
Reef Structural Complexity Influences Fish Community Metrics on a Remote Oceanic Island: Serranilla Island, Seaflower Biosphere Reserve, Colombia


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
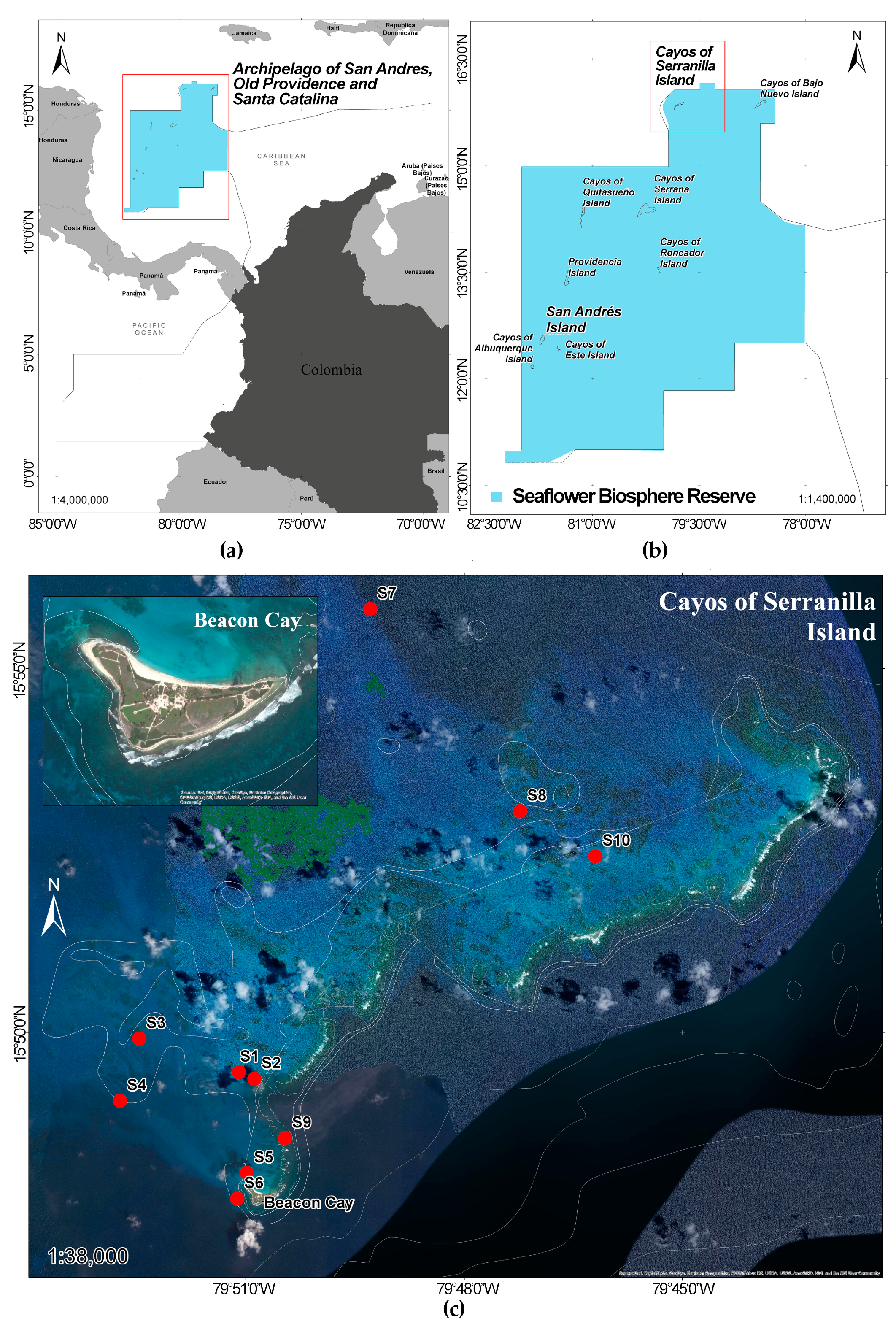
2.1. Study Site
2.2. Data Collection
2.3. Analysis
2.3.1. Relationship of Reef Fish Assemblages to Habitat Structure
2.3.2. Differences in Parrotfish Assemblages between Islands (Serranilla and San Andrés)
3. Results
3.1. Relationship of Reef Fish Assemblages to Habitat Structure
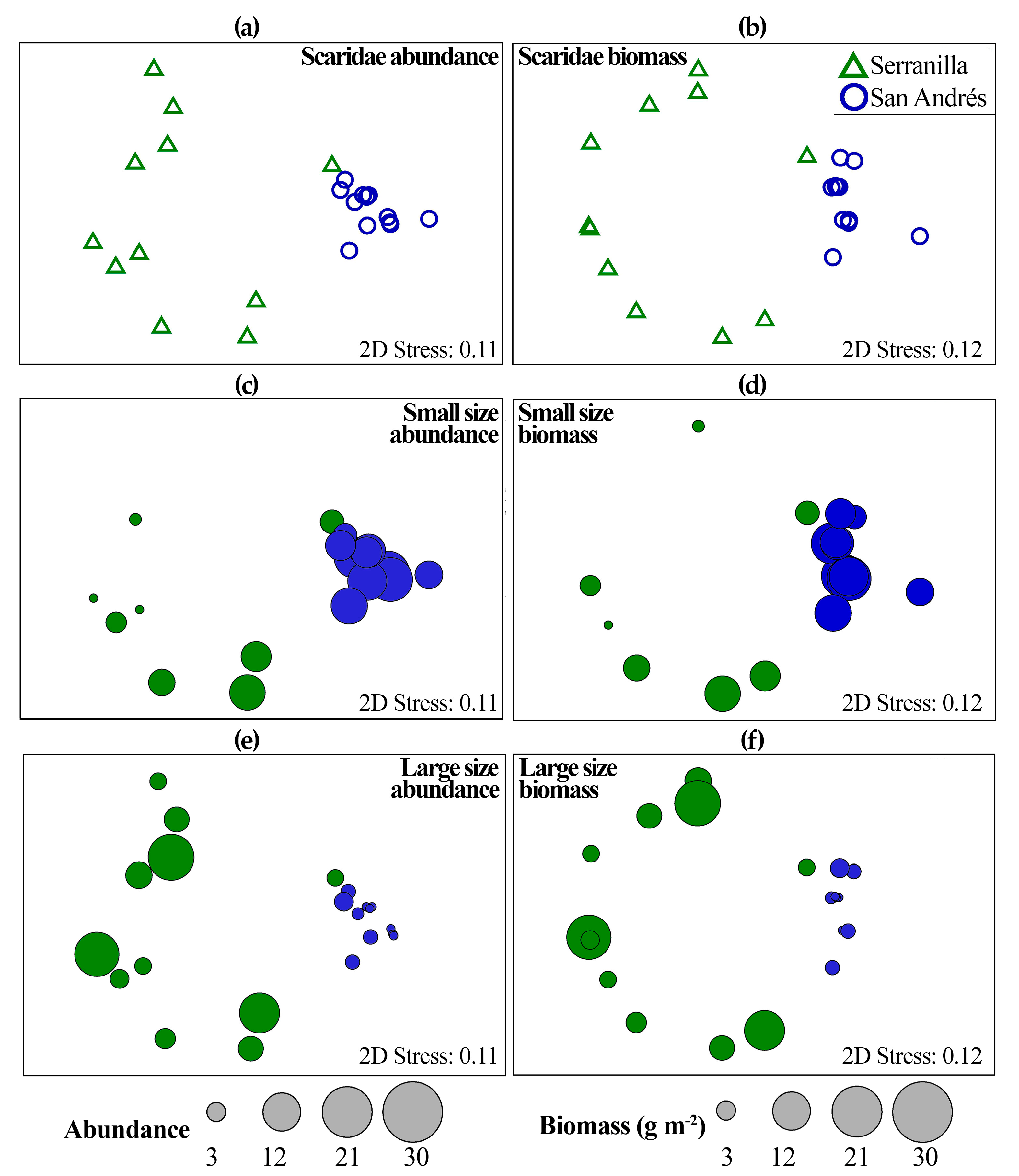
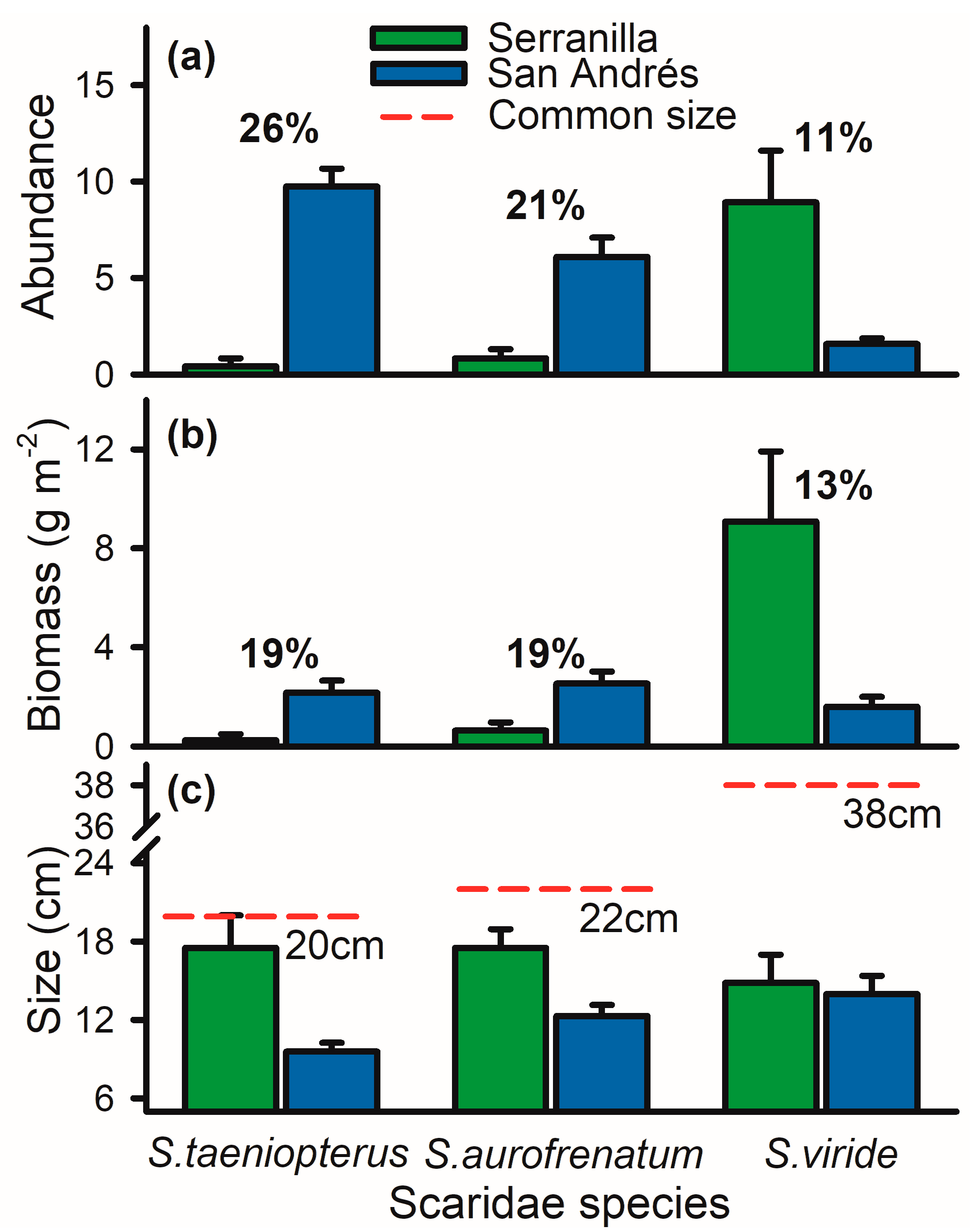
3.2. Parrotfish Differences between Islands (Serranilla and San Andrés)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moritz, C.; Vii, J.; Lee Long, W.; Tamelander, J.; Thomassin, A.; Planes, S. Status and Trends of Coral Reefs of the Pacific. In Global Coral Reef Monitoring Network; Technical Report; United Nations Environment Programme: Washington, DC, USA, 2018; pp. 47–50. [Google Scholar]
- Komyakova, V.; Munday, P.L.; Jones, G.P. Relative Importance of Coral Cover, Habitat Complexity and Diversity in Determining the Structure of Reef Fish Communities. PLoS ONE 2013, 8, e83178. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral reef biodiversity. In Life in the World’s Oceans: Diversity Distribution and Abundance, 1st ed.; McIntyre; Blackwell Publishing Ltd.: West Sussex, UK, 2010; pp. 65–78. [Google Scholar]
- Plaisance, L.; Caley, M.J.; Brainard, R.E.; Knowlton, N. The diversity of coral reefs: What are we missing? PLoS ONE 2011, 6, e25026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhead, A.J.; Hicks, C.C.; Norström, A.V.; Williams, G.J.; Graham, N.A.J. Coral reef ecosystem services in the Anthropocene. Funct. Ecol. 2019, 33, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Costanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the Global Value of Ecosystem Services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Waite, R.; Burke, L.; Gray, E. Coastal Capital: Ecosystem Valuation for Decision Making in the Caribbean; World Resources Institute: Washington, DC, USA, 2014; pp. 1–78. [Google Scholar]
- Elliff, C.I.; Silva, I.R. Coral Reefs as the First Line of Defense: Shoreline Protection in Face of Climate Change. Mar. Environ. Res. 2017, 127, 148–154. [Google Scholar] [CrossRef]
- Reguero, B.G.; Beck, M.W.; Agostini, V.N.; Kramer, P.; Hancock, B. Coral Reefs for Coastal Protection: A New Methodological Approach and Engineering Case Study in Grenada. J. Environ. Manag. 2018, 210, 146–161. [Google Scholar] [CrossRef]
- Samonte-Tan, G.P.B.; White, A.T.; Tercero, M.A.; Diviva, J.; Tabara, E.; Caballes, C. Economic Valuation of Coastal and Marine Resources: Bohol Marine Triangle, Philippines. Coast. Manag. 2007, 35, 319–338. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.; Spalding, M.D.; Perry, A. Reefs at Risk Revisited; World Resources Institute: Washington, DC, USA, 2011; p. 114. [Google Scholar]
- Pandolfi, J.M.; Connolly, S.R.; Marshall, D.J.; Cohen, A.L. Projecting Coral Reef Futures under Global Warming and Ocean Acidification. Science 2011, 333, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Long-Term Region-Wide Declines in Caribbean Corals. Science 2003, 301, 958–960. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.; Maidens, J. Reefs at Risk in the Caribbean; World Resources Institute: Washington, DC, USA, 2004; p. 80. [Google Scholar]
- Mumby, P.J.; Wolff, N.H.; Bozec, Y.M.; Chollett, I.; Halloran, P. Operationalizing the Resilience of Coral Reefs in an Era of Climate Change. Conserv. Lett. 2014, 7, 176–187. [Google Scholar] [CrossRef]
- Álvarez-Filip, L.; Dulvy, N.K.; Gill, J.A.; Côté, I.M.; Watkinson, A.R. Flattening of Caribbean Coral Reefs: Region-Wide Declines in Architectural Complexity. Proc. Royal Soc. B 2009, 276, 3019–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Filip, L.; Carricart-Ganivet, J.P.; Horta-Puga, G.; Iglesias-Prieto, R. Shifts in Coral-Assemblage Composition Do Not Ensure Persistence of Reef Functionality. Sci. Rep. 2013, 3, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zea, S.; Geister, J.; Garzón-Ferreira, J.; Díaz, J.M. Biotic Changes in the Reef Complex of San Andres Island (Southeastern Caribbean Sea, Columbia) Occuring over Three Decades. Atoll. Res. Bull. 1998, 456, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Guannel, G.; Arkema, K.; Ruggiero, P.; Verutes, G. The Power of Three: Coral Reefs, Seagrasses and Mangroves Protect Coastal Regions and Increase Their Resilience. PLoS ONE 2016, 11, e0158094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendleton, L.H.; Hoegh-Guldberg, O.; Langdon, C.; Comte, A. Multiple Stressors and Ecological Complexity Require a New Approach to Coral Reef Research. Front. Mar. Sci. 2016, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Mumby, P.J. The impact of exploiting grazers (Scaridae) on the dynamics of Caribbean coral reefs. Ecol. Appl. 2006, 16, 747–769. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.; Donovan, M.; Cramer, K.; Lam, V. Status and Trends of Caribbean Coral Reefs; Global Coral Reef Monitoring Network: Washington, DC, USA, 2014; p. 304. [Google Scholar]
- Jackson, J.B.C. Reefs since Columbus. Coral Reefs 1997, 16, S23–S32. [Google Scholar] [CrossRef]
- Carpenter, R.C. Mass Mortality of Diadema Antillarum II. Effects on Population Densities and Grazing Intensity of Parrotfishes and Surgeonfishes. Mar. Biol. 1990, 104, 79–86. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Hay, M.E. Herbivore Species Richness and Feeding Complementarity Affect Community Structure and Function on a Coral Reef. Proc. Natl. Acad. Sci. USA 2008, 105, 16201–16206. [Google Scholar] [CrossRef] [Green Version]
- Adam, T.C.; Schmitt, R.J.; Holbrook, S.J.; Brooks, A.J.; Edmunds, P.J.; Carpenter, R.C.; Bernardi, G. Herbivory, Connectivity, and Ecosystem Resilience: Response of a Coral Reef to a Large-Scale Perturbation. PLoS ONE 2011, 6, e23717. [Google Scholar] [CrossRef] [PubMed]
- Mumby, P.J.; Dahlgren, C.P.; Harborne, A.R.; Kappel, C.V.; Micheli, F.; Brumbaugh, D.R.; Holmes, K.E.; Mendes, J.M.; Broad, K.; Sanchirico, J.N.; et al. Fishing, Trophic Cascades, and the Process of Grazing on Coral Reefs. Science 2006, 311, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Bonaldo, R.; Hoey, A.; Bellwood, D. The Ecosystem Roles of Parrotfishes on Tropical Reefs. Oceanogr. Mar. Biol. Annu. Rev. 2014, 52, 81–132. [Google Scholar]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyströ, M. Confronting the Coral Reef Crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Hawkins, J.P.; Roberts, C.M. Effects of Artisanal Fishing on Caribbean Coral Reefs. Conserv. Biol. 2004, 18, 215–226. [Google Scholar] [CrossRef]
- Bozec, Y.M.; O’Farrell, S.; Bruggemann, J.H.; Luckhurst, B.E.; Mumby, P.J. Tradeoffs between Fisheries Harvest and the Resilience of Coral Reefs. Proc. Natl. Acad. Sci. USA 2016, 113, 4536–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-López, D.I.; Segura-Quintero, C.; Sierra-Correa, P.C.; Garay-Tinoco, J. Atlas de La Reserva de Biósfera Seaflower. Archipiélago de San Andrés, Providencia y Santa Catalina; Serie de Publicaciones Especiales; INVEMAR: Bogotá, Colombia, 2012; p. 180. [Google Scholar]
- Bruckner, A. Global Reef Expedition: San Andrés Archipelago, Colombia; Field Report; Khaled bin Sultan Living Oceans Foundation: Landover, MD, USA, 2012; p. 52. [Google Scholar]
- Samoilys, M.A.; Carlos, G. Determining Methods of Underwater Visual Census for Estimating the Abundance of Coral Reef Fishes. Environ. Biol. Fishes 2000, 57, 289–304. [Google Scholar] [CrossRef]
- World Wild Fund (WWF). Mejores prácticas de pesca en arrecifes coralinos. In Guía para la Colecta de Información que Apoye el Manejo de Pesquerías Basado en Ecosistemas; WWF: Cancún, México; San José, Costa Rica, 2006; p. 81. [Google Scholar]
- Caldwell, Z.R.; Zgliczynski, B.J.; Williams, G.J.; Sandin, S.A. Reef Fish Survey Techniques: Assessing the Potential for Standardizing Methodologies. PLoS ONE 2016, 11, e0153066. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. Fish Base. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 12 December 2019).
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Species by Family/Subfamily. Eschmeyer’s Catalog of Fishes. Available online: www.researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 21 December 2019).
- Risk, M.J. Fish Diversity on a Coral Reef in the Virgin Islands. Atoll. Res. Bull. 1972, 153, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Entropy and Diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice-Hall: New Jersey, NJ, USA, 2010; p. 944. [Google Scholar]
- Anderson, M.; Gorley, R.; Clarke, K.P. For PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v6 Permanova+; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Harborne, A.R.; Mumby, P.J.; Ferrari, R. The Effectiveness of Different Meso-Scale Rugosity Metrics for Predicting Intra-Habitat Variation in Coral-Reef Fish Assemblages. Environ. Biol. Fishes 2012, 94, 431–442. [Google Scholar] [CrossRef]
- Richardson, L.E.; Graham, N.A.J.; Pratchett, M.S.; Hoey, A.S. Structural Complexity Mediates Functional Structure of Reef Fish Assemblages among Coral Habitats. Environ. Biol. Fishes 2017, 100, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Van Zanten, B.T.; van Beukering, P.J.H.; Wagtendonk, A.J. Coastal Protection by Coral Reefs: A Framework for Spatial Assessment and Economic Valuation. Ocean Coast. Manag. 2014, 96, 94–103. [Google Scholar] [CrossRef]
- Monismith, S.G.; Rogers, J.S.; Koweek, D.; Dunbar, R.B. Frictional Wave Dissipation on a Remarkably Rough Reef. Geophys. Res. Lett. 2015, 42, 4063–4071. [Google Scholar] [CrossRef]
- Walker, B.K.; Jordan, L.K.B.; Spieler, R.E. Relationship of Reef Fish Assemblages and Topographic Complexity on Southeastern Florida Coral Reef Habitats. J. Coast. Res. 2009, 53, 39–48. [Google Scholar] [CrossRef]
- Rogers, A.; Blanchard, J.L.; Mumby, P.J. Vulnerability of Coral Reef Fisheries to a Loss of Structural Complexity. Curr. Biol. 2014, 24, 1000–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedlander, A.M.; Parrish, J.D. Habitat Characteristics Affecting Fish Assemblages on a Hawaiian Coral Reef. J. Exp. Mar. Biol. Ecol. 1998, 224, 1–30. [Google Scholar] [CrossRef]
- González-Rivero, M.; Harborne, A.R.; Herrera-Reveles, A.; Bozec, Y.M.; Rogers, A.; Friedman, A.; Ganase, A.; Hoegh-Guldberg, O. Linking Fishes to Multiple Metrics of Coral Reef Structural Complexity Using Three-Dimensional Technology. Sci. Rep. 2017, 7, 1–15. [Google Scholar]
- Kuffner, I.B.; Brock, J.C.; Grober-Dunsmore, R.; Bonito, V.E.; Hickey, T.D.; Wright, C.W. Relationships between Reef Fish Communities and Remotely Sensed Rugosity Measurements in Biscayne National Park, Florida, USA. Environ. Biol. Fishes 2007, 78, 71–82. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L. The Importance of Structural Complexity in Coral Reef Ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; Ferse, S.C.A.; Jompa, J.; Wild, C.; Teichberg, M. Fish Herbivory as Key Ecological Function in a Heavily Degraded Coral Reef System. Limnol. Oceanogr. 2015, 60, 1382–1391. [Google Scholar] [CrossRef]
- Paddack, M.J.; Reynolds, J.D.; Aguilar, C.; Appeldoorn, R.S.; Beets, J.; Burkett, E.W.; Chittaro, P.M.; Clarke, K.; Esteves, R.; Fonseca, A.C.; et al. Dynamic Fragility of Oceanic Coral Reef Ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 8425–8429. [Google Scholar]
- Pratchett, M.S.; Munday, P.; Wilson, S.K.; Graham, N.A.; Cinner, J.E.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.; McClanahan, T.R. Effects of Climate-Induced Coral Bleaching on Coral-Reef Fishes. Ecological and economic consequences. Oceanogr. Mar. Bio Annu. Rev. 2008, 46, 251–296. [Google Scholar]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; et al. Global Trajectories of the Long-Term Decline of Coral Reef Ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallès, H.; Gill, D.; Oxenford, H.A. Parrotfish Size as a Useful Indicator of Fishing Effects in a Small Caribbean Island. Coral Reefs 2015, 34, 789–801. [Google Scholar] [CrossRef]
- Cinner, J.E.; Graham, N.A.J.; Huchery, C.; Macneil, M.A. Global Effects of Local Human Population Density and Distance to Markets on the Condition of Coral Reef Fisheries. Conserv. Biol. 2013, 27, 453–458. [Google Scholar] [CrossRef]
- Brewer, T.D.; Cinner, J.E.; Fisher, R.; Green, A.; Wilson, S.K. Market Access, Population Density, and Socioeconomic Development Explain Diversity and Functional Group Biomass of Coral Reef Fish Assemblages. Glob. Environ. Chang. 2012, 22, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Prato, J.A.; Newball, R. Aproximación a La Valoración Económica Ambiental Del Departamento Archipiélago de San Andrés, Providencia y Santa Catalina; Comisión Colombiana del Océano: Bogotá, Colombia, 2016.
- Chasqui, V.; Polanco, L.A.; Acero, A.; Mejía-Falla, P.A.; Navia, A.; Zapata, L.A.; Caldas, J.P. Libro Rojo de Peces Marinos de Colombia; Instituto de Investigaciones Marinas y Costeras Invemar, Ministerio de Ambiente y Desarrollo Sostenible: Santa Marta, Colombia, 2017; p. 552.
- Friedlander, A.; Nowlis, J.S.; Sanchez, J.A.; Appeldoorn, R.; Usseglio, P.; Mccormick, C.; Bejarano, S.; Mitchell-Chui, A. Designing Effective Marine Protected Areas in Seaflower Biosphere Reserve, Colombia, Based on Biological and Sociological Information. Conserv. Biol. 2003, 17, 1769–1784. [Google Scholar] [CrossRef]
- McField, M.; Kramer, P.; Giró Petersen, A.; Soto, M.; Drysdale, I.; Craig, N.; Rueda-Flores, M. Mesoamerican Reef Report Card, Healthy Reefs Initiative. Available online: www.healthyreef.org (accessed on 28 June 2021).
- Bruggemann, J.H.; van Rooij, J.M.; Videler, J.J.; Breeman, A.M. Dynamics and Limitations of Herbivore Populations on a Caribbean Coral Reef. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 1995. [Google Scholar]
- Hernández-Landa, R.C.; Aguilar-Perera, A. Structure and composition of surgeonfish (Acanthuridae) and parrotfish (Labridae: Scarinae) assemblages in the south of the Parque Nacional Arrecife Alacranes, southern Gulf of Mexico. Mar. Biol. 2019, 49, 647–662. [Google Scholar] [CrossRef]
- Jaimes Rodríguez, L.I. Algunos Aspectos Biológico-Pesqueros de Las Principales Especies Ícticas Capturadas En El Sector de San Bernardo, Parque Nacional Natural Corales Del Rosario y de San Bernardo, Caribe Colombiano. Bachelor’s Thesis, Universidad de Bogotá Jorge Tadeo Lozano, Bogotá, Colombia, 2011. [Google Scholar]
- McClanahan, T.; Karnauskas, M. Relationships between Benthic Cover, Current Strength, Herbivory, and a Fisheries Closure in Glovers Reef Atoll, Belize. Coral Reefs 2011, 30, 9–19. [Google Scholar] [CrossRef]
- Balzan, M.V.; Caruana, J.; Zammit, A. Assessing the Capacity and Flow of Ecosystem Services in Multifunctional Landscapes: Evidence of a Rural-Urban Gradient in a Mediterranean Small Island State. Land Use Policy 2018, 75, 711–725. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | Num. of Variables | p-Values of Variables in the Model | Total Model | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Rugo | Bran | Sphe | Brain | Subm | Algae | R2adj | p-Value | ||
| Biomass | All | 0.46 | 0.81 | 0.87 | 0.41 | 0.71 | 0.16 | 0.364 | 0.066 |
| 5 | 0.54 | 0.34 | 0.37 | 0.87 | 0.22 | 0.378 | 0.427 | ||
| Abundance | All | 0.001 | 0.94 | 0.49 | 0.59 | 0.09 | 0.51 | 0.558 | 0.01 |
| 3 | 0.14 | 0.56 | 0.41 | 0.385 | 0.016 | ||||
| 0D | All | 0.03 | 0.9 | 0.81 | 0.7 | 0.94 | 0.71 | 0.463 | 0.028 |
| 1 | <0.001 | 0.592 | <0.001 | ||||||
| 1D | All | 0.3 | 0.83 | 0.71 | 0.98 | 0.52 | 0.14 | 0.389 | 0.034 |
| 3 | 0.23 | 0.39 | 0.06 | 0.502 | 0.003 | ||||
| 2D | All | 0.36 | 0.56 | 0.54 | 0.92 | 0.51 | 0.07 | 0.357 | 0.07 |
| 3 | 0.02 | 0.73 | 0.01 | 0.459 | 0.006 | ||||
| 2/1D | All | 0.22 | 0.23 | 0.24 | 0.49 | 0.64 | 0.12 | 0.237 | 0.156 |
| 4 | 0.16 | 0.6 | 0.48 | 0.26 | 0.261 | 0.083 | |||
| Small parrotfish abundance | All | 0.28 | <0.01 | 0.12 | <0.01 | 0.082 | 0.83 | 0.661 | 0.002 |
| 4 | 0.53 | <0.01 | <0.01 | 0.49 | 0.408 | 0.006 | |||
| Large parrotfish abundance | All | 0.02 | 0.15 | 0.11 | 0.77 | 0.23 | 0.61 | 0.376 | 0.06 |
| 2 | <0.01 | 0.22 | 0.408 | 0.006 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castaño, D.; Morales-de-Anda, D.; Prato, J.; Cupul-Magaña, A.L.; Echeverry, J.P.; Santos-Martínez, A. Reef Structural Complexity Influences Fish Community Metrics on a Remote Oceanic Island: Serranilla Island, Seaflower Biosphere Reserve, Colombia. Oceans 2021, 2, 611-623. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030034
Castaño D, Morales-de-Anda D, Prato J, Cupul-Magaña AL, Echeverry JP, Santos-Martínez A. Reef Structural Complexity Influences Fish Community Metrics on a Remote Oceanic Island: Serranilla Island, Seaflower Biosphere Reserve, Colombia. Oceans. 2021; 2(3):611-623. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030034
Chicago/Turabian StyleCastaño, Diana, Diana Morales-de-Anda, Julián Prato, Amílcar Leví Cupul-Magaña, Johanna Paola Echeverry, and Adriana Santos-Martínez. 2021. "Reef Structural Complexity Influences Fish Community Metrics on a Remote Oceanic Island: Serranilla Island, Seaflower Biosphere Reserve, Colombia" Oceans 2, no. 3: 611-623. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030034