
Reef Fish Associations with Natural and Artificial Structures in the Florida Keys
by
 , and
, and
Kara Noonan
1,* ,
,
Thomas Fair
2,
Kristiaan Matthee
2,
Kelsey Sox
2,
Kylie Smith
2 and
Michael Childress
2 1
Department of Ecology and Evolutionary Biology, Rice University, Houston, TX 77005, USA
2
Department of Biological Sciences, Clemson University, Clemson, SC 29634, USA
*
Author to whom correspondence should be addressed.
Oceans 2021, 2(3), 634-647; https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030036
Submission received: 29 October 2020
/
Revised: 22 July 2021
/
Accepted: 18 August 2021
/
Published: 8 September 2021
(This article belongs to the Special Issue The Future of Coral Reefs: Research Submitted to ICRS 2020, Bremen, Germany)
Abstract
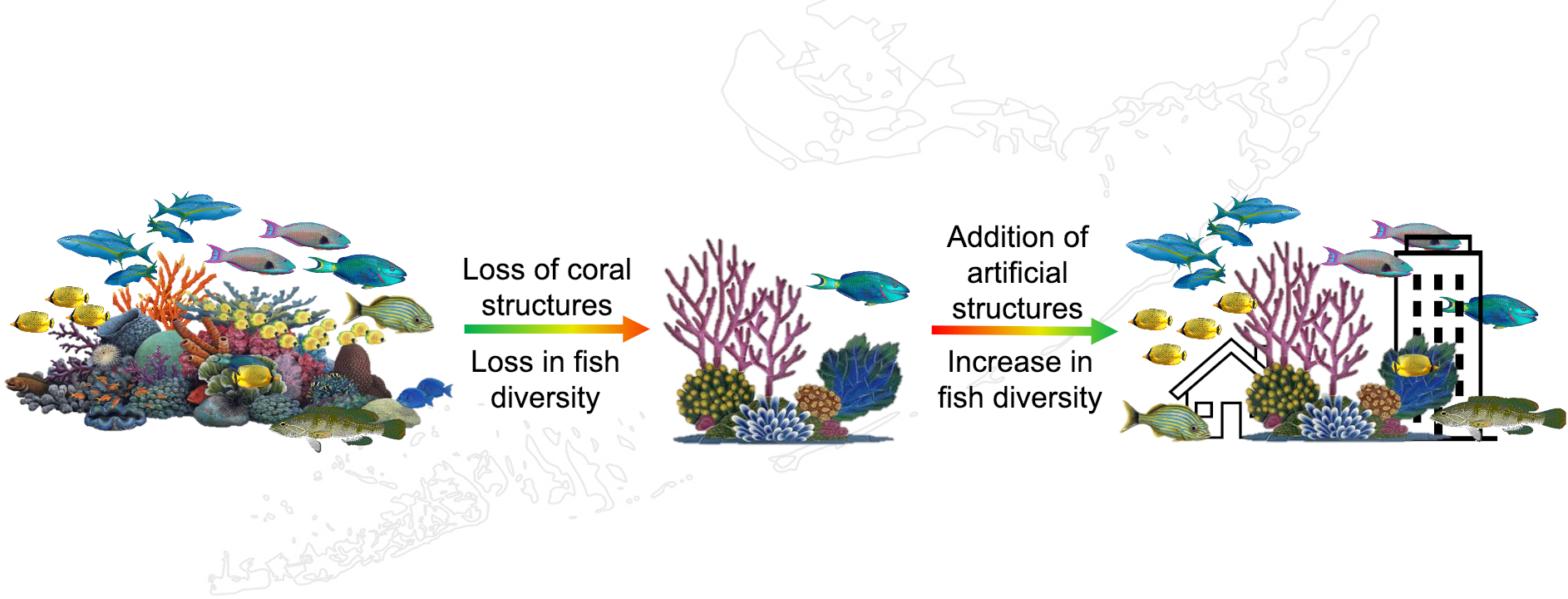
:Throughout the Caribbean, coral reefs are transitioning from rugose, coral-dominated communities to flat, soft coral-dominated habitats, triggering declines in biodiversity. To help mitigate these losses, artificial structures have been used to re-create substrate complexity and support reef inhabitants. This study used natural and artificial structures to investigate the factors influencing the use of habitat by reef fish. During 2018 and 2019, divers added artificial structures and monitored the fish assemblages associating with both the artificial structures and naturally occurring corals. Overall, there were more fish on natural structures than on artificial structures. While structure shape did not influence fish use, there was a non-significant trend for increased use of larger structures. Fish observations did not differ across a gradient of shallow, complex reefs to deeper, flatter reefs; however, analyses of feeding guilds revealed clearer patterns: herbivores and omnivores were positively associated with low rugosity reefs where macroalgal abundance was higher, whereas invertivores preferred more rugose reefs. These results suggest that as reefs lose structural complexity, fish communities may become dominated by herbivores and omnivores. It also appears that the addition of artificial structures of the type used here may not mitigate the effects of structure loss on reef fish assemblages.

1. Introduction
Coral reefs are amongst the most diverse and complex ecosystems in the ocean, despite occupying less than 1% of the ocean floor [1,2]. The combination of soft and hard structures creates an architecturally complex marine habitat that is heavily utilized by numerous organisms and is considered to provide hotspots of diversity and endemism [3,4,5,6]. There are both structural and biotic components that make coral reefs multifaceted environments, including rugosity [7,8,9,10,11,12,13], algae [8,9,14], hard coral morphology [15,16], and emergent limestone ledges [17,18]; all of these have been described to increase reef fish species diversity.
Caribbean coral reefs have been experiencing severe degradation due to continual disturbances including, but not limited to, disease, sedimentation, and eutrophication, which, together, are eliminating the complex landscapes [19,20]. In many areas, Caribbean coral reef degradation far surpasses that of Indo-Pacific coral reefs, so the Caribbean has become the focal area for studies analyzing the response of reef fishes to this rapid loss of reef structural complexity. Long-term studies following species-specific responses to coral decline found that 43 out of the 72 fish species censused had experienced declines greater than 50% [21,22,23,24,25,26,27,28,29]. Other studies have found similar trends, with estimated density losses of 2.7–6.0% per year [30], and with predictive models estimating continued losses for particular functional feeding guilds such as invertivores in the years to come [31].
To combat the loss of complex coral structures, artificial reef structures (ARs) have been used to increase the physical complexity and substrate available to support reef fish communities. There have been many studies that indicated a positive effect of ARs, but these have identified that particular characteristics are necessary for this mitigation strategy to be effective [32,33,34,35,36,37,38,39]. The overall height of ARs has a significant impact on their effectiveness [36,40,41], while the size, surface area available, and complexity of ARs appear to influence the diversity of reef fish across an entire reef [40,41,42]. In fact, some artificial reefs have been observed to contain species assemblages that are more diverse than those of natural reefs, leading to the conclusion that this method of intervention can be successful [36,37,43,44,45].
Previous studies have assessed reef fish community responses to coral decline [6,24,27,29] and the use of ARs [32,33,36,37,39], and predicted how reef fish communities may be structured in the future [31], but there is little research investigating how reef fish are utilizing the structures that remain in the Caribbean. The goal of the present study was to investigate how reef fishes utilize both natural and artificial structures, identify structural characteristics that may influence their use, and assess whether reef location and topographic complexity influence the use of structure by different functional feeding guilds. Based on previous literature, we investigated the hypotheses that reef fishes would utilize biotically complex natural structures more often than non-biotic artificial structures, that height and surface area would be the most effective characteristics driving use, and that reef fishes’ use of structures would be evident on reefs with higher rugosity but would differ between different feeding guilds. By identifying heavily utilized structures and their associated traits, we can better predict the reef fish community response to structural declines and assess whether artificial structures can mitigate further losses.
2. Materials and Methods
2.1. Site Selection and Substrate Survey
Field surveys were conducted in the summers of 2018 and 2019 across 8 reef sites, which varied in their distance from shore (1.62 to 8.86 km) and depth (3.0 to 8.1 m), in the middle of Florida Keys National Marine Sanctuary (Figure 1). Depths were calculated during the mid-tide transition using a depth finder on a boat when it was positioned over the middle of the reef. Each reef area was surveyed using a permanent 50 m transect that ran parallel to the primary axis of the reef, and 4 30 m transects that were laid perpendicular to and crossing the permanent transect at distances of 10, 20, 30, and 40 m, creating a 50 × 30 m grid. The substrate cover of the study area was recorded using digital photographs of 50 × 50 cm portions of the substrate, starting with 2 pictures on each side of the permanent 50 m transect at 0 m, 2 pictures again at 10 m, at 20 m, at 30 m, at 40 m, and again at 50 m (i.e., a total of 12 images per reef = 96 images for all reefs). Percent substrate cover by the major substrates (hard corals, soft corals, sponges, fleshy algae, turf algae, calcareous algae, sand) was estimated using 25 randomly selected points per photograph, with the aid of Coral Point Count with the Excel extension [46]. Rugosity was measured 3 times on each of the 4 30 m perpendicular transects using a chain and tape method. A 2 m chain was placed along the side of the 30 m transect and the beginning and end chain measurements were recorded and subtracted from each other. If a reef was flat, the chain would extend a full 2 m on the transect tape, giving an overall measurement of low rugosity. If a reef was more complex, the chain would not extend the full 2 m. The rugosity measurements for each site (3 per 30 m transect × 4 per 30 m transects = 12 rugosity measurements) were then averaged to obtain an overall rugosity across the site.
2.2. Video Transect Surveys as an Assessment of Reef Fish Abundance
Some reefs are known to have more abundant and diverse reef fish communities, and we suspected that this may affect how structures are used across a reef tract. We therefore began by establishing a baseline of abundance across the 8 sites. To evaluate the overall abundance and diversity of reef fish species within the functional feeding guilds present at the 8 sites, 8 videos (~3–4 min) were captured along each of the perpendicular 30 m transects (2 videos per transect) between the hours of 900 and 1500. A diver swam the length of the transect while holding a PVC camera frame with 2 forward-facing GoPro cameras attached at heights of 30 cm and 100 cm above the substrate, so as to capture both benthic and mid-water fish species. Each video was analyzed to identify both the fish species present and any behavioral interactions with the substrate including, but not limited to, using substrate for shelter or as a food resource. Fish species were classified into one of four functional feeding guilds (Table S1): herbivores, omnivores, invertivores, and piscivores [25,29,47]. Analysis of fish feeding guilds has been found to be adequate for comparing reef fish community structures and can distinguish functionally diverse communities from seemingly diverse communities that have functional redundancy [48,49,50]. Although fish of the family Haemulidae are omnivores, they were separated into their own functional feeding guild—invertivores—because of their nocturnal feeding activities and daytime use of the structure for rest.
2.3. Natural and Artificial Structures
On each of the 8 reef sites used for this study, 1 soft coral sea rod, and 2 healthy, 2 diseased, and 2 recently dead hard coral colonies were selected as the natural structures for further monitoring (Figure 2). Thus, a total of 8 soft coral sea rods, 16 healthy, 16 diseased, and 16 dead hard coral colonies were tracked across our 8 reef sites. The hard coral colonies included five common boulder coral species (Colpophyllia natans, Montastraea cavernosa, Orbicella faveolata, Porites astreoides, and Siderastrea siderea), whereas the soft coral was a species from the family Plexauridae. “Sea rod”, “dead”, “diseased”, and “healthy” were used as terms to refer to these different types of natural structure. In 2018, each of these corals was tagged, photographed, and analyzed using ImageJ software to estimate the percent of live, diseased, and dead coral tissue. Estimated percent cover of dead, diseased, or live tissue was converted into surface area using the surface area formula of a half-dome (2 πr2), with the radius estimated as half the mean of the height, length, and width of the coral. In 2019, the resulting 56 corals were re-photographed and re-analyzed to evaluate any changes in tissue cover and surface area. Of the 16 diseased hard corals infected with Stony Coral Tissue Loss Disease (SCTLD) in 2018, 15 had survived and were classified as “healed coral colonies” for the purpose of the 2019 survey.
Separately from the survey of the existing corals, at each reef site, 4 artificial reef structures (ARs) were deployed, 1 of each of 4 different types, variously created out of PVC piping, concrete, rope, tomato cages, and Vexar mesh (plastic coated wire grid) (Figure 2). The 4 structures were designed to imitate different types of natural habitats. The first type consisted of just 1 single length of PVC pipe standing up from a concrete block; this was a structure that lacked physical complexity. The other artificial structures represented a soft coral (created with PVC arms and frayed rope), a boulder coral (created with Vexar mesh around a tomato cage), and a branching elkhorn coral (created with arms wrapped in Vexar extending out from the center) (Figure 2). These different types of artificial structure were referred to as “control”, “soft”, “boulder”, and “elkhorn” respectively. These 4 artificial structures were designed to assess which characteristic (holes, surface area, edge space), if any, influenced use by the reef fish community. These artificial structures were similar in height (100 cm) and diameter (20–100 cm) to the natural boulder hard corals (height 25–200 cm and diameter 30–150 cm) and natural sea rod soft corals (height 30–125 cm and diameter 30–100 cm) present on the reef. In 2018, the artificial structures were deployed, left to acclimate for 2 weeks, surveyed, then removed from the site. In 2019, the artificial structures were redeployed for 2 weeks and surveyed again before being removed. The 2-week deployment and removing schedule was set to minimize the amount of biotic build up on the artificial structures as well as to avoid the potential for reef damage from breakage during the hurricane season.
2.4. Reef Fish Observations on Individual Structures
Reef fishes’ use of each individual structure was estimated using time-lapsed video-photography. A single, anchored GoPro was placed facing the structure and set to take a picture every minute for 60 min, the maximum time possible, given the power of the batteries. The camera was mounted 50 cm above the substrate and 150 cm away from the center of the structure. Every photograph after the first 5 min except the last 5 min was analyzed for fish species and for the relation of the fish to the structure (near or far). Only the photos captured after the first 5 min and before the final 5 min were analyzed to avoid any diver influence on reef fishes’ use of the structure. Juvenile and adult fishes were treated the same for this study. Only those fish near the camera and directly above, below, beside, or in front of the structure were considered to be associated with it. These observed fish were then assigned to the 4 functional feeding guilds referred to above (Table S1). Since observations could be influenced by multiple images of the same individual, we considered the individual structure surveys as a measure of coral use rather than an estimate of fish abundance.
Over the course of 2 summers, we visited the 8 reefs once per year, recording 64 artificial structures and 112 natural structures so as to obtain 10,496 photos, resulting in 29,279 fish observations. After eliminating those fish considered not to be directly in the vicinity of the structure, our dataset consisted of 18,881 counts of 109 species of fish from 31 families, divided among the 4 functional feeding guilds as follows: 6047 individual herbivores from 23 species, 6225 individual omnivores from 48 species, 5725 individual invertivores from 12 species, and 884 individual piscivores from 26 species (Table S1).
2.5. Statistical Analyses
We performed a mixed-model ANOVA for all reef fishes and each functional group of fish against structure type as the main effect, with site as a random factor. Since all models indicated a significant effect of site on reef fish counts, we performed a principal component analysis to create orthogonal component scores that characterized the sites by their physical distance from shore, depth, rugosity, algal, soft coral, hard coral, and sponge substrate covers. The three most significant component scores from this analysis accounted for 72.8% of the variation in physical traits (PC 1 = 35.1%, PC 2 = 27.0%, PC 3 = 10.7%) and were included in the ANCOVA as covariates. We then analyzed fishes’ use of the structures, using a nested analysis of covariance with structure state (artificial or natural) and structure type (control, soft, boulder, elkhorn, sea rod, diseased, dead, or healthy) nested within structure state as the fixed effects, and fish abundance from the video transect surveys and the three component scores as covariates. The first principal component score was the only covariate positively correlated with either of the main effects, so we included an interaction term of PC1 and structure state in our ANCOVA model. We used Tukey’s post-hoc comparisons to estimate differences within structure states.
Observations at each structure (n = 177) were natural log-transformed to meet the assumptions of normality of the residuals and homogeneity of the variances. ANCOVAs were performed separately on the log-transformed counts for all reef fishes together and on the log-transformed counts of each fish feeding guild considered separately (herbivores, omnivores, invertivores, and piscivores). We also conducted ANCOVAs on the five most abundant fish species: bicolor damselfish (Stegastes partitus), striped parrotfish (Scarus iseri), white grunts (Haemulon plumierii), blue-striped grunts (Haemulon sciurus), and schoolmaster snapper (Lutjanus apodus). Linear regressions were used to relate fish counts to measures of hard coral height and to percent cover of live and diseased tissues, as estimated using JMP Pro 14.1.0 software.
3. Results
The reef fish abundances (video transect survey) on the reef sites varied significantly by reef. The eight reef sites differed in rugosity (0.046 to 0.410), hard coral cover (0.0 to 21.4%), soft coral cover (0.0 to 27.7%), sponge cover (1.0 to 15.5%), fleshy algae cover (0.0 to 86.4%), calcareous algae cover (0.0 to 13.8%), turf algae cover (8.4 to 63.4%) and sand cover (0.0 to 42.7%). The differences in abundance (video transect survey) per site accounted for 76.9% of the variance in the use of natural structures (structure survey) and 78% of the variance in the use of artificial structures (structure survey) (Table S2). There was no significant effect of year on the use of the structures by the reef fish on either the natural (F = −1.4376, df = 1.279, p = 0.1517) or artificial structures (F = −1.5778, df = 1.127, p = 0.1171). The total number of reef fishes using the natural structures was significantly more than the total number of reef fishes using the artificial structures (F = 5.8812, df = 1.166, p = 0.0164) (Figure 3). There was, however, no statistically significant preference for a single type of structure, although diseased colonies had the most individuals associated with them and the soft artificial structures had the least. There were significantly more omnivore (F = 5.6278, df = 1.155, p = 0.0189) and invertivore individuals (F = 6.5732, df = 1.123, p = 0.0116) preferring natural structures over artificial structures; they preferred natural sea rod structures the most, although not significantly so. Herbivore use of the structures was not significantly influenced by the overall state of the structure (artificial versus natural) (F = 1.7337, df = 1.162, p = 0.1898), but utilized particular structure types significantly more than others, using diseased coral heads significantly more than any other structure type, and used natural sea rod structures the least. There was no significant effect of structure state or type on the number of piscivorous fishes (F = 0.2082, df = 1.115, p = 0.6491).
The data for the fish species that were most prevalent in the functional feeding guilds were analyzed to determine the reef characteristics that appeared to influence their use of habitats. Among the herbivores, the presence of bicolor damselfish (Stegastes partitus) was strongly correlated with low-complexity reefs (offshore) (F = 11.3251, df = 1.92, p = 0.0011). Striped parrotfish (Scarus iseri) were observed significantly more around natural and artificial structures when their abundance on the reef (video transect survey), as observed in the video transect surveys, was high (F = 8.2186, df = 1.84, p = 0.0052). This was also the pattern observed with the two most frequent invertivore species: blue-striped grunts (Haemulon sciurus) (F = 6.8165, df = 1.57, p = 0.0115) and white grunts (Haemulon plumierii) (F = 14.0461, df = 1.85, p = 0.0003). The most common piscivore was the schoolmaster snapper (Lutjanus apodus), which was significantly more numerous at natural structures (F = 6.3776, df = 1.27, p = 0.0117) than at artificial structures. The omnivores were mostly represented by bluehead wrasses (Thalassoma bifasciatum), which were also more numerous at natural structures than at artificial structures (F = 16.5063, df = 1.129, p < 0.0001) and at the low-complexity offshore sites (F = 4.6595, df = 1.129, p = 0.0327).
The principal component analysis identified three axes which best summarized reef character and substrate composition (Table 1). The first component score (PC1) accounted for 35.1% of the variation and loaded positively with both distance from the shore and depth, but negatively with physical rugosity. The axis characterized reef structural differences between nearshore and offshore reefs in the middle Keys [51,52,53]. The second component score (PC2) accounted for 27.0% of the variation and loaded positively with calcareous algae/sand cover and negatively with fleshy/turf algae cover. The third component score (PC3) accounted for 10.7% of the variance and loaded positively with sponge/hard coral cover and negatively with soft coral cover. The three component scores were used to evaluate whether fishes’ use of structure varied with differences in the physical structure, algal substrates, or hard/soft coral substrate.
We included four covariates in the ANCOVA model for fish use on individual structures including fish abundance (as estimated by video transect surveys), PC1 topography, PC2 algal cover, and PC3 coral cover (Figure 4). The fully fitted ANCOVA for all reef fishes combined was significant (F = 3.04, df = 12,164, p = 0.0007) with an adjusted r2 = 0.122 (Table 2). Four factors significantly contributed to the model: reef fish abundance on the video transects (F = 5.62, df = 1, p = 0.0189, estimate = 0.531), structure state (whether artificial or natural) (F = 10.24, df = 1, p = 0.0015, estimate (artificial) = −0.3601), topography (PC1) (F = 7.182, df = 1, p = 0.0058, estimate = 0.2793), and the structure type by rugosity interaction (F = 6.495, df = 1, p = 0.0117, estimate = 0.2427).
Nested ANCOVAs for each separate feeding guild of fish revealed differences in the use of the structures among fish guilds. For herbivores, the fully fitted ANCOVA was significant (F = 4.55, df = 11,162, p < 0.0001) with an adjusted r2 = 0.184 (Table 2). Three factors significantly contributed to the model: herbivore abundance in the video transect surveys (F = 8.41, df = 1, p = 0.0042, estimate = 0.5265), topography (F = 17.25, df = 1, p < 0.0001, estimate = 0.2217), and the type of natural structure (F = 2.52, df = 1, p = 0.0232) (Table 2). Herbivore use of the structures demonstrated a strong negative relationship with reef rugosity (or a positive relationship with deeper, offshore reefs) but no relationship with algal cover (Figure 4). For the omnivores, there were two factors that significantly contributed to the model of structure use: depth/rugosity (F = 28.69, df = 1, p < 0.0001, estimate = 0.3757) and structure state (F = 5.62, df = 1, p = 0.0189, estimate (artificial) = −0.3195) (Table 2). Omnivores demonstrated a strong negative relationship with reef rugosity (or a positive relationship with deeper, offshore reefs) (Figure 4). For invertivores, three factors significantly contributed to the model of structure use: invertivore abundance observed in the video transect surveys (F = 8.16, df = 1, p = 0.0050, estimate = 0.3742), topography (F = 11.78, df =1, p = 0.0008, estimate = −0.3693), and structure state (artificial or natural) (F = 6.57, df = 1, p = 0.0116, estimate (artificial) = −0.4964) (Table 2). In contrast to the findings for herbivores and omnivores, invertivores’ use of the structures demonstrated a strong positive relationship with reef rugosity (or a negative relationship with deeper, offshore reefs) (Figure 4). The abundance of piscivores across the video transect surveys was the only factor that significantly contributed to the model of structure use (F = 5.88, df = 1, p = 0.0170, estimate = 0.2663) (Table 2 and Figure 4).
4. Discussion
We found that there were no significant effects of rugosity, hard coral cover, soft coral cover, or algal cover on total reef fish and functional feeding guild abundance across our sites. This is different from previous studies that found a greater abundance and diversity of fishes on reefs that have higher hard coral cover and structural complexity [29]. There are several factors that could explain this unexpected finding. First, some studies have found that reef fish abundance generally increases with depth [54]. Second, the shallow-water nearshore habitats of the middle Florida Keys experience a great deal of disturbance, such as nutrient loading or extreme temperatures [55,56], so that there is a possibility of a delayed response of the reef fish community to decades of continuous perturbations [57]. Alternatively, it may be that our methods were not sensitive enough to detect an effect; for example, the acclimation period may not have been long enough, or the artificial structure not large enough to influence fish distribution.
We found that the natural structures were used by fish significantly more than the artificial structures, even though these were designed to mimic the size and shape of the available natural structures. Previous studies have also found that artificial reef structures (ARs) built with numerous distinct holes and crevices were unsuccessful overall [58]. Such studies have also suggested that artificial structures are used mostly at night, a possibility that we did not investigate with our artificial structures [59]. Evidence has also suggested that fish are attracted to the auditory and chemical cues of living hard corals, so the lack of cues from the artificial structures might explain the low rates of use [60,61].
Unsurprisingly, the abundance of fishes on each reef (video transect survey) accounted for more than half of the variance found in the structures’ associations. There were more fish using the structures when there were more fish available on the reef. Generally, reef fish used natural structures significantly more than artificial ones, but, surprisingly, the types of natural (sea rod, dead, diseased, and healthy hard corals) or artificial (control, soft, boulder, and elkhorn) structures did not influence fishes’ use. Natural structures, on average, had similar heights and total surface area to our artificial structures, which suggests that differences in observations of use were not strictly due to space competition. However, the natural structures were covered with living organisms and algae and provided more food than our artificial structures. The small amount of biota could explain why the artificial structures were used significantly less than natural structures [9,14]. Moreover, these differences in the resources available on artificial and natural structures could affect reef fish habitat specialists differently from habitat generalists (Table S2) [29]. We found that artificial structures were occupied more on low-rugosity sites than on high-rugosity sites. This supports previous observations that artificial structures have their greatest impact in environments with fewer structures [62,63].
Further characterization of structure use by different functional feeding guilds revealed other important patterns not apparent from the analysis of use by all reef fishes combined. Herbivore observations on natural and artificial structures were lower on shallow reefs with high rugosity, but they significantly preferred diseased corals over sea rods. This may be due to the surfaces on the coral heads being newly opened for turf algal colonization, a preferred foraging substrate for all parrotfish species, which commonly inhabit these reefs [51]. Alternatively, the newly available coral tissue could be providing nutritional benefits or a greater concentration of autotrophic organisms that attracted parrotfish foragers [64]. Herbivores on artificial structures were not significantly different from on natural structures, perhaps because after 2 weeks, we observed that the artificial structures had accumulated enough biofilm to be a suitable foraging substrate (Figure 3). The functionally important role that parrotfish play on Caribbean coral reefs is well understood [65,66], and our study suggests that with the flattening of reefs, the reefs of the future may see a decline in parrotfish abundance.
Piscivores rarely used either natural or artificial structures and were unrelated to any of the three component scores of reef substrate. This is unsurprising, due to previous findings that piscivore abundance is associated more with prey availability than any habitat characteristic [67,68]. In contrast, omnivores used natural structures significantly more than artificial structures and decreased structure use on shallow reefs with high rugosity, a response that has been predicted in recent models [31]. However, omnivores’ preference for low-rugosity reefs contradicts previous literature that has found that all feeding groups tend to be positively associated with increased complexity [11,12,13,69]. We expected that rugosity and hard coral cover would be correlated [25], but found that rugosity was not a function of hard coral cover. Presently, the reefs in the Florida Keys are composed of scattered boulder corals, abundant soft corals, and limestone ledges. Additionally, with the recent outbreak of stony coral tissue loss disease, there has been a decrease in live hard coral cover but a lingering presence of dead hard coral structures [70]. With time, overall rugosity and the presence of scattered boulder corals will continue decreasing as dead coral heads begin eroding away.
Similarly, invertivores used natural structures significantly more than artificial structures, but in contrast to the herbivores and omnivores, they increased on shallow reefs with greater rugosity. This suggests that grunts may be the one feeding guild most impacted by the loss of structural complexity and the flattening of the reef. This relationship is best explained by the reef component score associated with rugosity rather than the component score associated with hard coral cover, which suggests that it is physical habitat that matters more to grunts than the health status of the coral. Invertivores often depend on finer-scale shelters for their prey species to occupy and would explain why they, and their prey, would be negatively impacted by structural loss [71]. This negative response of invertivores to reef decline has been predicted as a response to climate change according to predictive climate change models [31].
All guilds had a predominant species that did not mirror the pattern of the rest of the guild. Bicolor damselfish (Stegastes partitus), striped parrotfish (Scarus iseri), blue-striped grunts (Haemulon sciurus), white grunts (Haemulon plumierii), schoolmaster snapper (Lutjanus apodus), and the bluehead wrasses (Thalassoma bifasciatum) had a disproportionate representation within their associated functional feeding guilds, but their patterns did not parallel the overall effect observed within their guild. Their presence may be due to their being habitat generalists, equally at home in hard coral- or soft coral-dominated reefs [67,72]. The reduced structure use by the bicolor damselfish, striped parrotfish, or bluehead wrasses could be explained by the higher abundance of piscivores, although we did not observe an increase in piscivore presence, even though these habitats had an abundance of prey, which should have driven their numbers up [67,68,71,72].
Our results suggest that use of structures, both natural and artificial structures, differs among reef fish functional feeding guilds. If the low-rugosity reefs with low hard coral cover are representative of the future reefs of the middle Florida Keys, we would predict a shift in reef fish community with increasing proportions of herbivores and omnivores, and a decreasing proportion of invertivores. These results can be used as a predictive model for reef fish community responses to changes in reef composition and may be useful in the design of future marine protected areas needed to preserve feeding guilds critical to the recovery of hard corals. Future studies should examine how the relative abundance of fish functional feeding guilds changes in response to this transition from hard-coral-dominated to soft-coral-dominated reefs in the middle Florida Keys.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/oceans2030036/s1. Table S1: Species included in this study, their functional feeding guild, their presence (Y or N) at artificial and natural structures, and their habitat use. Artificial structures = soft coral mimic (SC), control structure (C), boulder coral mimic (BC), and elkhorn coral mimic (EC). Natural structures = sea rods (SR), dead corals (DC), infected diseased corals (IC), and healthy corals (HC). Fish are separated into generalist or specialist habitat use classifications [29]. Table S2: Percent presence of generalist versus specialist species observed for the four artificial and four natural shelter types for each functional feeding guild. Artificial structures = soft coral mimic (SC), control structure (C), boulder coral mimic (BC), and elkhorn coral mimic (EC). Natural structures = sea rods (SR), dead corals (DC), infected diseased corals (IC), and healthy corals (HC).
Author Contributions
Conceptualization, K.N., K.S. (Kylie Smith) and M.C.; methodology, K.N., K.S. (Kylie Smith) and M.C.; formal analysis, K.N., T.F. and M.C.; investigation, K.N.; resources, K.N., K.S. (Kylie Smith) and M.C.; data curation, K.N., T.F., K.M. and K.S. (Kelsey Sox); writing—original draft preparation, K.N., T.F. and M.C.; writing—review and editing, K.N., K.M., K.S. (Kelsey Sox) and K.S. (Kylie Smith); supervision, K.N. and M.C.; funding acquisition, K.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the American Museum of Natural History Lerner-Gray Fund for Marine Research, the American Society of Ichthyologists and Herpetologists Raney Award, Clemson University’s Creative Inquiry Initiative, the Explorer’s Club Mamont Grant, the South Carolina Space Grant Consortium Kathryn D. Sullivan Earth and Marine Science Fellowship (# 2014105), and Clemson University.
Institutional Review Board Statement
This study was conducted according the guidelines of the Florida Keys National Marine Sanctuary (permit numbers FKNMS-2017-032, approved on 6 January 2017, and permit number FKNMS-2018-119, approved on 10 January 2018).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is archived at Clemson University and is available upon request to Michael Childress ([email protected]).
Acknowledgments
We thank Reanna Jeanes, Morgan Gardner, Riley Garvey, Rachel Radick, and Emma Crowfoot for assistance in data collection and processing. We also thank the two anonymous referees and the special issue editor for their great assistance in improving the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Weber, M.L.; Gradwohl, J. Life in the seas. In Weber MLaJG (ed) The Wealth of Oceans; W.W. Norton & Company: New York, NY, USA, 1995. [Google Scholar]
- Spalding, M.; Grenfell, A.M. New estimates of global and regional coral reef areas. Coral Reefs 1997, 16, 225–230. [Google Scholar] [CrossRef]
- Roberts, C.M.; McClean, C.J.; Veron, J.E.; Hawkins, J.P.; Allen, G.R.; McAllister, D.E.; Mittermeier, C.G.; Schueler, F.W.; Spalding, M.; Wells, F.; et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 2002, 295, 1280–1284. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nystrom, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Allen, G.R. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 541–556. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Gill, J.A.; Dulvy, N.K.; Perry, A.L.; Watkinson, A.R.; Côté, I.M. Drivers of region-wide declines in architectural complexity on Caribbean reefs. Coral Reefs 2011, 30, 1051–1060. [Google Scholar] [CrossRef]
- Risk, M.J. Fish Diversity on a Coral Reef in the Virgin Islands. Atoll Res. Bull. 1972, 153, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Luckhurst, B.E.; Luckhurst, K.L. Analysis of the influence of substrate variables onc oral reef fish communities. Mar. Biol. 1978, 49, 317–323. [Google Scholar] [CrossRef]
- Roberts, C.M.; Ormond, R.F.G. Habitat complexity and coral reef fish diversity and abundance on Red Sea fringing reefs. Mar. Ecol. Prog. Ser. 1987, 41, 1–8. [Google Scholar] [CrossRef]
- Ohman, M.C.; Rajasuriya, A. Relationships between habitat structure and fish communities on coral and sandstone reefs. Environ. Biol. Fishes 1998, 53, 19–31. [Google Scholar] [CrossRef]
- Gratwicke, B.; Speight, M.R. The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. J. Fish Biol. 2005, 66, 650–667. [Google Scholar] [CrossRef]
- Gonzalez-Rivero, M.; Harborne, A.R.; Herrera-Reveles, A.; Bozec, Y.M.; Rogers, A.; Friedman, A.; Ganase, A.; Hoegh-Guldberg, O. Linking fishes to multiple metrics of coral reef structural complexity using three-dimensional technology. Sci. Rep. 2017, 7, 13965. [Google Scholar] [CrossRef]
- Richardson, L.E.; Graham, N.A.J.; Pratchett, M.S.; Hoey, A.S. Structural Complexity Mediates Functional Structure of Reef Fish Assemblages among Coral Habitats. Environ. Biol. Fish. 2017, 100, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Molles, M.C. Fish species Diversity on Model and Natural Reef Patches: Experimental Insular Biogeography. Ecol. Monogr. 1978, 48, 289–305. [Google Scholar] [CrossRef]
- Carpenter, K.E. The influence of substrate structure on the local abundance and diversity of philippine reef fishes. In Proceedings of the Fourth International Coral Reef Syposium, Marine Sciences Center, University of the Philippines, Manila, Phillippines, 18–22 May 1981. [Google Scholar]
- Bell, J.O.; Galzin, R. Influence of live coral cover on coral-reef fish communities. Mar. Ecol. Prog. Ser. 1984, 15, 265–274. [Google Scholar] [CrossRef]
- Guidetti, P. Differences among fish assemblages associated with nearshore Posidonia oceanica Seagrass Beds, Rocky-algal Reefs and Unvegetated Sand Habitats in the Adriatic Sea. Estuarine. Coast. Shelf Sci. 2000, 50, 515–529. [Google Scholar] [CrossRef]
- Khalaf, M.A.; Kochzius, M. Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba, Red Sea. Mar. Ecol. Prog. Ser. 2002, 239, 287–299. [Google Scholar] [CrossRef]
- Harborne, A.R.; Rogers, A.; Bozec, Y.M.; Mumby, P.J. Multiple Stressors and the Functioning of Coral Reefs. Ann. Rev. Mar. Sci. 2017, 9, 445–468. [Google Scholar] [CrossRef]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.; Palumbi, S.R. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.V.C. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Chang. Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Munday, P.L.; Wilson, S.K.; Graham, N.A.J.; Cinner, J.E.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.C.; McClanahan, T.R. Effects of climate-induced coral bleaching on coral-reef fishes-ecological and economic consequences. Mar. Biol. Annu. Rev. 2008, 46, 257–302. [Google Scholar]
- Alvarez-Filip, L.; Dulvy, N.K.; Gill, J.A.; Cote, I.M.; Watkinson, A.R. Flattening of Caribbean coral reefs: Region-wide declines in architectural complexity. Proc. Biol. Sci. 2009, 276, 3019–3025. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Filip, L.; Gill, J.A.; Dulvy, N.K. Complex reef architecture supports more small-bodied and longer food chains on Caribbean reefs. Ecosphere 2011, 2, 1–17. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K.; Messmer, V.; Graham, N.A.J. Changes in biodiversity and functioning of reef fish assemblages following coral bleaching and coral loss. Diversity 2011, 3, 424–452. [Google Scholar] [CrossRef] [Green Version]
- Alevizon, W.S.; Porter, J.W. Coral loss and fish guild stability on a Caribbean coral reef: 2014, 1974–2000. Environ. Biol. Fishes 2015, 98, 1035–1045. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K. Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Curr. Opin. Environ. Sustain. 2014, 7, 37–43. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Paddack, M.J.; Collen, B.; Robertson, D.R.; Cote, I.M. Simplification of Caribbean Reef-Fish Assemblages over Decades of Coral Reef Degradation. PLoS ONE 2015, 10, e0126004. [Google Scholar] [CrossRef] [Green Version]
- Paddack, M.J.; Reynolds, J.D.; Aguilar, C.; Appeldoorn, R.S.; Beets, J.; Burkett, E.W.; Chittaro, P.M.; Clarke, K.; Esteves, R.; Fonseca, A.C.; et al. Recent region-wide declines in Caribbean reef fish abundance. Curr. Biol. 2009, 19, 590–595. [Google Scholar] [CrossRef]
- Inagaki, K.Y.; Pennino, M.G.; Floeter, S.R.; Hay, M.E.; Longo, G.O. Trophic interactions will expand geographically but be less intense as oceans warm. Glob. Chang. Biol. 2020, 26, 6805–6812. [Google Scholar] [CrossRef]
- Bohnsack, J.A. Are high densities of fishes at artificial reefs the result of habitat limitation or behavioral preference? Bull. Mar. Sci. 1989, 44, 631–645. [Google Scholar]
- Bohnsack, J.A.; Harper, D.E.; McClellan, D.B.; Hulsbeck, M. Effects of reef size on colonization and assemblage structure of fishes at artificial reefs off southeastern Florida, USA. Bull. Mar. Sci. 1994, 55, 796–823. [Google Scholar]
- Carr, M.H.; Hixon, M.A. Artificial reefs: The importance of comparisons with natural reefs. Fisheries 1997, 22, 28–33. [Google Scholar] [CrossRef]
- Clark, S.; Edwards, A.J. An evaluation of artificial reef structures as tools for marine habitat rehabilitation in the Maldives. Aquat. Conserv. Mar. Freshw. Ecosyst. 1999, 9, 5–21. [Google Scholar] [CrossRef]
- Rilov, G.; Benayahu, Y. Fish assemblage on natural versus vertical artificial reefs: The rehabilitation perspective. Mar. Biol. 2000, 136, 931–942. [Google Scholar] [CrossRef]
- Abelson, A.; Shlesinger, Y. Comparison of the development of coral and fish communities on rock-aggregated artificial reefs in Eilat, Red Sea. ICES J. Mar. Sci. 2002, 59, S122–S126. [Google Scholar] [CrossRef]
- Edwards, R.A.; Smith, S.D. Subtidal assemblages associated with a geotextile reef in south-east Queensland, Australia. Mar. Freshw. Res. 2005, 56, 133–142. [Google Scholar] [CrossRef]
- Clynick, B.G.; Chapman, M.G.; Underwood, A.J. Fish assemblages associated with urban structures and natural reefs in Sydney, Australia. Austral Ecol. 2008, 33, 140–150. [Google Scholar] [CrossRef]
- Bohnsack, J.A. Habitat structure and the design of artificial reefs. In Habitat Structure; Springer: Dordrecht, The Netherlands, 1991; pp. 41–426. [Google Scholar]
- Rilov, G.; Benayahu, Y. Rehabilitation of coral reef-fish communities: The importance of artificial-reef relief to recruitment rates. Bull. Mar. Sci. 2002, 70, 185–197. [Google Scholar]
- Rilov, G.; Benayahu, Y. Vertical artificial structures as an alternative habitat for coral reef fishes in disturbed environments. Mar. Environ. Res. 1998, 45, 431–451. [Google Scholar] [CrossRef]
- Alevizon, W.S.; Gorham, J.C. Effects of artificial reef deployment on nearby resident fishes. Bull. Mar. Sci. 1989, 44, 646–661. [Google Scholar]
- Pérez-Ruzafa, A.; Garcıa-Charton, J.A.; Barcala, E.; Marcos, C. Changes in benthic fish assemblages as a consequence of coastal works in a coastal lagoon: The Mar Menor (Spain, Western Mediterranean). Mar. Pollut. Bull. 2006, 53, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Burt, J.; Bartholomew, A.; Usseglio, P.; Bauman, A.; Sale, P.F. Are artificial reefs surrogates of natural habitats for corals and fish in Dubai, United Arab Emirates? Coral Reefs 2009, 28, 663–675. [Google Scholar] [CrossRef]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Halpern, B.S.; Floeter, S.R. Functional diversity responses to changing species richness in reef fish communities. Mar. Ecol. Prog. Ser. 2008, 364, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.R.; Gunderson, L.; Johnson, A.R. The use of discontinuities and functional groups to assess relative resilience in complex systems. Ecosystems 2005, 8, 958–966. [Google Scholar] [CrossRef]
- Folke, C. Resilience: The emergence of a perspective for social-ecological systems analyses. Glob. Environ. Chang. 2006, 16, 253–267. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B.; Blomberg, S.P.; Montague-Drake, R.; Felton, A.; Stein, J.A. Functional richness and relative resilience of bird communities in regions with different land use intensities. Ecosystems 2007, 10, 964–974. [Google Scholar] [CrossRef]
- Smith, K.M.; Quirk-Royal, B.E.; Drake-Lavelle, K.; Childress, M.J. Influences of ontogenetic phase and resource availability on parrotfish foraging preferences in the Florida Keys, FL USA. Mar. Ecol. Prog. Ser. 2018, 603, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.M.; Payton, T.G.; Sims, R.J.; Stroud, C.S.; Jeanes, R.C.; Hyatt, T.B.; Childress, M.J. Impacts of consecutive bleaching events on transplanted coral colonies in the Florida Keys. Coral Reefs 2019, 38, 851–861. [Google Scholar] [CrossRef]
- Manzello, D.P.; Enochs, I.C.; Kolodziej, G.; Carlton, R. Recent decade of growth and calcification of Orbicella faveolata in the florida keys: An inshore-offshore comparison. Mar. Ecol. Prog. Ser. 2015, 521, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Thresher, R.E.; Colin, P.L. Trophic structure, diversity and abundance of fishes of the deep reef (30–300 m) at Enewetak, Marshall Islands. Bull. Mar. Sci. 1986, 38, 253–272. [Google Scholar]
- Ginsburg, R.N.; Shinn, E.A. Preferential distribution of reefs in the Florida reef tract: The past is the key to the present. In Proceedings of the Colloquium on Global Aspects of Coral Reefs: Health, Hazards and History; Ginsburg, R.N., Ed.; Rosenstiel School of Marine and Atmospheric Science, University of Miami: Coral Gables, FL, USA, 1994; pp. 21–26. [Google Scholar]
- Leichter, J.J.; Stewart, H.L.; Miller, S.L. Episodic nutrient transport to Florida coral reefs. Limnol. Oceanogr. 2003, 48, 1394–1407. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Renema, W.; Rosen, B.R. Biodiversity hotspots, evolution and coral reef biogeography. In Biotic Evolution and Environmental Change in Southeast Asia; Cambridge University Press: Cambridge, UK, 2012; Volume 216. [Google Scholar]
- Nemeth, M.; Appeldoorn, R. The distribution of herbivorous coral reef fishes within fore-reef habitats: The role of depth, light and rugosity. Caribb. J. Sci. 2009, 45, 247–253. [Google Scholar] [CrossRef]
- Harborne, A.R.; Mumby, P.J.; Ferrari, R. The effectiveness of different meso-scale rugosity metrics for predicting intra-habitat variation in coral-reef fish assemblages. Environ. Biol. Fishes 2012, 94, 431–442. [Google Scholar] [CrossRef]
- Gordon, T.A.; Radford, A.N.; Davidson, I.K.; Barnes, K.; McCloskey, K.; Nedelec, S.L.; Simpson, S.D. Acoustic enrichment can enhance fish community development on degraded coral reef habitat. Nat. Commun. 2019, 10, 5414. [Google Scholar] [CrossRef]
- Hu, Y.; Majoris, J.E.; Buston, P.M.; Webb, J.F. Potential roles of smell and taste in the orientation behaviour of coral-reef fish larvae: Insights from morphology. J. Fish Biol. 2019, 95, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Komyakova, V.; Chamberlain, D.; Jones, G.P.; Swearer, S.E. Assessing the performance of artificial reefs as substitute habitat for temperate reef fishes: Implications for reef design and placement. Sci. Total. Environ. 2019, 668, 139–152. [Google Scholar] [CrossRef]
- Komyakova, V.; Swearer, S.E. Contrasting patterns in habitat selection and recruitment of temperate reef fishes among natural and artificial reefs. Mar. Environ. Res. 2019, 143, 71–81. [Google Scholar] [CrossRef]
- Gutiérrez-Isaza, N.; Espinoza-Avalos, J.; León-Tejera, H.P.; González-Solís, D. Endolithic community composition of Orbicella faveolata (Scleractinia) underneath the interface between coral tissue and turf algae. Coral Reefs 2015, 34, 625–630. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Hay, M.E. Impact of herbivore identity on algal succession and coral growth on a Caribbean reef. PLoS ONE 2010, 5, e8963. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.C.; Soares, M.C.; Oxenford, H.A.; Côté, I.M. Interspecific differences in foraging behaviour and functional role of Caribbean parrotfish. Mar. Biodivers. Rec. 2009, 2, e148. [Google Scholar] [CrossRef]
- Hixon, M.A.; Beets, J.P. Predation, prey refuges, and the structure of coral-reef fish assemblages. Ecol. Monogr. 1993, 63, 77–101. [Google Scholar] [CrossRef] [Green Version]
- Stewart, B.D.; Jones, G.P. Associations between the abundance of piscivorous fishes and their prey on coral reefs: Implications for prey-fish mortality. Mar. Biol. 2001, 138, 383–397. [Google Scholar] [CrossRef]
- Hensel, E.; Allgeier, J.E.; Layman, C.A. Effects of predator presence and habitat complexity on reef fish communities in The Bahamas. Mar. Biol. 2019, 166, 136. [Google Scholar] [CrossRef]
- Noonan, K.R.; Childress, M.J. Association of butterflyfishes and stony coral tissue loss disease in the Florida Keys. Coral Reefs 2020, 39, 1581–1590. [Google Scholar] [CrossRef]
- Yates, D.C.; Lonhart, S.I.; Hamilton, S.L. Effects of marine reserves on predator-prey interactions in central California kelp forests. Mar. Ecol. Prog. Ser. 2020, 655, 139–155. [Google Scholar] [CrossRef]
- Boaden, A.E.; Kingsford, M.J. Predators drive community structure in coral reef fish assemblages. Ecosphere 2015, 6, 1–33. [Google Scholar] [CrossRef]
Figure 1.
Map to show the location of the eight study sites in the middle of Florida Keys, USA, surveyed during the summers of 2018 and 2019. These eight sites are located inside the Florida Keys National Marine Sanctuary.
Figure 1.
Map to show the location of the eight study sites in the middle of Florida Keys, USA, surveyed during the summers of 2018 and 2019. These eight sites are located inside the Florida Keys National Marine Sanctuary.

Figure 2.
Reef fishes’ use of structures was monitored on natural and artificial structures. Natural structures (top) included sea rods, and dead, actively diseased, and healthy boulder corals. Artificial structures (bottom) included a control and three structures that were built to mimic historically abundant natural structures including soft coral, boulder coral, and branching coral (elkhorn coral, Acropora palmata).
Figure 2.
Reef fishes’ use of structures was monitored on natural and artificial structures. Natural structures (top) included sea rods, and dead, actively diseased, and healthy boulder corals. Artificial structures (bottom) included a control and three structures that were built to mimic historically abundant natural structures including soft coral, boulder coral, and branching coral (elkhorn coral, Acropora palmata).

Figure 3.
Log n (ln) mean per structure of the counts for all reef fishes combined (gray) and for each functional group—herbivores (green), omnivores (orange), predators (blue), and invertivores (yellow)—obtained during the time-lapse photography observations on the different natural and artificial corals. Error bars are ±1 SEM. Differences in the letter located over each bar indicate when values were shown to be statistically different from one another by Tukey’s post hoc comparisons.
Figure 3.
Log n (ln) mean per structure of the counts for all reef fishes combined (gray) and for each functional group—herbivores (green), omnivores (orange), predators (blue), and invertivores (yellow)—obtained during the time-lapse photography observations on the different natural and artificial corals. Error bars are ±1 SEM. Differences in the letter located over each bar indicate when values were shown to be statistically different from one another by Tukey’s post hoc comparisons.

Figure 4.
Linear regressions of mean numbers of fish observed (log-transformed) for the structure surveys within a reef site versus the three component scores that best describe the reef characteristics. The three component scores, from left to right, are topography (PC1), algae (PC2), and coral cover (PC3). The specific loadings of the reef measures for each component score are in Table 1. Graphs include all reef fish combined (gray), herbivores (green), omnivores (orange), invertivores (yellow), and predators (blue).
Figure 4.
Linear regressions of mean numbers of fish observed (log-transformed) for the structure surveys within a reef site versus the three component scores that best describe the reef characteristics. The three component scores, from left to right, are topography (PC1), algae (PC2), and coral cover (PC3). The specific loadings of the reef measures for each component score are in Table 1. Graphs include all reef fish combined (gray), herbivores (green), omnivores (orange), invertivores (yellow), and predators (blue).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Principal component score correlations for reef substrate measures. Bold indicates loadings with p < 0.05.
Table 1.
Principal component score correlations for reef substrate measures. Bold indicates loadings with p < 0.05.
| Measures | PC 1 Topo | PC 2 Algal Cover | PC 3 Coral Cover |
|---|---|---|---|
| Rugosity | −0.8573 | −0.0397 | −0.1667 |
| Distance to shore | 0.9207 | 0.1501 | 0.0139 |
| Depth | 0.7603 | 0.2546 | 0.0807 |
| % Sand | 0.2249 | 0.8930 | −0.1249 |
| % Fleshy algae | 0.7219 | −0.5776 | −0.0785 |
| % Calcareous | −0.2716 | 0.7073 | −0.0971 |
| % Turf algae | −0.0191 | −0.8884 | 0.2747 |
| % Hard coral | −0.4116 | 0.2856 | 0.3895 |
| % Soft coral | −0.3962 | −0.3304 | −0.7455 |
| % Sponge | −0.6138 | −0.0196 | 0.4706 |
Table 2.
Nested analysis of covariance with fish abundance (from video transect surveys), topography, algal cover, and coral cover as covariates and structure type (control, soft, barrel, elkhorn, sea rod, dead coral, diseased coral, and healthy coral) nested within structure state (natural versus artificial). Bolded values indicate significant values.
Table 2.
Nested analysis of covariance with fish abundance (from video transect surveys), topography, algal cover, and coral cover as covariates and structure type (control, soft, barrel, elkhorn, sea rod, dead coral, diseased coral, and healthy coral) nested within structure state (natural versus artificial). Bolded values indicate significant values.
| Variable | Source | Df | F Ratio | p | adj r2 |
|---|---|---|---|---|---|
| All Fish | All Fish # | 1 | 5.620 | 0.0189 | 0.122 |
| Topography (PC1) | 1 | 7.182 | 0.0058 | ||
| Algae (PC2) | 1 | 0.163 | 0.6867 | ||
| Coral (PC3) | 1 | 1.071 | 0.3021 | ||
| Structure Type | 1 | 6.495 | 0.0117 | ||
| Structure State | 6 | 10.24 | 0.0015 | ||
| Herbivores | Herbivore # | 1 | 8.412 | 0.0042 | 0.1844 |
| Topography (PC1) | 1 | 17.25 | 0.0001 | ||
| Algae (PC2) | 1 | 1.536 | 0.2169 | ||
| Coral (PC3) | 1 | 0.909 | 0.3417 | ||
| Structure Type | 1 | 1.733 | 0.1898 | ||
| Structure State | 6 | 2.523 | 0.0232 | ||
| Omnivores | Omnivore # | 1 | 0.964 | 0.3275 | 0.1909 |
| Topography (PC1) | 1 | 28.691 | 0.0001 | ||
| Algae (PC2) | 1 | 0.020 | 0.8876 | ||
| Coral (PC3) | 1 | 0.470 | 0.4938 | ||
| Structure Type | 1 | 5.627 | 0.0189 | ||
| Structure State | 6 | 0.463 | 0.8344 | ||
| Invertivores | Invertivore # | 1 | 8.169 | 0.0050 | 0.3227 |
| Topography (PC1) | 1 | 11.78 | 0.0008 | ||
| Algae (PC2) | 1 | 0.224 | 0.6381 | ||
| Coral (PC3) | 1 | 0.627 | 0.4300 | ||
| Structure Type | 1 | 6.573 | 0.0116 | ||
| Structure State | 6 | 0.968 | 0.4499 | ||
| Predators | Predator # | 1 | 5.882 | 0.0170 | 0.1106 |
| Topography (PC1) | 1 | 1.231 | 0.2697 | ||
| Algae (PC2) | 1 | 0.957 | 0.3300 | ||
| Coral (PC3) | 1 | 0.255 | 0.6143 | ||
| Structure Type | 1 | 0.208 | 0.6491 | ||
| Structure State | 6 | 0.743 | 0.6163 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Noonan, K.; Fair, T.; Matthee, K.; Sox, K.; Smith, K.; Childress, M. Reef Fish Associations with Natural and Artificial Structures in the Florida Keys. Oceans 2021, 2, 634-647. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030036
AMA Style
Noonan K, Fair T, Matthee K, Sox K, Smith K, Childress M. Reef Fish Associations with Natural and Artificial Structures in the Florida Keys. Oceans. 2021; 2(3):634-647. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030036
Chicago/Turabian StyleNoonan, Kara, Thomas Fair, Kristiaan Matthee, Kelsey Sox, Kylie Smith, and Michael Childress. 2021. "Reef Fish Associations with Natural and Artificial Structures in the Florida Keys" Oceans 2, no. 3: 634-647. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans2030036