Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
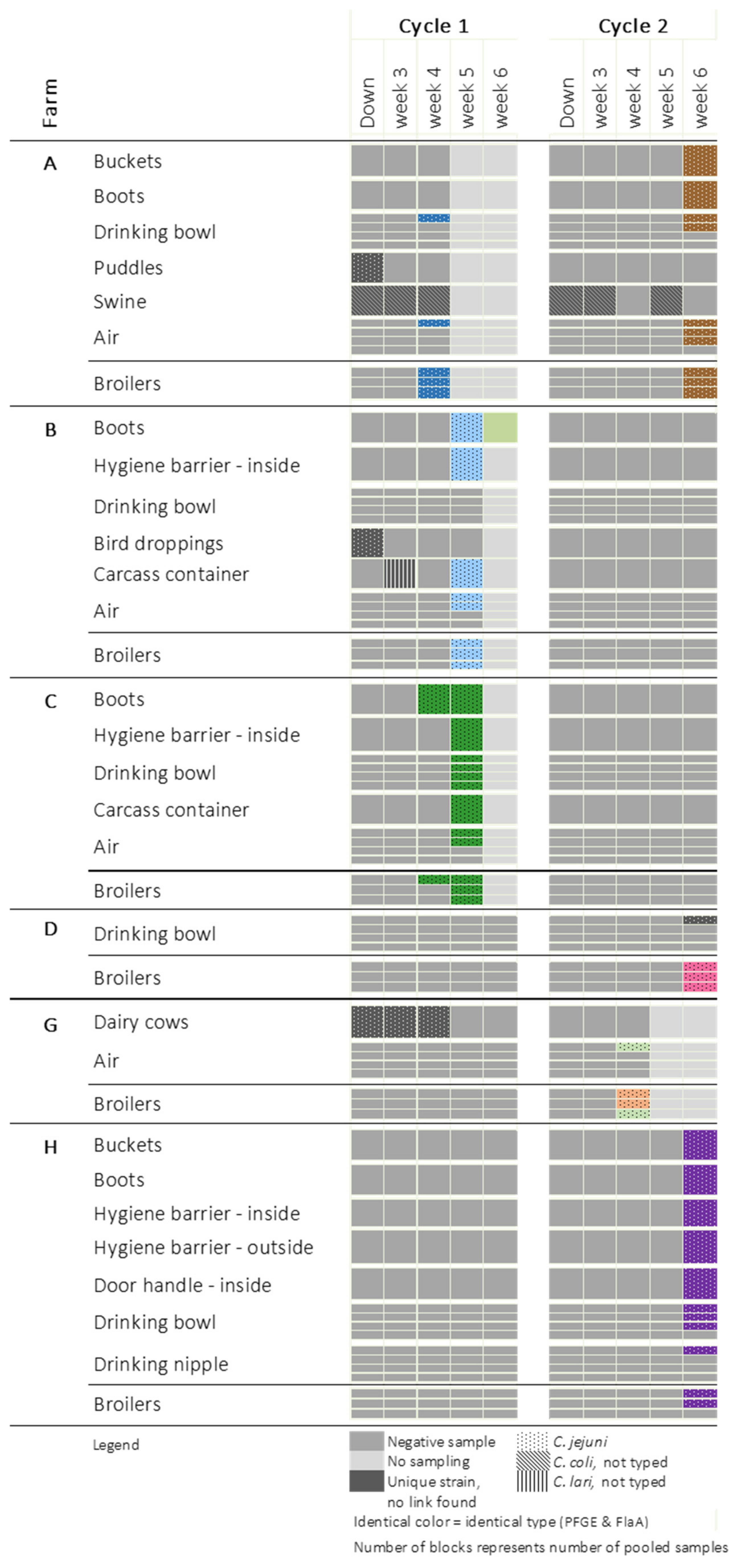
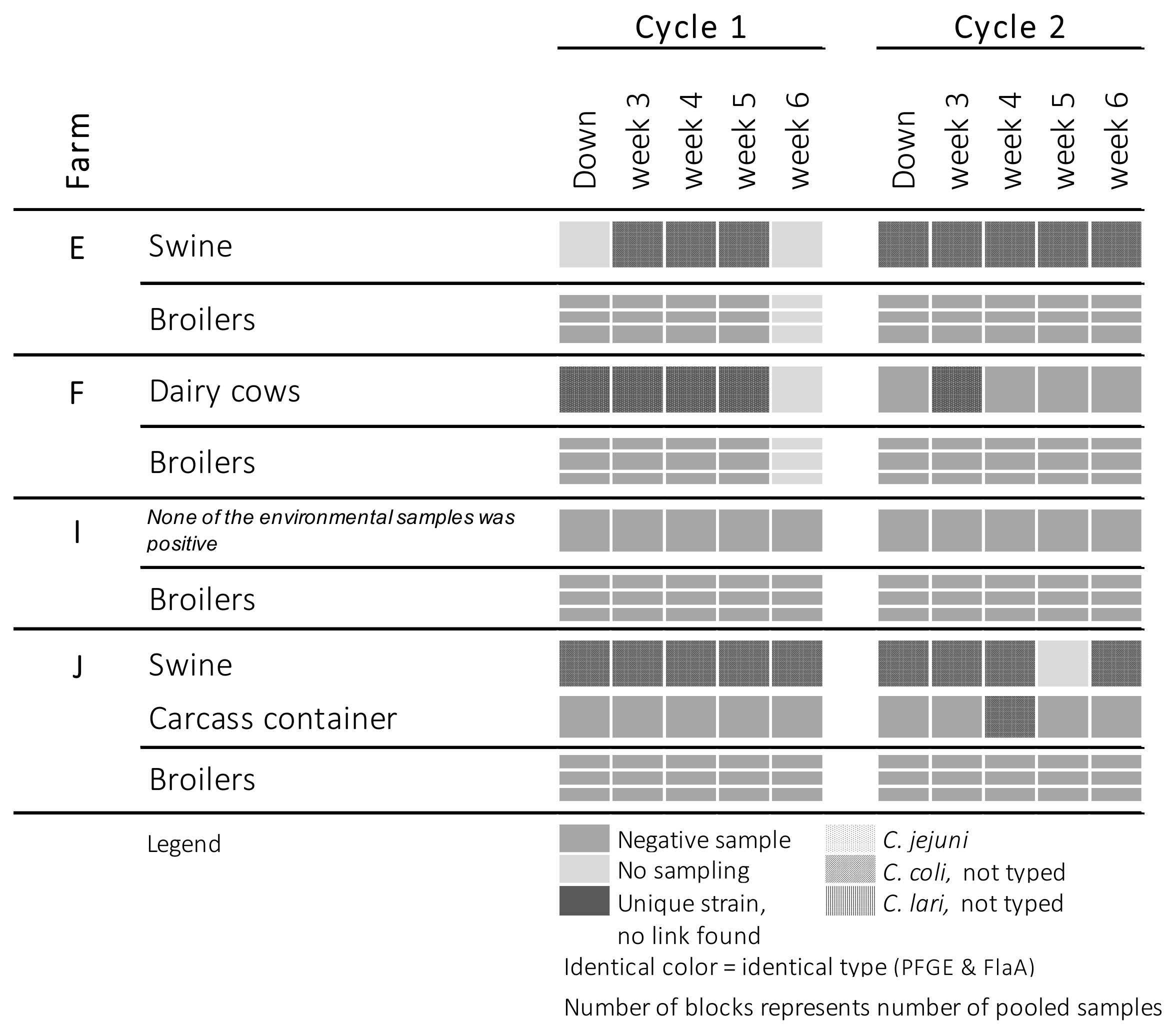
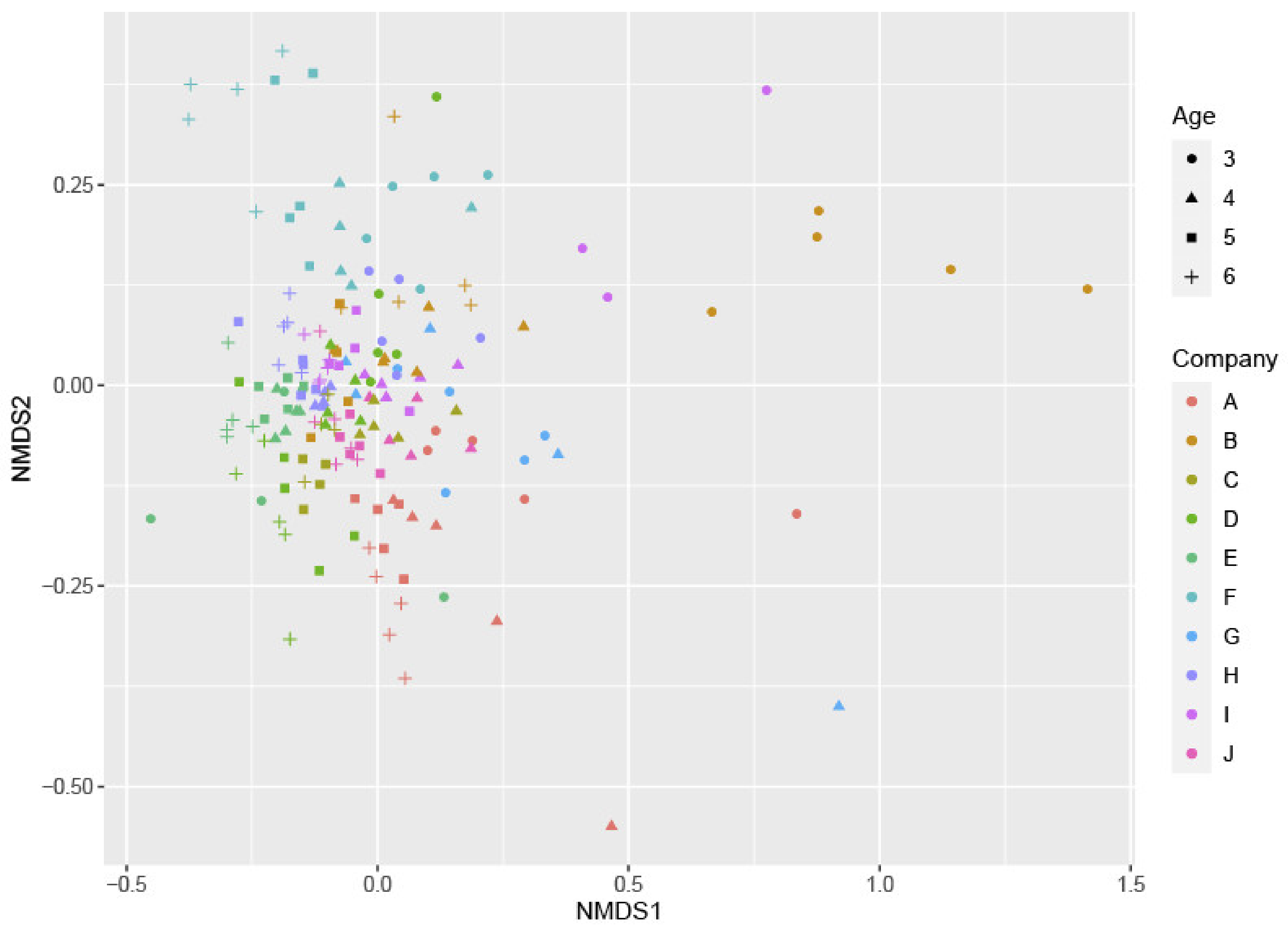
2. Results
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Microbiological Analysis
4.3. Molecular Analysis
4.4. 16 S Metabarcoding
4.4.1. DNA Extraction, Library Preparation and Sequencing
4.4.2. Processing of Sequence Reads and Downstream Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EFSA; ECDC. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar] [CrossRef] [Green Version]
- EFSA; ECDC. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2009. EFSA J. 2011, 9. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Pijnacker, R.; Coipan, C.; Mulder, A.C.; Veludo, A.F.; De Rijk, S.; Van Hoek, A.H.; Buij, R.; Muskens, G.; Koene, M.; et al. Sources and transmission routes of campylobacteriosis: A combined analysis of genome and exposure data. J. Infect. 2020, 20, 30733–30737. [Google Scholar] [CrossRef]
- Messens, W.; Hartnett, E.; Gellynck, X.; Viaene, J.; Halet, D.; Herman, L.; Grijspeerdt, K. Quantitative Risk Assessment of Human Campylobacteriosis through the Consumption of Chicken Meat in Belgium. In Proceedings of the 18th European Symposium on the Quality of Poultry Meat, Prague, Czech Republic, 2–5 September 2007; pp. 167–168. [Google Scholar]
- Hermans, D.; Pasmans, F.; Messens, W.; Martel, A.; Van Immerseel, F.; Rasschaert, G.; Heyndrickx, M.; Van Deun, K.; Haesebrouck, F. Poultry as a Host for the Zoonotic PathogenCampylobacter jejuni. Vector Borne Zoonotic Dis. 2012, 12, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Wysok, B.; Wojtacka, J. Detection of virulence genes determining the ability to adhere and invade in Campylobacter spp. from cattle and swine in Poland. Microb. Pathog. 2018, 115, 257–263. [Google Scholar] [CrossRef]
- Plishka, M.; Sargeant, J.M.; Greer, A.L.; Hookey, S.; Winder, C. The Prevalence of Campylobacter in Live Cattle, Turkey, Chicken, and Swine in the United States and Canada: A Systematic Review and Meta-Analysis. Foodborne Pathog. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, C.; Scheu, K.D.; Keel, M.; Renggli, F.; Stephan, R. Occurrence and genotypes of Campylobacter in broiler flocks, other farm animals, and the environment during several rearing periods on selected poultry farms. Int. J. Food Microbiol. 2008, 125, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Robyn, J.; Rasschaert, G.; Pasmans, F.; Heyndrickx, M. Thermotolerant Campylobacter during Broiler Rearing: Risk Factors and Intervention. Compr. Rev. Food Sci. F 2015, 14, 81–105. [Google Scholar] [CrossRef]
- Agunos, A.; Waddell, L.; Léger, D.; Taboada, E. A Systematic Review Characterizing On-Farm Sources of Campylobacter spp. for Broiler Chickens. PLoS ONE 2014, 9, e104905. [Google Scholar] [CrossRef] [Green Version]
- Georgiev, M.; Beauvais, W.; Guitian, J. Effect of enhanced biosecurity and selected on-farm factors on Campylobacter colonization of chicken broilers. Epidemiol. Infect. 2016, 145, 553–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hald, B.; Sommer, H.M.; Skovgård, H. Use of Fly Screens to ReduceCampylobacterspp. Introduction in Broiler Houses. Emerg. Infect. Dis. 2007, 13, 1951–1953. [Google Scholar] [CrossRef]
- Bahrndorff, S.; Rangstrup-Christensen, L.; Nordentoft, S.; Hald, B. Foodborne Disease Prevention and Broiler Chickens with ReducedCampylobacterInfection. Emerg. Infect. Dis. 2013, 19, 425–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Willer, T.; Colin, P.; Pielsticker, C.; Rychlik, I.; Velge, P.; Kaspers, B.; Rautenschlein, S. Influence of the Gut Microbiota Composition on Campylobacter jejuni Colonization in Chickens. Infect. Immun. 2017, 85, e00380-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seliwiorstow, T.; Duarte, A.; Baré, J.; Botteldoorn, N.; Dierick, K.; Uyttendaele, M.; De Zutter, L. Comparison of Sample Types and Analytical Methods for the Detection of Highly Campylobacter-Colonized Broiler Flocks at Different Stages in the Poultry Meat Production Chain. Foodborne Pathog. Dis. 2015, 12, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Rasschaert, G.; Houf, K.; Van Hende, J.; De Zutter, L. Investigation of the concurrent colonization with Campylobacter and Salmonella in poultry flocks and assessment of the sampling site for status determination at slaughter. Vet. Microbiol. 2007, 123, 104–109. [Google Scholar] [CrossRef]
- Herman, L.; Heyndrickx, M.; Grijspeerdt, K.; Vandekerchove, D.; Rollier, I.; De Zutter, L. Routes for Campylobacter contamination of poultry meat: Epidemiological study from hatchery to slaughterhouse. Epidemiol. Infect. 2003, 131, 1169–1180. [Google Scholar] [CrossRef]
- Fernandez, H.; Vergara, M.; Tapia, F. Dessication resistance in thermotolerant campylobacter species. Infection 1985, 13, 197. [Google Scholar] [CrossRef]
- Van Gerwe, T.; Miflin, J.K.; Templeton, J.M.; Bouma, A.; Wagenaar, J.A.; Jacobs-Reitsma, W.F.; Stegeman, A.; Klinkenberg, D. Quantifying Transmission of Campylobacter jejuni in Commercial Broiler Flocks. Appl. Environ. Microbiol. 2008, 75, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Stern, N.J.; Cox, N.A.; Musgrove, M.T.; Park, C.M. Incidence and Levels of Campylobacter in Broilers After Exposure to an Inoculated Seeder Bird. J. Appl. Poult. Res. 2001, 10, 315–318. [Google Scholar] [CrossRef]
- Messens, W.; Herman, L.; De Zutter, L.; Heyndrickx, M. Multiple typing for the epidemiological study of contamination of broilers with thermotolerant Campylobacter. Vet. Microbiol. 2009, 138, 120–131. [Google Scholar] [CrossRef]
- Sparks, N. The role of the water supply system in the infection and control of Campylobacter in chicken. World Poult. Sci. J. 2009, 65, 459–474. [Google Scholar] [CrossRef]
- Newell, D.G.; Fearnley, C. Sources of Campylobacter Colonization in Broiler Chickens. Appl. Environ. Microbiol. 2003, 69, 4343–4351. [Google Scholar] [CrossRef] [Green Version]
- Gregory, E.; Barnhart, H.; Dreesen, D.W.; Stern, N.J.; Corn, J.L. Epidemiological study of Campylobacter spp. in broilers: Source, time of colonization, and prevalence. Avian Dis. 1998, 41, 890. [Google Scholar] [CrossRef]
- Evans, S.J. Introduction and spread of thermophilic campylobacters in broiler flocks. Vet. Rec. 1992, 131, 574–576. [Google Scholar]
- Søndergaard, M.S.R.; Josefsen, M.H.; Löfström, C.; Christensen, L.S.; Wieczorek, K.; Osek, J.; Hoorfar, J. Low-Cost Monitoring of Campylobacter in Poultry Houses by Air Sampling and Quantitative PCR. J. Food Prot. 2014, 77, 325–330. [Google Scholar] [CrossRef]
- Carrique-Mas, J.; Bryant, J.E.; Cuong, N.V.; Hoang, N.V.M.; Campbell, J.; Dung, T.T.N.; Duy, D.; Hoa, N.T.; Thompson, C.; Hien, V.V.; et al. An epidemiological investigation ofCampylobacterin pig and poultry farms in the Mekong delta of Vietnam. Epidemiol. Infect. 2013, 142, 1425–1436. [Google Scholar] [CrossRef] [Green Version]
- Ragimbeau, C.; Schneider, F.; Losch, S.; Even, J.; Mossong, J. Multilocus Sequence Typing, Pulsed-Field Gel Electrophoresis, and fla Short Variable Region Typing of Clonal Complexes of Campylobacter jejuni Strains of Human, Bovine, and Poultry Origins in Luxembourg. Appl. Environ. Microbiol. 2008, 74, 7715–7722. [Google Scholar] [CrossRef] [Green Version]
- Patriarchi, A.; Fox, Á.; Maunsell, B.; Fanning, S.; Bolton, D. Molecular Characterization and Environmental Mapping of Campylobacter Isolates in a Subset of Intensive Poultry Flocks in Ireland. Foodborne Pathog. Dis. 2011, 8, 99–108. [Google Scholar] [CrossRef]
- Rapp, D.; Ross, C.M.; Pleydell, E.J.; Muirhead, R.W. Differences in the Fecal Concentrations and Genetic Diversities of Campylobacter jejuni Populations among Individual Cows in Two Dairy Herds. Appl. Environ. Microbiol. 2012, 78, 7564–7571. [Google Scholar] [CrossRef] [Green Version]
- Hertogs, K.; Heyndrickx, M.; Gelaude, P.; De Zutter, L.; Dewolf, J.; Rasschaert, G. The effect of partial depopulation on Campylobacter introduction in broiler houses. Poult. Sci. 2020. [Google Scholar] [CrossRef]
- Rasschaert, G.; De Zutter, L.; Herman, L.; Heyndrickx, M. Campylobacter contamination of broilers: The role of transport and slaughterhouse. Int. J. Food Microbiol. 2020, 322, 108564. [Google Scholar] [CrossRef]
- Ridley, A.; Morris, V.; Gittins, J.; Cawthraw, S.; Harris, J.; Edge, S.; Allen, V. Potential sources of Campylobacter infection on chicken farms: Contamination and control of broiler-harvesting equipment, vehicles and personnel. J. Appl. Microbiol. 2011, 111, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Chaisowwong, W.; Kusumoto, A.; Hashimoto, M.; Harada, T.; Maklon, K.; Kawamoto, K. Physiological Characterization of Campylobacter jejuni under Cold Stresses Conditions: Its Potential for Public Threat. J. Vet. Med. Sci. 2012, 74, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Yu, H.; Liu, T.; Gill, J.; Chambers, J.; Wheatcroft, R.; Sabour, P. Effects of zinc bacitracin, bird age and access to range on bacterial microbiota in the ileum and caeca of broiler chickens. J. Appl. Microbiol. 2008, 104, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.M.; Impey, C.S.; Stevens, B. Factors affecting the incidence and anti-salmonella activity of the anaerobic caecal flora of the young chick. J. Hyg. 1979, 82, 263–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeant, M.J.; Constantinidou, C.; Cogan, T.A.; Bedford, M.R.; Penn, C.W.; Pallen, M.J. Extensive Microbial and Functional Diversity within the Chicken Cecal Microbiome. PLoS ONE 2014, 9, e91941. [Google Scholar] [CrossRef] [PubMed]
- Walugembe, M.; Hsieh, J.C.F.; Koszewski, N.J.; Lamont, S.J.; Persia, M.E.; Rothschild, M.F. Effects of dietary fiber on cecal short-chain fatty acid and cecal microbiota of broiler and laying-hen chicks. Poult. Sci. 2015, 94, 2351–2359. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important Metabolic Pathways and Biological Processes Expressed by Chicken Cecal Microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- González-Fandos, E.; Maya, N.; Pérez-Arnedo, I. Effect of propionic acid on Campylobacter jejuni attached to chicken skin during refrigerated storage. Int. Microbiol. 2015, 18, 171–175. [Google Scholar]
- Scupham, A.J.; Jones, J.A.; Rettedal, E.A.; Weber, T.E. Antibiotic Manipulation of Intestinal Microbiota to Identify Microbes Associated with Campylobacter jejuni Exclusion in Poultry. Appl. Environ. Microbiol. 2010, 76, 8026–8032. [Google Scholar] [CrossRef] [Green Version]
- Guerin, M.T.; Martin, W.; Reiersen, J.; Berke, O.; McEwen, S.A.; Bisaillon, J.-R.; Lowman, R. A farm-level study of risk factors associated with the colonization of broiler flocks with Campylobacter spp. in Iceland, 2001–2004. Acta Vet. Scand. 2007, 49, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hald, B.; Skovgård, H.; Pedersen, K.; Bunkenborg, H. Influxed Insects as Vectors for Campylobacter jejuni and Campylobacter coli in Danish Broiler Houses. Poult. Sci. 2008, 87, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Nather, G.; Alter, T.; Martin, A.; Ellerbroek, L. Analysis of risk factors for Campylobacter species infection in broiler flocks. Poult. Sci. 2009, 88, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Battersby, T.; Whyte, P.; Bolton, D. The pattern of Campylobacter contamination on broiler farms; external and internal sources. J. Appl. Microbiol. 2016, 120, 1108–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Høg, B.B.; Sommer, H.; Larsen, L.; Sørensen, A.; David, B.; Hofshagen, M.; Rosenquist, H. Farm specific risk factors for Campylobacter colonisation in Danish and Norwegian broilers. Prev. Vet. Med. 2016, 130, 137–145. [Google Scholar] [CrossRef]
- Sommer, H.; Høg, B.B.; Larsen, L.; Sørensen, A.I.V.; Williams, N.J.; Merga, J.; Cerdà-Cuéllar, M.; Urdaneta, S.; Dolz, R.; Wieczorek, K.; et al. Analysis of farm specific risk factors for Campylobacter colonization of broilers in six European countries. Microb. Risk Anal. 2016, 2, 16–26. [Google Scholar] [CrossRef]
- KMI. Available online: https://www.meteo.be/resources/climateReportWeb/klimatologisch_jaaroverzicht_2018.pdf (accessed on 22 December 2020).
- Linton, D.; Lawson, A.J.; Owen, R.J.; Stanley, J. PCR detection, identification to species level, and fingerprinting of Campylobacter jejuni and Campylobacter coli direct from diarrheic samples. J. Clin. Microbiol. 1997, 35, 2568–2572. [Google Scholar] [CrossRef] [Green Version]
- Linton, D.; Owen, R.; Stanley, J. Rapid identification by PCR of the genus Campylobacter and of five Campylobacter species enteropathogenic for man and animals. Res. Microbiol. 1996, 147, 707–718. [Google Scholar] [CrossRef]
- Nachamkin, I.; Bohachick, K.; Patton, C.M. Flagellin gene typing of Campylobacter jejuni by restriction fragment length polymorphism analysis. J. Clin. Microbiol. 1993, 31, 1531–1536. [Google Scholar] [CrossRef] [Green Version]
- PulseNet. Standard Operating Procedure for PulseNet PFGE of Campylobacter jejuni. 2017. Available online: https://www.cdc.gov/pulsenet/pdf/campylobacter-pfge-protocol-508c.pdf (accessed on 22 December 2020).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 22 December 2020).
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 22 December 2020).
- Morgan, M.; Anders, S.; Lawrence, M.; Aboyoun, P.; Pagès, H.; Gentleman, R. ShortRead: A bioconductor package for input, quality assessment and exploration of high-throughput sequence data. Bioinformatics 2009, 25, 2607–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High resolution sample inference from amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-2; The R Foundation: Vienna, Austria, 2019. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2 Elegant Graphics for Data Analysis. J. R. Stat. Soc. Ser. A 2016. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Occurrence (%) a | Log2-Fold Change b | Adjusted p-Value |
|---|---|---|---|---|
| Staphylococcaceae | Staphylococcus | 1.76 | −2.15 | 2.64 × 10−6 |
| Corynebacteriaceae | Corynebacterium | 2.09 | −2.06 | 1.10 × 10−5 |
| Dermabacteraceae | Brachybacterium | 0.90 | −1.86 | 0.000304 |
| Leuconostocaceae | Weisella | 0.81 | −1.79 | 0.000212 |
| Bacteroidaceae | Bacteroides | 0.74 | −1.00 | 0.000194 |
| Lactobacillaceae | Lactobacillus | 7.47 | −0.43 | 0.033147 |
| Ruminococcaceae | Subdoligranulum | 1.08 | 0.25 | 0.041226 |
| Lachnospiraceae | Fusicatenibacter | 1.21 | 0.69 | 9.18 × 10−8 |
| Streptococcaceae | Streptococcus | 1.53 | 2.58 | 5.49 × 10−9 |
| Family | Genus | Occurrence (%) a | Log2-Fold Change b | Adjusted p-Value |
|---|---|---|---|---|
| Aerococcaceae | Globicatella | 0.004 | −4.28 | 2.00 × 10−6 |
| Staphylococcaceae | Aliicoccus | 0.005 | −3.38 | 4.55 × 10−5 |
| Family XI_2 | Gallicola | 0.005 | −3.37 | 0.001126 |
| Family XI_2 | Anaerococcus | 0.010 | −3.15 | 3.23 × 10−5 |
| Carnobacteriaceae | Atopostipes | 0.005 | −3.12 | 0.000213 |
| Aerococcaceae | Facklamia | 0.122 | −2.45 | 3.23 × 10−5 |
| Dietziaceae | Dietzia | 0.038 | −2.37 | 9.76 × 10−7 |
| Streptococcaceae | Streptococcus | 1.535 | −1.44 | 0.000291 |
| Aerococcaceae | Aerococcus | 0.426 | −1.18 | 0.025612 |
| Ruminococcaceae | Ruminococcus 1 | 0.014 | −0.59 | 0.012061 |
| Lachnospiraceae | Marvinbryantia | 0.125 | −0.50 | 0.00272 |
| Lachnospiraceae | CHKCI001 | 0.166 | −0.39 | 0.013882 |
| Ruminococcaceae | Subdoligranulum | 1.084 | 0.24 | 0.011057 |
| Lachnospiraceae | Fusicatenibacter | 1.211 | 0.29 | 0.011057 |
| Ruminococcaceae | Ruminococcaceae UCG-013 | 0.040 | 0.33 | 0.004396 |
| Erysipelotrichaceae | Erysipelatoclostridium | 0.187 | 0.42 | 7.94 × 10−5 |
| Lachnospiraceae | Tyzzerella 3 | 0.004 | 0.47 | 0.020315 |
| X Defluviitaleaceae | Defluviitaleaceae UCG-011 | 0.018 | 0.48 | 0.020809 |
| Eggerthellaceae | Gordonibacter | 0.005 | 0.49 | 0.016537 |
| Anaeroplasmataceae | Anaeroplasma | 0.029 | 0.75 | 0.040975 |
| Rikenellaceae | Alistipes | 0.301 | 0.88 | 0.000422 |
| Enterococcaceae | Enterocuccus | 0.280 | 0.89 | 0.011057 |
| Lachnospiraceae | Ruminococcus gauveauii group | 0.001 | 1.07 | 0.017146 |
| Lachnospiraceae | Lachnoclostridium | 0.006 | 1.37 | 0.028353 |
| Bacillaceae | Oceanobacillus | 0.004 | 1.77 | 0.022935 |
| Veillonellaceae | Megamonas | 0.725 | 3.92 | 9.76 × 10−7 |
| Burkholderiaceae | Sutterella | 0.025 | 5.01 | 0.002039 |
| Helicobacteraceae | Helicobacter | 0.087 | 5.07 | 9.76 × 10−7 |
| Barnesiellaceae | Barnesiella | 0.037 | 5.20 | 0.000557 |
| Place of Sampling | Sample | Material Used 1 | No. Samples 2 | No. Pools |
|---|---|---|---|---|
| Anteroom | Boots | Sponge stick | All | 1 |
| Buckets | Sponge stick | All | 1 | |
| Door handle | Cotton swab | 2 | 2 | |
| Sink | Sponge stick | 1 | 1 | |
| Flies | Sticky paper | One strip | 1 | |
| Boot dip | Sponge stick | 1 | 1 | |
| Floor hygiene barrier | Sponge stick | 2 | 2 | |
| Poultry house | Beetles | Sterile tweezers | Undefined | 1 |
| Drinking nipples | Cotton swab | 20 | 4 | |
| Drinking bowl | Cotton swab | 20 | 4 | |
| Cracks in floor and walls | Cotton swab | 5 | 1 | |
| Puddles on floor | Sponge stick | 2 | 2 | |
| Air | Air sampler | 4 | 4 | |
| External environment | Puddles on concrete | Sponge stick | 2 | 2 |
| Cattle and/or swine | Overshoes | One pair | 1 | |
| Carcass container | Sponge stick | 1 | 1 | |
| Bird droppings | Sponge stick | 1 | 1 |
| Place of Sampling | Sample | Material Used 1 | No. Samples 2 | No. Pools |
|---|---|---|---|---|
| Anteroom | Boots | Sponge stick | All | 1 |
| Buckets | Sponge stick | All | 1 | |
| Door handle | Cotton swab | 2 | 2 | |
| Sink | Sponge stick | 1 | 1 | |
| Boot dip | Sponge stick | 1 | 1 | |
| Floor hygiene barrier | Sponge stick | 2 | 2 | |
| Wheelbarrow (wheels) | Sponge stick | 1 | 1 | |
| Poultry house | Broilers | Cecal droppings | 50 | 5 |
| Drinking nipples | Cotton swab | 20 | 4 | |
| Drinking bowl | Cotton swab | 20 | 4 | |
| Air | Airsampler | 4 | 4 | |
| External environment | Puddles on concrete | Sponge stick | 2 | 2 |
| Cattle and/or swine | Overshoes | One pair | 2 | |
| Carcass container | Sponge stick | 1 | 1 | |
| Bird droppings | Sponge stick | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hertogs, K.; Haegeman, A.; Schaumont, D.; Gelaude, P.; De Zutter, L.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter. Pathogens 2021, 10, 66. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010066
Hertogs K, Haegeman A, Schaumont D, Gelaude P, De Zutter L, Dewulf J, Heyndrickx M, Rasschaert G. Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter. Pathogens. 2021; 10(1):66. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010066
Chicago/Turabian StyleHertogs, Karolien, Annelies Haegeman, Dries Schaumont, Philippe Gelaude, Lieven De Zutter, Jeroen Dewulf, Marc Heyndrickx, and Geertrui Rasschaert. 2021. "Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter" Pathogens 10, no. 1: 66. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10010066