
After the Fall: The Demographic Destiny of a Gorgonian Population Stricken by Catastrophic Mortality

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
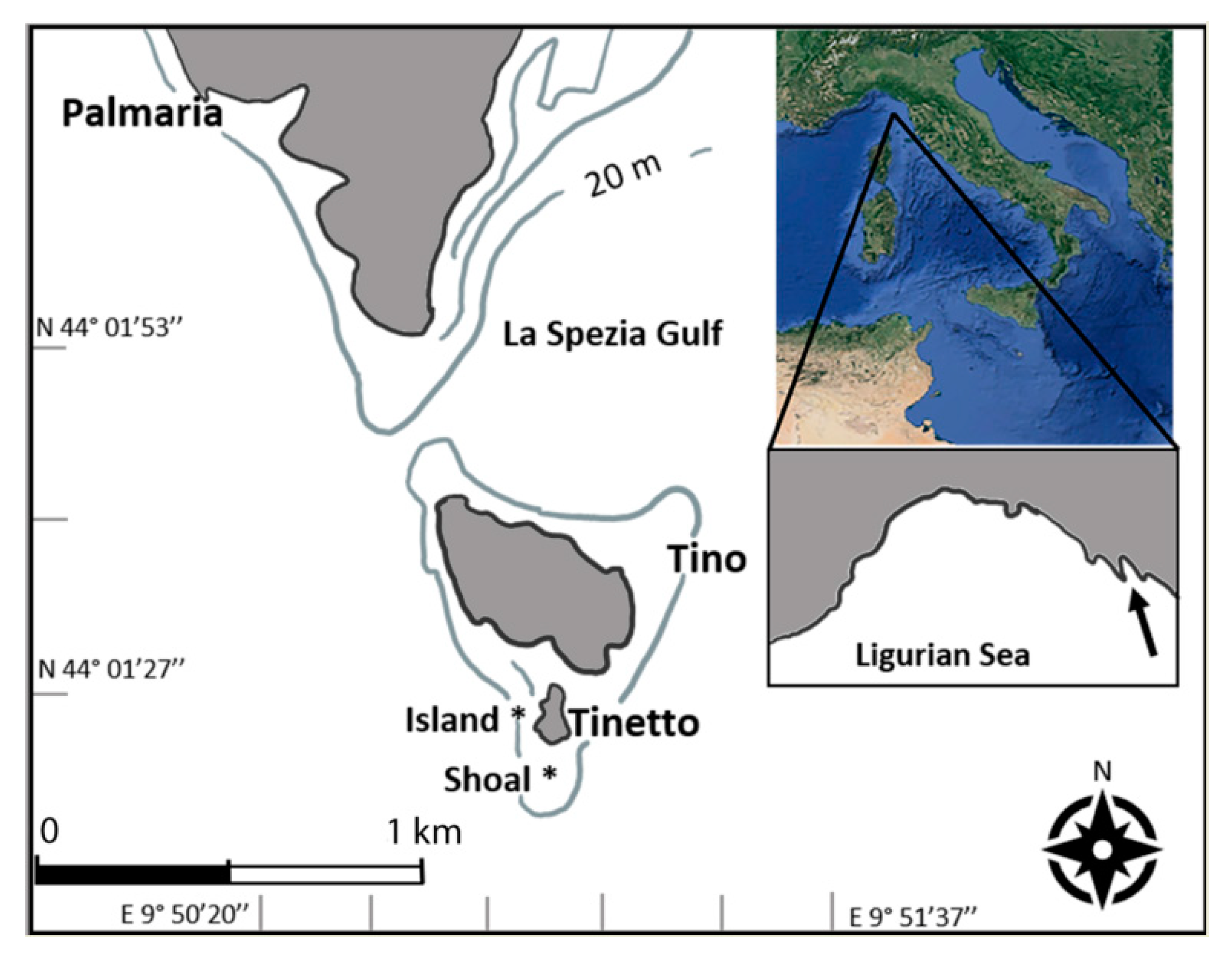
2.1. Study Area and Sampling Design
2.2. Size Structure and Density of the P. clavata Population
2.3. Data Analysis
3. Results
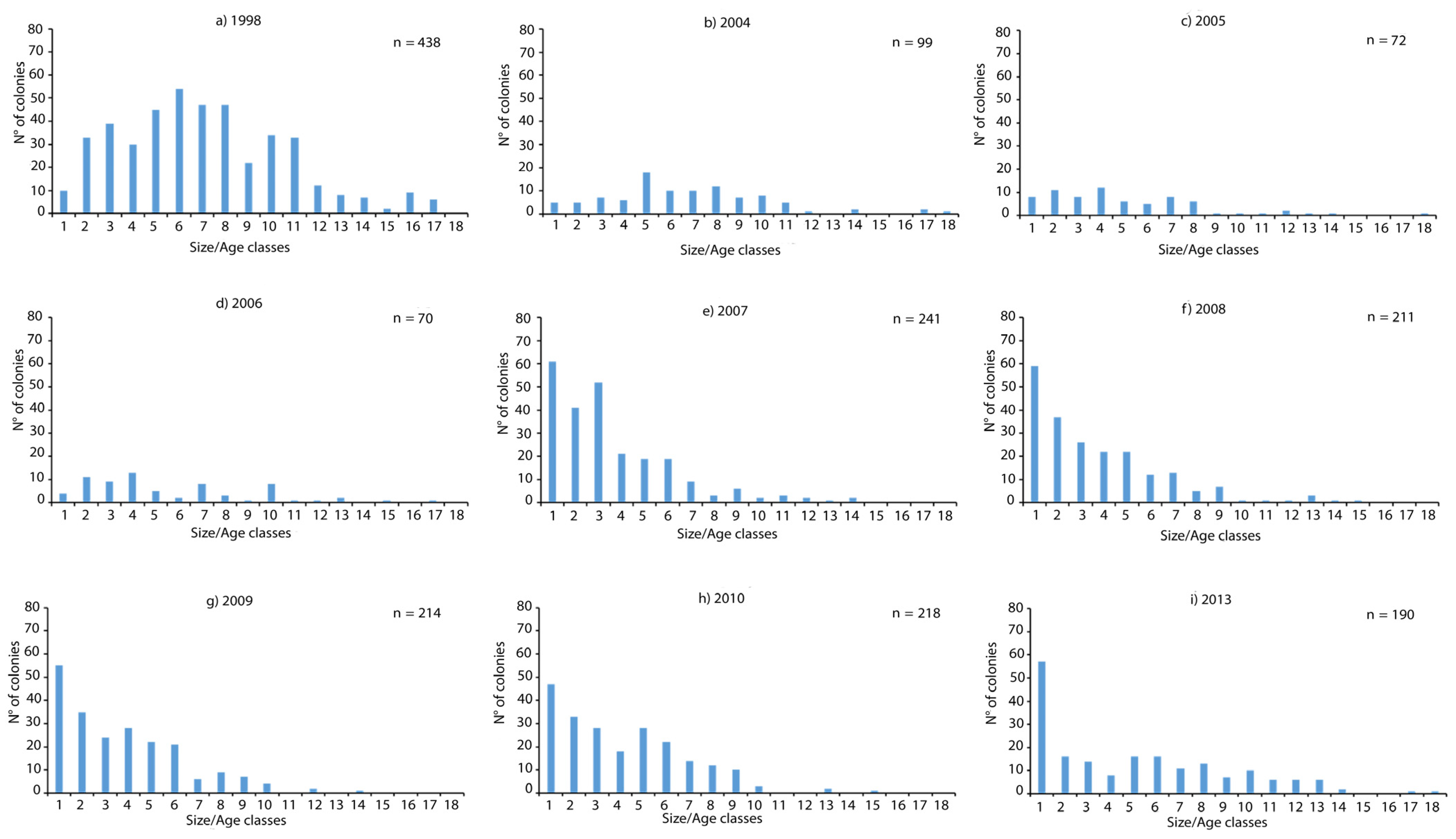
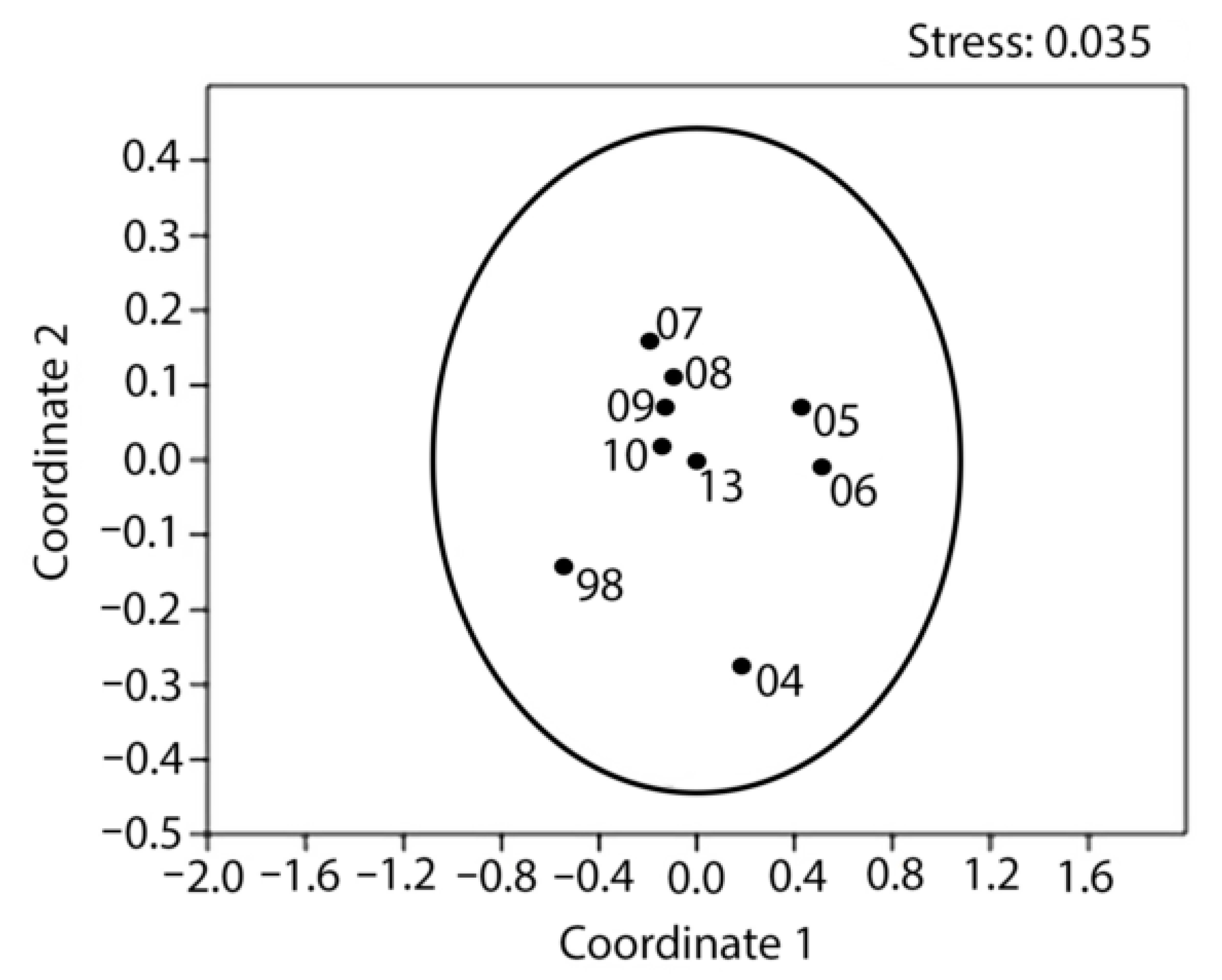
3.1. Population Density and Size/Age Structure over the 1998–2013 Period
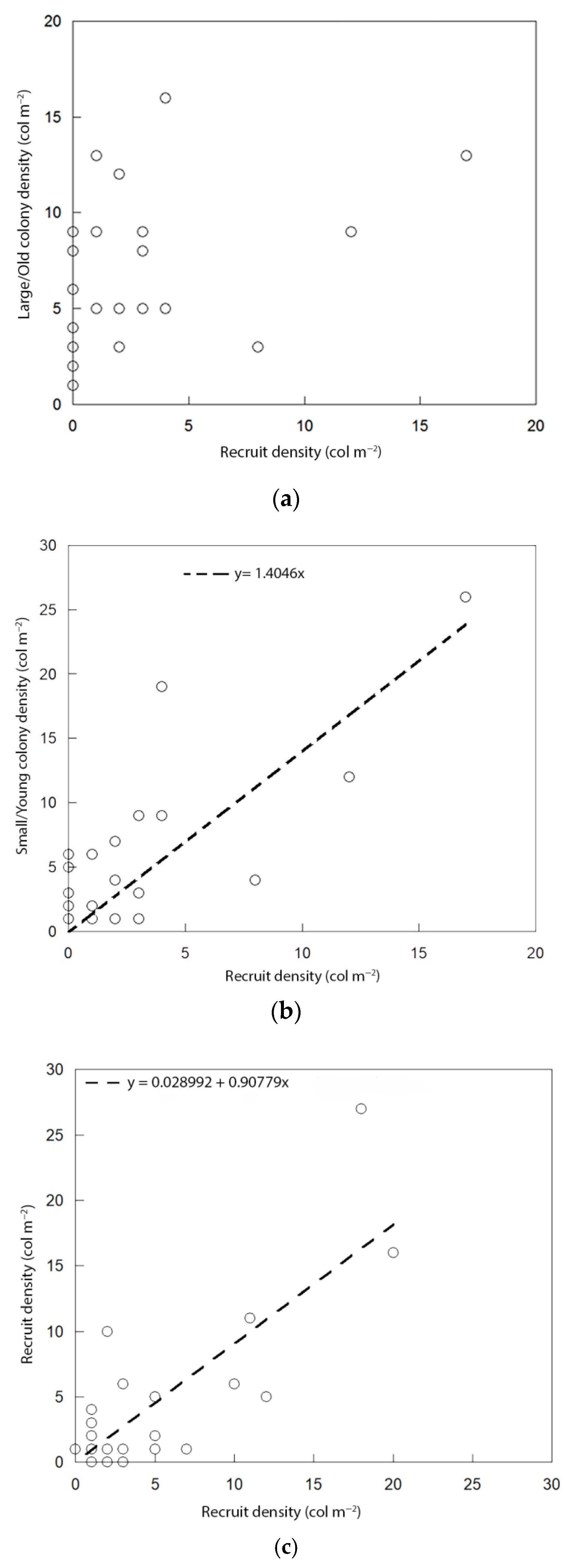
3.2. Percentage Cover of Benthic Organisms and Density of Paramuricea Clavata
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frölicher, T.L.; Laufkötter, C. Emerging risks from marine heat waves. Nat. Commun. 2018, 9, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Decarlo, T.M.; Cohen, A.L.; Wong, G.T.F.; Davis, K.A.; Lohmann, P.; Soong, K. Mass coral mortality under local amplification of 2 °C ocean warming. Sci. Rep. 2017, 7, srep44586. [Google Scholar] [CrossRef]
- Langangen, Ø.; Ohlberger, J.; Stige, L.C.; Durant, J.M.; Ravagnan, E.; Stenseth, N.C.; Hjermann, D.Ø. Cascading effects of mass mortality events in Arctic marine communities. Glob. Chang. Biol. 2016, 23, 283–292. [Google Scholar] [CrossRef]
- Verdura, J.; Linares, C.; Ballesteros, E.; Coma, R.; Uriz, M.J.; Bensoussan, N.; Cebrian, E. Biodiversity loss in a Mediterranean ecosystem due to an extreme warming event unveils the role of an engineering gorgonian species. Sci. Rep. 2019, 9, 5911. [Google Scholar] [CrossRef]
- Lirman, D.; Schopmeyer, S.; Manzello, D.; Gramer, L.J.; Precht, W.F.; Muller-Karger, F.; Banks, K.; Barnes, B.; Bartels, E.; Bourque, A.; et al. Severe 2010 Cold-Water Event Caused Unprecedented Mortality to Corals of the Florida Reef Tract and Reversed Previous Survivorship Patterns. PLoS ONE 2011, 6, e23047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danovaro, R.; Dell’Anno, A.; Fabiano, M.; Pusceddu, A.; Tselepides, A. Deep-sea ecosystem response to climate changes: The eastern Mediterranean case study. Trends Ecol. Evol. 2001, 16, 505–510. [Google Scholar] [CrossRef]
- Teagle, H.; Smale, D.A. Climate-driven substitution of habitat-forming species leads to reduced biodiversity within a temperate marine community. Divers. Distrib. 2018, 24, 1367–1380. [Google Scholar] [CrossRef] [Green Version]
- Marshall, P.A.; Baird, A.H. Bleaching of corals on the Great Barrier Reef: Differential susceptibilities among taxa. Coral Reefs 2000, 19, 155–163. [Google Scholar] [CrossRef]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; Van Woesik, R. Coral bleaching: The winners and the losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Brown, B.E.; Dunne, R.P.; Goodson, M.S.; Douglas, A.E. Experience shapes the susceptibility of a reef coral to bleaching. Coral Reefs 2002, 21, 119–126. [Google Scholar] [CrossRef]
- Garzón-Ferreira, J.; Zea, S. A mass mortality of Gorgonia ventalina (Cnidaria: Gorgoniidae) in the Santa Marta area, Caribbean coast of Colombia. Bull. Mar. Sci. 1992, 50, 522–526. [Google Scholar]
- Santangelo, G.; Bramanti, L.; Iannelli, M. Population dynamics and conservation biology of the over-exploited Mediterranean red coral. J. Theor. Biol. 2007, 244, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, G.; Cupido, R.; Cocito, S.; Bramanti, L.; Priori, C.; Erra, F.; Iannelli, M. Effects of increased mortality on gorgonian corals (Cnidaria, Octocorallia): Different demographic features may lead affected populations to unexpected recovery and new equilibrium points. Hydrobiologia 2015, 759, 171–187. [Google Scholar] [CrossRef]
- Linares, C.; Doak, D.F. Forecasting the combined effects of disparate disturbances on the persistence of long-lived gorgonians: A case study of Paramuricea clavata. Mar. Ecol. Prog. Ser. 2010, 402, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Prada, C.; Weil, E.; Yoshioka, P.M. Octocoral bleaching during unusual thermal stress. Coral Reefs 2010, 29, 41–45. [Google Scholar] [CrossRef]
- Sánchez, J.A.; Ardila, N.E.; Andrade, J.; Dueñas, L.F.; Navas, R.; Ballesteros, D. Octocoral densities and mortalities in Gorgona Island, Colombia, Tropical Eastern Pacific. Rev. Biol. Trop. 2014, 62, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Arizmendi-Mejía, R.; LeDoux, J.-B.; Civit, S.; Antunes, A.; Thanopoulou, Z.; Garrabou, J.; Linares, C. Demographic responses to warming: Reproductive maturity and sex influence vulnerability in an octocoral. Coral Reefs 2015, 34, 1207–1216. [Google Scholar] [CrossRef]
- Bramanti, L.; Benedetti, M.C.; Cupido, R.; Cocito, S.; Priori, C.; Erra, F.; Iannelli, M.; Santangelo, G. Demography of Animal Forests: The Example of Mediterranean Gorgonians. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 529–548. [Google Scholar]
- Tsounis, G.; Edmunds, P.J. Three decades of coral reef community dynamics in St. John, USVI: A contrast of scleractinians and octocorals. Ecosphere 2017, 8, e01646. [Google Scholar] [CrossRef]
- Cupido, R.; Cocito, S.; Barsanti, M.; Sgorbini, S.; Peirano, A.; Santangelo, G. Unexpected long-term population dynamics in a canopy-forming gorgonian coral following mass mortality. Mar. Ecol. Prog. Ser. 2009, 394, 195–200. [Google Scholar] [CrossRef]
- Cupido, R.; Cocito, S.; Manno, V.; Ferrando, S.; Peirano, A.; Iannelli, M.; Bramanti, L.; Santangelo, G. Sexual structure of a highly reproductive, recovering gorgonian population: Quantifying reproductive output. Mar. Ecol. Prog. Ser. 2012, 469, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Bramanti, L.; Edmunds, P.J. Density-associated recruitment mediates coral population dynamics on a coral reef. Coral Reefs 2016, 35, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Riegl, B.; Johnston, M.; Purkis, S.; Howells, E.; Burt, J.; Steiner, S.C.; Sheppard, C.R.C.; Bauman, A. Population collapse dynamics in Acropora downingi, an Arabian/Persian Gulf ecosystem-engineering coral, linked to rising temperature. Glob. Chang. Biol. 2018, 24, 2447–2462. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S. The destruction of the ‘animal forests’ in the oceans: Towards an over-simplification of the benthic ecosystems. Ocean Coast. Manag. 2013, 84, 77–85. [Google Scholar] [CrossRef]
- Bavestrello, G.; Bertone, S.; Cattaneo-Vietti, R.; Cerrano, C.; Gaino, E.; Zanzi, D. Mass mortality of Paramuricea clavata (Anthozoa, Cnidaria) on Portofino Promontory cliffs, Ligurian Sea, Mediterranean Sea. Mar. Life 1994, 4, 15–19. [Google Scholar]
- Huete-Stauffer, C.; Vielmini, I.; Palma, M.; Navone, A.; Panzalis, P.; Vezzulli, L.; Misic, C.; Cerrano, C. Paramuricea clavata (Anthozoa, Octocorallia) loss in the Marine Protected Area of Tavolara (Sardinia, Italy) due to a mass mortality event. Mar. Ecol. 2011, 32, 107–116. [Google Scholar] [CrossRef]
- Turicchia, E.; Abbiati, M.; Sweet, M.; Ponti, M. Mass mortality hits gorgonian forests at Montecristo Island. Dis. Aquat. Org. 2018, 131, 79–85. [Google Scholar] [CrossRef]
- Carpine, C.; Grasshoff, M. Les gorgonaires de la Mediterranee. Bull. Inst. Océanogr. (Monaco) 1975, 71, 1–140. [Google Scholar]
- Coma, R.; Zabala, M.; Gili, J.M. Sexual reproductive effort in the Mediterranean gorgonian Paramuricea clavata. Mar. Ecol. Prog. Ser. 1995, 117, 185–192. [Google Scholar] [CrossRef]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. 2006, 48, 123–195. [Google Scholar]
- Ponti, M.; Perlini, R.A.; Ventra, V.; Grech, D.; Abbiati, M.; Cerrano, C. Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PLoS ONE 2014, 9, e102782. [Google Scholar]
- Cupido, R.; Cocito, S.; Sgorbini, S.; Bordone, A.; Santangelo, G. Response of a gorgonian (Paramuricea clavata) population to mortality events: Recovery or loss? Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 984–992. [Google Scholar] [CrossRef]
- García-Gómez, J.C.; González, A.R.; Maestre, M.J.; Espinosa, F. Detect coastal disturbances and climate change effects in coralligenous community through sentinel stations. PLoS ONE 2020, 15, e0231641. [Google Scholar] [CrossRef]
- Padrón, M.; Costantini, F.; Bramanti, L.; Guizien, K.; Abbiati, M. Genetic connectivity supports recovery of gorgonian populations affected by climate change. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 776–787. [Google Scholar] [CrossRef] [Green Version]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-Vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-western Mediterranean), summer 1999. Ecol. Lett. 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Garrabou, J.; Coma, R.; Bensoussan, N.; Bally, M.; Chevaldonné, P.; Ciglianos, M.; Díaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: Effects of the 2003 heat wave. Glob. Chang. Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Bally, M.; Garrabou, J. Thermodependent bacterial pathogens and mass mortalities in temperate benthic communities: A new case of emerging disease linked to climate change. Glob. Chang. Biol. 2007, 13, 2078–2088. [Google Scholar] [CrossRef]
- Astraldi, M.; Gasparini, G.P.; Manzella, G.M.R.; Hopkins, T.S. Temporal variability of currents in the eastern Ligurian Sea. J. Geophys. Res. Space Phys. 1990, 95, 1515–1522. [Google Scholar] [CrossRef]
- Connell, S.; Foster, M.; Airoldi, L. What are algal turfs? Towards a better description of turfs. Mar. Ecol. Prog. Ser. 2014, 495, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Bramanti, L.; Vielmini, I.; Rossi, S.; Tsounis, G.; Iannelli, M.; Cattaneo-Vietti, R.; Priori, C.; Santangelo, G. Demographic parameters of two populations of red coral (Corallium rubrum L. 1758) in the North Western Mediterranean. Mar. Biol. 2014, 161, 1015–1026. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Levene, H. Robust Tests for Equality of Variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling; Olkin, I., Ed.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Siegel, S. Nonparametric Statistics for the Behavioral Sciences; McGraw-Hill: New York, NY, USA, 1956. [Google Scholar]
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Caswell, H. Matrix Population Models; Sinauer Associates: Sunderland, MA, USA, 2001; p. 722. [Google Scholar]
- Cupido, R. Demography of the Red Gorgonian Paramuricea clavata (Anthozoa, Octocorallia) Damaged by Repeated Anomalous Mortality Events. Ph.D. Thesis, University of Pisa, Pisa, Italy, 2010. [Google Scholar]
- Santangelo, G.; Fronzoni, L. Global Climate Change and the Ecology of the Next Decade; Edizioni ETS: Pisa, Italy, 2008. [Google Scholar]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.N.; Steneck, R.S.; Mumby, P.J. Running the gauntlet: Inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 2010, 414, 91–105. [Google Scholar] [CrossRef]
- Linares, C.; Cebrian, E.; Coma, R. Effects of turf algae on recruitment and juvenile survival of gorgonian corals. Mar. Ecol. Prog. Ser. 2012, 452, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Pilczynska, J.; Cocito, S.; Boavida, J.; Serrão, E.; Queiroga, H. Genetic Diversity and Local Connectivity in the Mediterranean Red Gorgonian Coral after Mass Mortality Events. PLoS ONE 2016, 11, e0150590. [Google Scholar] [CrossRef] [Green Version]
- Coma, R.; Gili, J.M.; Zabala, M.; Riera, T. Feeding and prey capture cycles in the aposymbiontic gorgonian Paramuricea clavata. Mar. Ecol. Prog. Ser. 1994, 115, 257–270. [Google Scholar] [CrossRef]
- Linares, C.; Coma, R.; Garrabou, J.; Díaz, D.; Zabala, M. Size distribution, density and disturbance in two Mediterranean gorgonians: Paramuricea clavata and Eunicella singularis. J. Appl. Ecol. 2008, 45, 688–699. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2013 | 1998 | ||||
|---|---|---|---|---|---|
| Tinetto Islet Area | Tinetto Island | Tinetto Shoal | Tinetto Islet Area | ||
| n = 23 | n = 12 | n = 11 | n = 24 | ||
| % Cover ± ES | TSH | 65.1 ± 1.4 | 65 ± 2.3 | 65.3 ± 1.7 | 39 ± 1 |
| Undetermined area | 1.3 ± 1.4 | 1.5 ± 0.2 | 1 ± 0.2 | 0 | |
| Occupied substrate | 33.6 ± 1.5 | 33.5 ± 2.5 | 33.7 ± 1.8 | 61 ± 1 | |
| Porifera | 9.3 ± 0.7 | 10.9 ± 0.6 * | 7.6 ± 1.2 * | - | |
| Hexacorallia | 4 ± 0.4 | 5.8 ± 1 * | 2 ± 0.2 * | - | |
| Algae | 2 ± 0.04 | 1.1 ± 0.1 * | 2.9 ± 0.6 * | - | |
| Ascidiacea, Bryozoa, and Polychetae | 0.29 ± 0.06 | 0.4 ± 0.1 * | 0.2 ± 0.03 * | - | |
| Paramuricea clavata | 16.8 ± 1.7 | 14.0 ± 2.3 * | 19.9 ± 2.4 * | 52.0 ± 1 | |
| Colony density (col m−2) ± ES | Recruits | 3.75 ± 1.74 | 1.33 ± 0.48 * | 6.18 ± 2.14 * | 0.83 ± 0.27 |
| Adults | 17.10 ± 3.01 | 12.07 ± 1.81* | 21.30 ± 4.17* | 35.67 ± 3.20 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruffaldi Santori, S.; Benedetti, M.C.; Cocito, S.; Peirano, A.; Cupido, R.; Erra, F.; Santangelo, G. After the Fall: The Demographic Destiny of a Gorgonian Population Stricken by Catastrophic Mortality. Oceans 2021, 2, 337-350. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2020020
Ruffaldi Santori S, Benedetti MC, Cocito S, Peirano A, Cupido R, Erra F, Santangelo G. After the Fall: The Demographic Destiny of a Gorgonian Population Stricken by Catastrophic Mortality. Oceans. 2021; 2(2):337-350. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2020020
Chicago/Turabian StyleRuffaldi Santori, Simona, Maria Carla Benedetti, Silvia Cocito, Andrea Peirano, Roberta Cupido, Fabrizio Erra, and Giovanni Santangelo. 2021. "After the Fall: The Demographic Destiny of a Gorgonian Population Stricken by Catastrophic Mortality" Oceans 2, no. 2: 337-350. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2020020