plasma insulin levels. Moreover, those rats also exhibited low melatonin levels [
27]. This study showed that Group D exhibited low plasma melatonin levels and high glucose levels after NA/STZ induction, and an oral melatonin intervention recovered plasma melatonin levels. According to previous studies, melatonin administration can reduce BW, fat mass, FBG, and insulin levels in gene- or diet-induced diabetic [
13,
28,
29,
30,
31,
32] obese rodents. Nevertheless, it is still controversial that the effects of melatonin on blood glucose in an STZ-induced diabetic model. Several researchers reported that supplementation with melatonin did not lower FBG in an STZ diabetic rat model [
33,
34,
35,
36]. In contrast, other researchers reported that 10–20 mg/kg melatonin exerted lower FBG in an STZ- or alloxan-induced diabetic rat model [
37,
38,
39]. The current study showed that oral melatonin administration did not improve FBG and HOMA-IR in mice with NA/STZ-induced hyperglycemia, which corresponded to the latter results for FBG. However, Group H showed a significantly lower AUC compared to Group D. This may indicate that melatonin delayed diabetic progression by increasing glucose sensitivity and the insulin level.
Hyperglycemia usually results in hypertriglyceridemia and hypercholesterolemia [
40]. STZ is able to induce hyperglycemia and results in hypertriglyceridemia and hypercholesterolemia [
25]. In this study, plasma triglyceride and cholesterol and the hepatic cholesterol level in Group D significantly increased. In previous studies, melatonin administration lowered plasma triglyceride and cholesterol and increased plasma HDL-C level at 10–20 mg/kg in a gene- [
41] or diet-induced [
28,
29,
31] obese rat model and an STZ-induced diabetic rat model [
38,
39]. In high-cholesterol diet rat model, melatonin could decrease plasma total cholesterol and liver cholesterol and triglyceride, and increase plasma HDL-C, which exerted similar effect as hypocholesterolemia agents, cholestyramine [
18]. The current study showed that oral melatonin administration could increase plasma HDL-C level and lower triglyceride and cholesterol in both the plasma and liver of mice with NA/STZ-induced diabetes, which corresponded to previous studies. Melatonin could increase HDL-C level along with lower plasma cholesterol, which leading to lower total cholesterol/HDL-C ratio. In previous study, plasma total cholesterol/HDL-C ratio the strongest independent predictors of development for atherosclerosis [
42]. This may indicate that melatonin improve blood lipid composition, which is beneficial to diabetes patients.
In diabetic models, it is usually observed that oxidative stress increases. Several studies reported that increased MDA levels in diabetic models were decreased by melatonin, regardless of whether the glucose level was lower or not [
19,
30,
34,
37,
43,
44]. In this study, melatonin lowered MDA levels in the liver and plasma. However, SOD and GPx activities may increase [
19,
37,
44,
45], decrease [
44,
46] or remain unchanged [
43,
46] in diabetic models. In some research, melatonin did not increase SOD or GPx activities [
43,
46]. However, other researchers reported that melatonin increased SOD or GPx activities [
35,
37,
44]. In this study, both hepatic SOD and GPx activities were lower in Group D. Melatonin administration increased liver SOD activity but not GPx activity. Nevertheless, melatonin decreased MDA levels and increased melatonin levels in all intervention groups, but did not significantly increase SOD activities in groups L or M. Otherwise, because melatonin’s strong anti-oxidative ability, increased plasma melatonin levels in all intervention groups might also exert higher anti-oxidative ability. It was assumed that melatonin mainly reduced MDA levels by its anti-oxidative property in groups L and M, and reduced MDA levels by combining its anti-oxidative property with increasing SOD activity in Group H.
In a gene-induced obesity diabetic model, leptin secretion increased along with increasing fat, but adiponectin secretion decreased, leading to an increase in the leptin/adiponectin ratio. However, supplementation with melatonin can increase adiponectin and decrease leptin levels in this model [
13]. Moreover, supplementation with melatonin can decrease leptin secretion and increase adiponectin secretion in a diet-induced model [
28,
30,
31]. On the other hand, leptin and adiponectin decreased in an NA/STZ-induced rat model, and adipocytes exerted lower glucose intake and oxidation ability [
23,
24]. Few studies reported effects of melatonin on adipokines in an NA/STZ-induced diabetic model. De Oliveira et al. reported that supplementation with melatonin can increase adiponectin and lower leptin levels in a neonatal STZ-induced diabetic model [
47]. In this study, melatonin increased adiponectin in all intervention groups and lowered leptin levels in groups M and H, which corresponded to results of de Oliveira and coworkers’ study. The leptin/adiponectin ratio is considered a better anti-atherogenic index than adiponectin alone [
48]. In our study, melatonin lowered the leptin/adiponectin ratio in Group L, as it did in groups M and H compared to Group L.
Although oral administration of melatonin did not decrease blood glucose, it improved hyperglycemia-induced high oxidative stress, low adiponectin levels, and dyslipidemia. Otherwise, melatonin could improve blood lipid composition and decrease total cholesterol/HDL-c ratio, which was thought to reduce the risk atherogenesis. If diabetic patients receive an oral hypoglycemic agent with melatonin administration to control blood glucose, it might be beneficial to improve diabetes-related phenomena.






{kind=link}
{kind=link}